
Biotechnological Advances in Pharmacognosy and In Vitro Manipulation of Pterocarpus marsupium Roxb.



 ,
,  ,
,
Abstract
:1. Introduction
2. Botanical Description of P. marsupium
3. Phytochemistry and Therapeutic Values of P. marsupium
3.1. Active Constituents
3.2. Medicinal Properties
4. Propagation of Pterocarpus marsupium
4.1. Mechanism of Seed Germination
4.2. Propagation from Seed
4.3. Micropropagation through Various Methods
4.3.1. Cotyledonary Node Culture
4.3.2. Nodal Segment Culture
4.3.3. Shoot Tip Culture
4.3.4. Immature Zygotic Embryos Culture
4.3.5. Intact Seedling Culture
4.3.6. Somatic Embryogenesis
4.4. Rooting and Acclimatization
5. Molecular Studies of P. marsupium
5.1. Genetic Fidelity Assay
5.2. DNA Barcoding
5.3. Genetic Transformation
6. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keenan, R.J.; Reams, G.A.; Achard, F.; de Freitas, J.V.; Grainger, A.; Lindquist, E. Dynamics of global forest area: Results from the FAO Global Forest Resources Assessment 2015. For. Ecol. Manag. 2015, 352, 9–20. [Google Scholar] [CrossRef]
- Klitgaard, B.; Lavin, M.D. Dalbergieae: Legumes of the world. In Legumes of the World; Lewis, G.P., Schrire, B., MacKinder, B., Lock, M., Eds.; Royal Botanic Gardens Kew: Richmond, UK, 2005; pp. 307–335. [Google Scholar]
- Baker, E.G. The Leguminosae of Tropical Africa; Erasmus Press: Ghent, Belgium, 1929. [Google Scholar]
- Bentham, G. A Synopsis of Dalbergieae: A Tribe of the Leguminosae. J. Proc. Linn. Soc. 1860, 4, 65–80. [Google Scholar] [CrossRef]
- De Candolle, A.P. Memoires sur la Famille des Legumineuses; Auguste Belin: Paris, France, 1825. [Google Scholar]
- Lewis, G.P. Legumes of Bahia; Royal Botanic Gardens Kew: Richmond, UK, 1987. [Google Scholar]
- Rojo, J.P. Pterocarpus (Leguminosae-Papilionaceae) Revised for the World; Verlag Von J. Cramer: Lehre, Germany, 1972. [Google Scholar]
- Taubert, P. Leguminosae. In Die Natürlichen Pflanzenfamilien; Engler, A., Prantl, K., Eds.; Engelmann: Lemgo, Germany, 1894. [Google Scholar]
- Saslis-Lagoudakis, C.H.; Klitgaard, B.B.; Forest, F.; Francis, L.; Savolainen, V.; Williamson, E.M.; Hawkins, J.A. The use of phylogeny to interpret cross-cultural patterns in plant use and guide medicinal plant discovery: An example from Pterocarpus (Leguminosae). PLoS ONE 2011, 6, e22275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barstow, M. Pterocarpus marsupium. The IUCN Red List of Threatened Species 2017: E. T34620A67802995; IUCN: Colombo, Sri Lanka, 2017. [Google Scholar]
- Anis, M.; Ahmad, N. Plant Tissue Culture: Propagation, Conservation and Crop Improvement; Springer: Singapore, 2016. [Google Scholar]
- Phillips, G.C.; Garda, M. Plant tissue culture media and practices: An overview. In Vitr. Cell. Dev. Biol.-Plant 2019, 55, 242–257. [Google Scholar] [CrossRef]
- Dobránszki, J.; da Silva, J.A.T. Micropropagation of apple—A review. Biotechnol. Adv. 2010, 28, 462–488. [Google Scholar] [CrossRef]
- Teixeira da Silva, J.A.; Kher, M.M.; Soner, D.; Nataraj, M. Red sandalwood (Pterocarpus santalinus L. f.): Biology, importance, propagation and micropropagation. J. For. Res. 2019, 30, 745–754. [Google Scholar] [CrossRef] [Green Version]
- Teixeira da Silva, J.A.; Zeng, S.; Godoy-Hernández, G.; Rivera-Madrid, R.; Dobránszki, J. Bixa orellana L. (achiote) tissue culture: A review. In Vitr. Cell. Dev. Biol.-Plant 2019, 55, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Jiao, L.; Yu, M.; Wiedenhoeft, A.C.; He, T.; Li, J.; Liu, B.; Jiang, X.; Yin, Y. DNA Barcode Authentication and Library Development for the Wood of Six Commercial Pterocarpus Species: The Critical Role of Xylarium Specimens. Sci. Rep. 2018, 8, 1945. [Google Scholar] [CrossRef]
- Kress, W.J. Plant DNA barcodes: Applications today and in the future. J. Syst. Evol. 2017, 55, 291–307. [Google Scholar] [CrossRef] [Green Version]
- Kalimuthu, K.; Lakshmanan, K. Preliminary investigation on micropropagation of Pterocarpus marsupium Roxb. Indian J. For. 1994, 17, 192–195. [Google Scholar]
- Mishra, Y.; Rawat, R.; Nema, B.; Shirin, F. Effect of Seed Orientation and Medium Strength on In vitro Germination of Pterocarpus marsupium Roxb. Not. Sci. Biol. 2013, 5, 476–479. [Google Scholar] [CrossRef]
- Ahmad, A. In Vitro Morphogenesis and Assessment of Genetic Diversity in Pterocarpus marsupium Roxb. Using Molecular Markers; Aligarh Muslim University: Aligarh, India, 2019. [Google Scholar]
- Singh, P.; Bajpai, V.; Gupta, A.; Gaikwad, A.N.; Maurya, R.; Kumar, B. Identification and quantification of secondary metabolites of Pterocarpus marsupium by LC–MS techniques and its in-vitro lipid lowering activity. Ind. Crops Prod. 2019, 127, 26–35. [Google Scholar] [CrossRef]
- Ahmad, A.; Anis, M. Meta-topolin Improves In vitro Morphogenesis, Rhizogenesis and Biochemical Analysis in Pterocarpus marsupium Roxb.: A Potential Drug-Yielding Tree. J. Plant Growth Regul. 2019, 38, 1007–1016. [Google Scholar] [CrossRef]
- World Conservation Monitoring Centre (WCMC). Pterocarpus Marsupium. The IUCN Red List of Threatened Species; WCMC: Cambridge, UK, 1998. [Google Scholar]
- Chaudhuri, A.B.; Sarkar, D.D. Biodiversity Endangered: India’s Threatened Wildlife and Medicinal Plants; Scientific: Jodhpur, India, 2002. [Google Scholar]
- Anis, M.; Husain, M.K.; Shahzad, A. In vitro plantlet regeneration of Pterocarpus marsupium Roxb., an endangered leguminous tree. Curr. Sci. 2005, 88, 861–863. [Google Scholar]
- Tiwari, S.; Shah, P.; Singh, K. In vitro propagation of Pterocarpus marsupium Roxb.: An endangered medicinal tree. Indian J. Biotechnol. 2004, 3, 422–425. [Google Scholar]
- Garzuglia, M. Threatened, Endangered and Vulnerable Tree Species: A Comparison between FRA 2005 and the IUCN Red List; Forest Resources Assessment (FRA) Working Paper 108/E; FAO, Forestry Department: Rome, Italy, 2005. [Google Scholar]
- MOE. Ministry of Environment, The National Red List 2012 of Sri Lanka: Conservation Status of the Fauna and Flora; Ministry of Environment: Colombo, Sri Lanka, 2012.
- NMPB. National Medicinal Plant Board, Department of AYUSH, Ministry of Health and Family Welfare, Government of India, Agro Techniques of Selected Medicinal Plants; TERI Press: New Delhi, India, 2008. [Google Scholar]
- Maurya, R.; Singh, R.; Deepak, M.; Handa, S.S.; Yadav, P.P.; Mishra, P.K. Constituents of Pterocarpus marsupium: An ayurvedic crude drug. Phytochemistry 2004, 65, 915–920. [Google Scholar] [CrossRef] [PubMed]
- Jahromi, M.A.F.; Ray, A.B.; Chansouria, J.P.N. Antihyperlipidemic Effect of Flavonoids from Pterocarpus marsupium. J. Nat. Prod. 1993, 56, 989–994. [Google Scholar] [CrossRef]
- Manickam, M.; Ramanathan, M.; Farboodniay Jahromi, M.A.; Chansouria, J.P.N.; Ray, A.B. Antihyperglycemic Activity of Phenolics from Pterocarpus marsupium. J. Nat. Prod. 1997, 60, 609–610. [Google Scholar] [CrossRef]
- Maruthupandian, A.; Mohan, V. GC-MS analysis of some bioactive constituents of Pterocarpus marsupium Roxb. Int. J. Chem. Tech. Res. 2011, 3, 1652–1657. [Google Scholar]
- Maurya, R.; Ray, A.; Duah, F.; Slatkin, D.; Schiff, P., Jr. Constituents of Pterocarpus marsupium. J. Nat. Prod. 1984, 47, 179–181. [Google Scholar] [CrossRef]
- Chakravarthy, B.; Gode, K. Isolation of (-)-epicatechin from Pterocarpus marsupium and its pharmacological actions. Planta Med. 1985, 51, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, J.; Joshi, T. Flavonoids from Pterocarpus marsupium. Planta Med. 1988, 54, 371–372. [Google Scholar] [CrossRef]
- Tripathi, J.; Joshi, T. Phytochemical Investigation of Roots of Pterocarpus marsupium. Isolation and Structural Studies of Two New Flavanone Glycosides. Z. Nat. C 1988, 43, 184–186. [Google Scholar] [CrossRef] [PubMed]
- Handa, S.; Singh, R.; Maurya, R.; Satti, N.; Suri, K.; Suri, O. Pterocarposide, an isoaurone C-glucoside from Pterocarpus marsupium. Tetrahedron Lett. 2000, 41, 1579–1581. [Google Scholar] [CrossRef]
- Suri, K.; Satti, N.; Gupta, B.; Suri, O. 1-(2′, 6′-Dihydroxyphenyl)-β-glucopyranoside, a novel C-glycoside from Pterocarpus marsupium. Indian J. Chem. 2003, 42, 432–433. [Google Scholar] [CrossRef]
- Mutharaian, N.; Sasikumar, J.M.; Pavai, P.; Bai, V.N. In vitro antioxidant activity of Pterocarpus marsupium Roxb. Leaves. Int. J. Biomed. Pharm. Sci. 2009, 3, 29–33. [Google Scholar]
- Patil, U.H.; Gaikwad, D.K. Phytochemical screening and microbicidal activity of stem bark of Pterocarpus marsupium. Int. J. Pharm. Sci. Res. 2011, 2, 36–40. [Google Scholar]
- Mishra, A.; Srivastava, R.; Srivastava, S.P.; Gautam, S.; Tamrakar, A.K.; Maurya, R.; Srivastava, A.K. Antidiabetic activity of heart wood of Pterocarpus marsupium Roxb. and analysis of phytoconstituents. Indian J. Exp. Biol. 2013, 51, 363–374. [Google Scholar] [PubMed]
- Deguchi, T.; Miyamoto, A.; Miyamoto, K.; Kawata-Tominaga, T.; Yoshioka, Y.; Iwaki, M.; Murata, K. Determination of (+)-Dihydrorobinetin as An Active Constituent of the Radical-Scavenging Activity of Asana (Pterocarpus marsupium) Heartwood. Nat. Prod. Commun. 2019, 14, 1–5. [Google Scholar] [CrossRef]
- Yadav, V.K.; Mishra, A. In vitro & in silico study of hypoglycemic potential of Pterocarpus marsupium heartwood extract. Nat. Prod. Res. 2019, 33, 3298–3302. [Google Scholar] [CrossRef] [PubMed]
- Mohire, N.C.; Salunkhe, V.R.; Bhise, S.B.; Yadav, A.V. Cardiotonic activity of aqueous extract of heartwood of Pterocarpus marsupium. Indian J. Exp. Biol. 2007, 45, 532–537. [Google Scholar] [PubMed]
- Bressers, J. Botany of Ranchi District, Bihar, India; Catholic Press: Ranchi, India, 1951; p. 96. [Google Scholar]
- Trivedi, P.C. Medicinal Plants Traditional Knowledge; I.K. International Publishing House: New Delhi, India, 2006. [Google Scholar]
- Yesodharan, K.; Sujana, K. Ethnomedicinal knowledge among Malamalasar tribe of Parambikulam wildlife sanctuary, Kerala. Indian J. Tradit. Knowl. 2007, 6, 481–485. [Google Scholar]
- Chopra, R.N.; Nayar, R.L.; Chopra, I.C. Glossary of Indian Medicinal Plants; Council of Scientific and Industrial Research: New Dehli, India, 1956; p. 78. [Google Scholar]
- Anonymous. The Wealth of India; CSIR: New Delhi, India, 1969; Volume III. [Google Scholar]
- Pullaiah, T. Medicinal Plants of Andhra Pradesh (India); Regency Publication: New Delhi, India, 1999; p. 165. [Google Scholar]
- Hougee, S.; Faber, J.; Sanders, A.; de Jong, R.B.; van den Berg, W.B.; Garssen, J.; Hoijer, M.A.; Smit, H.F. Selective COX-2 Inhibition by a Pterocarpus marsupium Extract Characterized by Pterostilbene, and its Activity in Healthy Human Volunteers. Planta Med. 2005, 71, 387–392. [Google Scholar] [CrossRef] [Green Version]
- Tolomeo, M.; Grimaudo, S.; Cristina, A.D.; Roberti, M.; Pizzirani, D.; Meli, M.; Dusonchet, L.; Gebbia, N.; Abbadessa, V.; Crosta, L.; et al. Pterostilbene and 3′-hydroxypterostilbene are effective apoptosis-inducing agents in MDR and BCR-ABL-expressing leukemia cells. Int. J. Biochem. Cell Biol. 2005, 37, 1709–1726. [Google Scholar] [CrossRef]
- Remsberg, C.M.; Yáñez, J.A.; Ohgami, Y.; Vega-Villa, K.R.; Rimando, A.M.; Davies, N.M. Pharmacometrics of pterostilbene: Preclinical pharmacokinetics and metabolism, anticancer, antiinflammatory, antioxidant and analgesic activity. Phytother. Res. 2008, 22, 169–179. [Google Scholar] [CrossRef]
- Chakraborty, A.; Gupta, N.; Ghosh, K.; Roy, P. In vitro evaluation of the cytotoxic, anti-proliferative and anti-oxidant properties of pterostilbene isolated from Pterocarpus marsupium. Toxicol. In Vitr. 2010, 24, 1215–1228. [Google Scholar] [CrossRef]
- Tippani, R.; Vemunoori, A.K.; Yarra, R.; Nanna, R.S.; Abbagani, S.; Thammidala, C. Adventitious shoot regeneration from immature zygotic embryos of Indian Kino tree (Pterocarpus marsupium Roxb.) and genetic integrity analysis of in vitro derived plants using ISSR markers. Hortic. Environ. Biotechnol. 2013, 54, 531–537. [Google Scholar] [CrossRef]
- Maruthupandian, A.; Mohan, V. Antidiabetic, antihyperlipidaemic and antioxidant activity of Pterocarpus marsupium Roxb. in alloxan induced diabetic rats. Int. J. Pharm. Tech. Res. 2011, 3, 1681–1687. [Google Scholar]
- Tippani, R.; Jaya Shankar Prakhya, L.; Porika, M.; Sirisha, K.; Abbagani, S.; Thammidala, C. Pterostilbene as a potential novel telomerase inhibitor: Molecular docking studies and its in vitro evaluation. Curr. Pharm. Biotechnol. 2013, 14, 1027–1035. [Google Scholar] [CrossRef]
- Kosaraju, J.; Madhunapantula, S.V.; Chinni, S.; Khatwal, R.B.; Dubala, A.; Muthureddy Nataraj, S.K.; Basavan, D. Dipeptidyl peptidase-4 inhibition by Pterocarpus marsupium and Eugenia jambolana ameliorates streptozotocin induced Alzheimer’s disease. Behav. Brain Res. 2014, 267, 55–65. [Google Scholar] [CrossRef]
- Gupta, P.; Jain, V.; Pareek, A.; Kumari, P.; Singh, R.; Agarwal, P.; Sharma, V. Evaluation of effect of alcoholic extract of heartwood of Pterocarpus marsupium on in vitro antioxidant, anti-glycation, sorbitol accumulation and inhibition of aldose reductase activity. J. Tradit. Complementary Med. 2017, 7, 307–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, K.; Deguchi, T.; Yasuda, M.; Endo, R.; Fujita, T.; Matsumura, S.; Yoshioka, Y.; Matsuda, H. Improvement of Blood Rheology by Extract of Asana, Pterocarpus marsupium-Suppression of Platelet Aggregation Activity and Pterostilbene, as a Main Stilbene in the Extract. Nat. Prod. Commun. 2017, 12, 1089–1093. [Google Scholar] [CrossRef] [Green Version]
- Qadeer, F.; Abidi, A.; Fatima, F.; Rizvi, D.A. Effect of Pterocarpus marsupium in animal model of high carbohydrate diet-induced metabolic syndrome. Natl. J. Physiol. Pharm. Pharmacol. 2018, 8, 1509–1514. [Google Scholar] [CrossRef]
- Majeed, M.; Majeed, S.; Jain, R.; Mundkur, L.; Rajalakshmi, H.R.; Lad, P.; Neupane, P. A Randomized Study to Determine the Sun Protection Factor of Natural Pterostilbene from Pterocarpus marsupium. Cosmetics 2020, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- Satyavati, G.V.; Gupta, A.K.; Tandon, N. Medicinal Plants of India; ICMR Publications: New Delhi, India, 1987; Volume 2, p. 533.
- Grover, J.K.; Yadav, S.; Vats, V. Medicinal plants of India with anti-diabetic potential. J. Ethnopharmacol. 2002, 81, 81–100. [Google Scholar] [CrossRef]
- Chatterjee, A.; Pakrashi, S.C. The Treatise on Indian Medicinal Plants; Publications and Information Directorate: New Delhi, India, 1992; Volume 2. [Google Scholar]
- Grover, J.K.; Vats, V.; Yadav, S.S. Pterocarpus marsupium extract (Vijayasar) prevented the alteration in metabolic patterns induced in the normal rat by feeding an adequate diet containing fructose as sole carbohydrate. Diabetes Obes. Metab. 2005, 7, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Messina, F.; Guglielmini, G.; Curini, M.; Orsini, S.; Gresele, P.; Marcotullio, M.C. Effect of substituted stilbenes on platelet function. Fitoterapia 2015, 105, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Green, B.D.; Flatt, P.R.; Bailey, C.J. Dipeptidyl peptidase IV (DPP IV) inhibitors: A newly emerging drug class for the treatment of type 2 diabetes. Diabetes Vasc. Dis. Res. 2006, 3, 159–165. [Google Scholar] [CrossRef]
- Kosaraju, J.; Gali, C.C.; Khatwal, R.B.; Dubala, A.; Chinni, S.; Holsinger, R.M.D.; Madhunapantula, V.S.R.; Muthureddy Nataraj, S.K.; Basavan, D. Saxagliptin: A dipeptidyl peptidase-4 inhibitor ameliorates streptozotocin induced Alzheimer’s disease. Neuropharmacology 2013, 72, 291–300. [Google Scholar] [CrossRef]
- Beard, E.L., Jr. The American Society of Health System Pharmacists. JONA’s Healthc. Law Ethics Regul. 2001, 3, 78–79. [Google Scholar] [CrossRef] [PubMed]
- Smith, U. Abdominal obesity: A marker of ectopic fat accumulation. J. Clin. Investig. 2015, 125, 1790–1792. [Google Scholar] [CrossRef] [Green Version]
- Al-Goblan, A.S.; Al-Alfi, M.A.; Khan, M.Z. Mechanism linking diabetes mellitus and obesity. Diabetes Metab. Syndr. Obes. 2014, 7, 587–591. [Google Scholar] [CrossRef] [Green Version]
- Holtzman, M.J.; Turk, J.; Shornick, L.P. Identification of a pharmacologically distinct prostaglandin H synthase in cultured epithelial cells. J. Biol. Chem. 1992, 267, 21438–21445. [Google Scholar] [CrossRef]
- Hla, T.; Neilson, K. Human cyclooxygenase-2 cDNA. Proc. Natl. Acad. Sci. USA 1992, 89, 7384–7388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrasekharan, N.V.; Dai, H.; Roos, K.L.T.; Evanson, N.K.; Tomsik, J.; Elton, T.S.; Simmons, D.L. COX-3, a cyclooxygenase-1 variant inhibited by acetaminophen and other analgesic/antipyretic drugs: Cloning, structure, and expression. Proc. Natl. Acad. Sci. USA 2002, 99, 13926–13931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sander, S.; Smit, H.; Garssen, J.; Faber, J.; Hoijer, M. Pterocarpus marsupium extract exhibits antiinflammatory activity in human subjects. Planta Med. 2005, 71, 387–392. [Google Scholar]
- Kodan, A.; Kuroda, H.; Sakai, F. A stilbene synthase from Japanese red pine (Pinus densiflora): Implications for phytoalexin accumulation and down-regulation of flavonoid biosynthesis. Proc. Natl. Acad. Sci. USA 2002, 99, 3335–3339. [Google Scholar] [CrossRef] [Green Version]
- Frémont, L. Biological effects of resveratrol. Life Sci. 2000, 66, 663–673. [Google Scholar] [CrossRef]
- Stivala, L.A.; Savio, M.; Carafoli, F.; Perucca, P.; Bianchi, L.; Maga, G.; Forti, L.; Pagnoni, U.M.; Albini, A.; Prosperi, E.; et al. Specific Structural Determinants Are Responsible for the Antioxidant Activity and the Cell Cycle Effects of Resveratrol. J. Biol. Chem. 2001, 276, 22586–22594. [Google Scholar] [CrossRef] [Green Version]
- Rimando, A.M.; Cuendet, M.; Desmarchelier, C.; Mehta, R.G.; Pezzuto, J.M.; Duke, S.O. Cancer Chemopreventive and Antioxidant Activities of Pterostilbene, a Naturally Occurring Analogue of Resveratrol. J. Agric. Food Chem. 2002, 50, 3453–3457. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, P.; Asensi, M.; Segarra, R.; Ortega, A.; Benlloch, M.; Obrador, E.; Varea, M.T.; Asensio, G.; Jordá, L.; Estrela, J.M. Association between pterostilbene and quercetin inhibits metastatic activity of B16 melanoma. Neoplasia 2005, 7, 37–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husain, M.K.; Anis, M. Rapid in vitro multiplication of Melia azedarach L. (a multipurpose woody tree). Acta Physiol. Plant. 2009, 31, 765–772. [Google Scholar] [CrossRef]
- Javed, S.B.; Anis, M.; Khan, P.R.; Aref, I.M. In vitro regeneration and multiplication for mass propagation of Acacia ehrenbergiana Hayne: A potential reclaiment of denude arid lands. Agrofor. Syst. 2013, 87, 621–629. [Google Scholar] [CrossRef]
- Perveen, S.; Anis, M.; Aref, I.M. In vitro plant regeneration of Albizia lebbeck (L.) from seed explants. For. Syst. 2013, 22, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Vijayan, A.; Padmesh Pillai, P.; Hemanthakumar, A.S.; Krishnan, P.N. Improved In vitro propagation, genetic stability and analysis of corosolic acid synthesis in regenerants of Lagerstroemia speciosa (L.) Pers. by HPLC and gene expression profiles. Plant Cell Tissue Organ Cult. 2015, 120, 1209–1214. [Google Scholar] [CrossRef]
- Ahmed, M.R.; Anis, M.; Alatar, A.A.; Faisal, M. In vitro clonal propagation and evaluation of genetic fidelity using RAPD and ISSR marker in micropropagated plants of Cassia alata L.: A potential medicinal plant. Agrofor. Syst. 2017, 91, 637–647. [Google Scholar] [CrossRef]
- Javed, S.B.; Alatar, A.A.; Anis, M.; El-Sheikh, M.A. In vitro Regeneration of Coral Tree from Three Different Explants Using Thidiazuron. HortTechnology 2019, 29, 946–951. [Google Scholar] [CrossRef] [Green Version]
- Shu, K.; Liu, X.-d.; Xie, Q.; He, Z.-h. Two Faces of One Seed: Hormonal Regulation of Dormancy and Germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Miransari, M.; Smith, D.L. Plant hormones and seed germination. Environ. Exp. Bot. 2014, 99, 110–121. [Google Scholar] [CrossRef]
- Ludewig, K.; Zelle, B.; Eckstein, R.L.; Mosner, E.; Otte, A.; Donath, T.W. Differential effects of reduced water potential on the germination of floodplain grassland species indicative of wet and dry habitats. Seed Sci. Res. 2014, 24, 49–61. [Google Scholar] [CrossRef]
- White, C.N.; Rivin, C.J. Gibberellins and seed development in maize. II. Gibberellin synthesis inhibition enhances abscisic acid signaling in cultured embryos. Plant Physiol. 2000, 122, 1089–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuan, P.A.; Kumar, R.; Rehal, P.K.; Toora, P.K.; Ayele, B.T. Molecular Mechanisms Underlying Abscisic Acid/Gibberellin Balance in the Control of Seed Dormancy and Germination in Cereals. Front. Plant Sci. 2018, 9, 668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular Aspects of Seed Dormancy. Annu. Rev. Plant Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.P.; Lopez-Molina, L. A seed coat bedding assay to genetically explore in vitro how the endosperm controls seed germination in Arabidopsis thaliana. J. Vis. Exp. 2013, 2013, e50732. [Google Scholar] [CrossRef] [Green Version]
- Das, T.; Chatterjee, A. In vitro studies of Pterocarpus marsupium—An endangered tree. Indian J. Plant Physiol. 1993, 36, 269–272. [Google Scholar]
- Kalimuthu, K.; Lakshmanan, K. Effect of different treatments on pod germination of Pterocarpus species. Indian J. For. 1995, 18, 104–106. [Google Scholar]
- Husain, M.K.; Anis, M.; Shahzad, A. Seed germination studies in Pterocarpus marsupium Roxb., an important multipurpose tree. Hamdard Med. 2007, 50, 112–115. [Google Scholar]
- Lenser, T.; Theißen, G. Molecular mechanisms involved in convergent crop domestication. Trends Plant Sci. 2013, 18, 704–714. [Google Scholar] [CrossRef]
- Meyer, R.S.; Purugganan, M.D. Evolution of crop species: Genetics of domestication and diversification. Nat. Rev. Genet. 2013, 14, 840–852. [Google Scholar] [CrossRef]
- Simsek, S.; Ohm, J.-B.; Lu, H.; Rugg, M.; Berzonsky, W.; Alamri, M.S.; Mergoum, M. Effect of pre-harvest sprouting on physicochemical changes of proteins in wheat. J. Sci. Food Agric. 2014, 94, 205–212. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Rolston, M.P. Water impermeable seed dormancy. Bot. Rev. 1978, 44, 365–396. [Google Scholar] [CrossRef]
- Khan, A.A. Hormonal regulation of primary and secondary seed dormancy. Isr. J. Bot. 1980, 29, 207–224. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Tigabu, M.; Oden, P. Effect of scarification, gibberellic acid and temperature on seed germination of two multipurpose Albizia species from Ethiopia. Seed Sci. Technol. 2001, 29, 11–20. [Google Scholar]
- Barmukh, R.; Nikam, T. Promotion of seed germination in Pterocarpus marsupium Roxb. Indian J. Plant Physiol. 2008, 13, 143–150. [Google Scholar]
- Baskin, J.M.; Baskin, C.C.; Spooner, D.M. Role of temperature, light and date: Seeds were exhumed from soil on germination of four wetland perennials. Aquat. Bot. 1989, 35, 387–394. [Google Scholar] [CrossRef]
- Azad, S.; Manik, M.R.; Hasan, S.; Matin, A. Effect of different pre-sowing treatments on seed germination percentage and growth performance of Acacia auriculiformis. J. For. Res. 2011, 22, 183. [Google Scholar] [CrossRef]
- Tadros, M.J.; Samarah, N.H.; Alqudah, A.M. Effect of different pre-sowing seed treatments on the germination of Leucaena leucocephala (Lam.) and Acacia farnesiana (L.). New For. 2011, 42, 397–407. [Google Scholar] [CrossRef]
- Dayanand, T.; Lohidas, T. Effect of different treatments on pod germination of red sanders (Pterocarpus santalinus Linn.). Indian J. For. 1988, 11, 87–88. [Google Scholar]
- Park, J.; Kim, Y.-S.; Kim, S.-G.; Jung, J.-H.; Woo, J.-C.; Park, C.-M. Integration of auxin and salt signals by the NAC transcription factor NTM2 during seed germination in Arabidopsis. Plant Physiol. 2011, 156, 537–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- White, P.R. The Cultivation of Animal and Plant Cells, 2nd ed.; Ronald Press: New York, NY, USA, 1963. [Google Scholar]
- Ahmad, N.; Alia, A.; Khan, F.; Kour, A.; Khan, S. In vitro seed germination and shoot multiplication of Pterocarpus marsupium Roxb.—An endangered medicinal tree. Researcher 2012, 4, 20–24. [Google Scholar]
- Lee, J.-H.; Kim, Y.-K.; Oh, E.-Y.; Jung, K.-Y.; Ko, K.-S. Optimization of In vitro seed germination of Taraxacum platycarpum. Korean J. Environ. Agric. 2009, 28, 403–408. [Google Scholar] [CrossRef] [Green Version]
- Masilamani, P.; Singh, B.G.; Chinnusamy, C.; Annadurai, K. Short communication: Influence af seed orientation and depth of sowing on germination and vigour of Anjan (Hardwickia binata Roxb). Trop. Agric. Res. Ext. 1999, 2, 76–78. [Google Scholar]
- Ahmad, A.; Siddique, I.; Varshney, A. Selection of Elite Genotype of a Multipurpose Forest Tree (Pterocarpus marsupium Roxb.) from Naturally Grown Populations in Central India. In Propagation and Genetic Manipulation of Plants; Siddique, I., Ed.; Springer: Singapore, 2021; pp. 101–119. [Google Scholar]
- Lee, S.; Cheng, H.; King, K.E.; Wang, W.; He, Y.; Hussain, A.; Lo, J.; Harberd, N.P.; Peng, J. Gibberellin regulates Arabidopsis seed germination via RGL2, a GAI/RGA-like gene whose expression is up-regulated following imbibition. Genes Dev. 2002, 16, 646–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varshney, A.; Anis, M.; Aref, I. Control of bioregulants on plant resurgence in vitro from mature seeds of Egyptian myrobalan tree (Balanites aegyptiaca del.)—A plant affluent in saponins. Int. J. Pharm. Sci. Rev. Res. 2013, 22, 23–28. [Google Scholar]
- Justamante, M.S.; Ibáñez, S.; Villanova, J.; Pérez-Pérez, J.M. Vegetative propagation of argan tree (Argania spinosa (L.) Skeels) using in vitro germinated seeds and stem cuttings. Sci. Hortic. 2017, 225, 81–87. [Google Scholar] [CrossRef]
- Khazaei Kojori, Z.; Rezaei, M.; Sarkhosh, A.; Gharangik, S. The effects of bud-scale removing and gibberellin (GA3) on dormancy break of apricot (P. armeniaca L.) vegetative buds. J. Appl. Hortic. 2018, 20, 50–54. [Google Scholar] [CrossRef]
- Gubler, F.; Chandler, P.M.; White, R.G.; Llewellyn, D.J.; Jacobsen, J.V. Gibberellin signaling in barley aleurone cells. Control of SLN1 and GAMYB expression. Plant Physiol. 2002, 129, 191–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.-F.; He, J.-X. Mechanisms of signaling crosstalk between brassinosteroids and gibberellins. Plant. Signal. Behav. 2013, 8, e24686. [Google Scholar] [CrossRef] [Green Version]
- Weiss, D.; Ori, N. Mechanisms of cross talk between gibberellin and other hormones. Plant Physiol. 2007, 144, 1240–1246. [Google Scholar] [CrossRef] [Green Version]
- Gantait, S.; Kundu, S.; Das, P.K. Acacia: An exclusive survey on in vitro propagation. J. Saudi Soc. Agric. Sci. 2018, 17, 163–177. [Google Scholar] [CrossRef] [Green Version]
- Fatima, N.; Anis, M. Role of growth regulators on In vitro regeneration and histological analysis in Indian ginseng (Withania somnifera L.) Dunal. Physiol. Mol. Biol. Plants 2012, 18, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, N.; Javed, S.B.; Khan, M.I.; Anis, M. Rapid plant regeneration and analysis of genetic fidelity in micropropagated plants of Vitex trifolia: An important medicinal plant. Acta Physiol. Plant. 2013, 35, 2493–2500. [Google Scholar] [CrossRef]
- Nagar, D.S.; Jha, S.K.; Jani, J. Direct adventitious shoot bud formation on hypocotyls explants in Millettia pinnata (L.) Panigrahi-a biodiesel producing medicinal tree species. Physiol. Mol. Biol. Plants 2015, 21, 287–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perveen, S.; Khanam, M.N.; Anis, M.; Atta, H.A.E. In vitro mass propagation of Murraya koenigii L. J. Appl. Res. Med. Aromat. Plants 2015, 2, 60–68. [Google Scholar] [CrossRef]
- Howell, S.H.; Lall, S.; Che, P. Cytokinins and shoot development. Trends Plant Sci. 2003, 8, 453–459. [Google Scholar] [CrossRef]
- Santner, A.; Estelle, M. Recent advances and emerging trends in plant hormone signalling. Nature 2009, 459, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Moubayidin, L.; Di Mambro, R.; Sabatini, S. Cytokinin–auxin crosstalk. Trends Plant Sci. 2009, 14, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.W.; Werr, W. Cytokinin–auxin crosstalk in cell type specification. Trends Plant Sci. 2015, 20, 291–300. [Google Scholar] [CrossRef]
- Chand, S.; Singh, A.K. In vitro shoot regeneration from cotyledonary node explants of a multipurpose leguminous tree Pterocarpus marsupium Roxb. In Vitr. Cell. Dev. Biol.-Plant 2004, 40, 464–466. [Google Scholar] [CrossRef]
- Husain, M.K.; Anis, M.; Shahzad, A. In vitro propagation of Indian Kino (Pterocarpus marsupium Roxb.) using Thidiazuron. In Vitr. Cell. Dev. Biol.-Plant 2007, 43, 59–64. [Google Scholar] [CrossRef]
- Pradhan, C.; Kar, S.; Pattnaik, S.; Chand, P.K. Propagation of Dalbergia sissoo Roxb. through In vitro shoot proliferation from cotyledonary nodes. Plant Cell Rep. 1998, 18, 122–126. [Google Scholar] [CrossRef]
- Da Silva, A.L.L.; Gollo, A.L.; Brondani, G.E.; Horbach, M.A.; Oliveira, L.; Machado, M.P.; Lima, K.; Costa, J. Micropropagation of Eucalyptus saligna Sm. from cotyledonary nodes. Pak. J. Bot. 2015, 47, 311–318. [Google Scholar]
- Moharana, A.; Das, A.; Subudhi, E.; Naik, S.K.; Barik, D.P. High frequency shoot proliferation from cotyledonary node of Lawsonia inermis L. and validation of their molecular finger printing. J. Crop Sci. Biotechnol. 2018, 20, 405–416. [Google Scholar] [CrossRef]
- Venkatachalam, P.; Jinu, U.; Gomathi, M.; Mahendran, D.; Ahmad, N.; Geetha, N.; Sahi, S.V. Role of silver nitrate in plant regeneration from cotyledonary nodal segment explants of Prosopis cineraria (L.) Druce.: A recalcitrant medicinal leguminous tree. Biocatal. Agric. Biotechnol. 2017, 12, 286–291. [Google Scholar] [CrossRef]
- Gupta, A.; Thomas, T.; Khan, S. Regeneration of Aegle marmelos (l.) Through Enhanced Axillary Branching from Cotyledenory Node. UK J. Pharm. Biosci. 2018, 6, 24. [Google Scholar] [CrossRef]
- Husain, M.K.; Anis, M.; Shahzad, A. In vitro propagation of a multipurpose leguminous tree (Pterocarpus marsupium Roxb.) using nodal explants. Acta Physiol. Plant. 2008, 30, 353–359. [Google Scholar] [CrossRef]
- Husain, M.K.; Anis, M.; Shahzad, A. Somatic embryogenesis and plant regeneration in Pterocarpus marsupium Roxb. Trees 2010, 24, 781–787. [Google Scholar] [CrossRef]
- Tippani, R.; Yarra, R.; Bulle, M.; Porika, M.; Abbagani, S.; Thammidala, C. In vitro plantlet regeneration and Agrobacterium tumefaciens-mediated genetic transformation of Indian Kino tree (Pterocarpus marsupium Roxb.). Acta Physiol. Plant. 2013, 35, 3437–3446. [Google Scholar] [CrossRef]
- Jaiswal, S.; Choudhary, M.; Arya, S.; Kant, T. Micropropagation of adult tree of Pterocarpus marsupium Roxb. using nodal explants. J. Plant Dev. 2015, 22, 21–30. [Google Scholar]
- Ahmad, A.; Ahmad, N.; Anis, M. Preconditioning of Nodal Explants in Thidiazuron-Supplemented Liquid Media Improves Shoot Multiplication in Pterocarpus marsupium (Roxb.). In Thidiazuron: From Urea Derivative to Plant Growth Regulator; Ahmad, N., Faisal, M., Eds.; Springer: Singapore, 2018; pp. 175–187. [Google Scholar]
- Tippani, R.; Nanna, R.S.; Mamidala, P.; Thammidala, C. Assessment of genetic stability in somatic embryo derived plantlets of Pterocarpus marsupium Roxb. using inter-simple sequence repeat analysis. Physiol. Mol. Biol. Plants 2019, 25, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Anis, M.; Khanam, M.N.; Alatar, A.A. Direct shoot organogenesis from shoot tip explants of a highly medicinal valued tree Pterocarpus marsupium Roxb. In Vitr. Cell. Dev. Biol.-Plant 2020, 56, 670–681. [Google Scholar] [CrossRef]
- Ahmad, A.; Ahmad, N.; Anis, M.; Alatar, A.A.; Abdel-Salam, E.M.; Qahtan, A.A.; Faisal, M. Gibberellic acid and thidiazuron promote micropropagation of an endangered woody tree (Pterocarpus marsupium Roxb.) using in vitro seedlings. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 144, 449–462. [Google Scholar] [CrossRef]
- Kubalakova, M.; Strnad, M. The effects of aromatic cytokinins (populins) on micropropagation and regeneration of sugar beet in vitro. Biol. Plant 1992, 34, 578–579. [Google Scholar]
- Strnad, M.; Hanuš, J.; Vaněk, T.; Kamínek, M.; Ballantine, J.A.; Fussell, B.; Hanke, D.E. Meta-topolin, a highly active aromatic cytokinin from poplar leaves (Populus × canadensis Moench., cv. Robusta). Phytochemistry 1997, 45, 213–218. [Google Scholar] [CrossRef]
- Amoo, S.O.; Finnie, J.F.; Van Staden, J. The role of meta-topolins in alleviating micropropagation problems. Plant Growth Regul. 2011, 63, 197–206. [Google Scholar] [CrossRef]
- Aremu, A.O.; Bairu, M.W.; Szüčová, L.; Dolezal, K.; Finnie, J.; Van Staden, J. Assessment of the role of meta-topolins on in vitro produced phenolics and acclimatization competence of micropropagated ‘Williams’ banana. Acta Physiol. Plant. 2012, 34, 2265–2273. [Google Scholar] [CrossRef]
- Werbrouck, S.P.O.; Strnad, M.; Van Onckelen, H.A.; Debergh, P.C. Meta-topolin, an alternative to benzyladenine in tissue culture? Physiol. Plant. 1996, 98, 291–297. [Google Scholar] [CrossRef]
- Gentile, A.; Jàquez Gutiérrez, M.; Martinez, J.; Frattarelli, A.; Nota, P.; Caboni, E. Effect of meta-Topolin on micropropagation and adventitious shoot regeneration in Prunus rootstocks. Plant Cell Tissue Organ Cult. 2014, 118, 373–381. [Google Scholar] [CrossRef]
- Drewes, F.E.; Van Staden, J. The effect of 6-benzyladenine derivatives on the rooting of Phaseolus vulgaris L. primary leaf cuttings. Plant Growth Regul. 1989, 8, 289–296. [Google Scholar] [CrossRef]
- Mala, J.; Machova, P.; Cvrckova, H.; Karady, M.; Novak, O.; Mikulik, J.; Dostal, J.; Strnad, M.; Dolezal, K. The role of cytokinins during micropropagation of wych elm. Biol. Plant. 2013, 57, 174–178. [Google Scholar] [CrossRef]
- Liu, J.; Moore, S.; Chen, C.; Lindsey, K. Crosstalk Complexities between Auxin, Cytokinin, and Ethylene in Arabidopsis Root Development: From Experiments to Systems Modeling, and Back Again. Mol. Plant 2017, 10, 1480–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magyar-Tábori, K.; Dobránszki, J.; Teixeira da Silva, J.A.; Bulley, S.M.; Hudák, I. The role of cytokinins in shoot organogenesis in apple. Plant Cell Tissue Organ Cult. 2010, 101, 251–267. [Google Scholar] [CrossRef]
- Naaz, A.; Hussain, S.A.; Anis, M.; Alatar, A.A. Meta-topolin improved micropropagation in Syzygium cumini and acclimatization to ex vitro conditions. Biol. Plant. 2019, 63, 174–182. [Google Scholar] [CrossRef]
- Webster, C.A.; Jones, O.P. Micropropagation of some cold-hardy dwarfing rootstocks for apple. J. Hortic. Sci. 1991, 66, 1–6. [Google Scholar] [CrossRef]
- Huyluoğlu, Z.; Ünal, M.; Palavan-Ünsal, N. Cytological evidences of the role of meta-topolin and Benzyladenin in barley root tips. Adv. Mol. Biol. 2008, 2, 31–37. [Google Scholar]
- Bairu, M.W.; Kulkarni, M.G.; Street, R.A.; Mulaudzi, R.B.; Van Staden, J. Studies on seed germination, seedling growth, and In vitro shoot induction of Aloe ferox Mill., a commercially important species. HortScience 2009, 44, 751–756. [Google Scholar] [CrossRef] [Green Version]
- De Diego, N.; Montalbán, I.; Moncaleán, P. In vitro regeneration of Pinus spp. adult trees: New method for obtaining clonal plants. Acta Hortic. 2008, 865, 361–365. [Google Scholar] [CrossRef]
- Nas, M.N.; Bolek, Y.; Sevgin, N. The effects of explant and cytokinin type on regeneration of Prunus microcarpa. Sci. Hortic. 2010, 126, 88–94. [Google Scholar] [CrossRef]
- Meyer, E.M.; Touchell, D.H.; Ranney, T.G. In vitro Shoot Regeneration and Polyploid Induction from Leaves of Hypericum Species. HortScience 2009, 44, 1957–1961. [Google Scholar] [CrossRef] [Green Version]
- Niedz, R.P.; Evens, T.J. The effects of benzyladenine and meta-topolin on In vitro shoot regeneration of a Citrus citrandarin rootstock. Res. J. Agric. Biol. Sci. 2010, 6, 45–53. [Google Scholar]
- Vasudevan, R.; Van Staden, J. Cytokinin and explant types influence in vitro plant regeneration of Leopard Orchid (Ansellia africana Lindl.). Plant Cell Tissue Organ Cult. 2011, 107, 123–129. [Google Scholar] [CrossRef]
- Magyar-Tábori, K.; Dobránszky, J.; Jámbor-Benczúr, É.; Bubán, T.; Lazányi, J.; Szalai, J.; Ferenczy, A. Post-effects of cytokinins and auxin levels of proliferation media on rooting ability of in vitro apple shoots (Malus domestica Borkh.) ’Red Fuji’. Int. J. Hortic. Sci. 2001, 7, 26–29. [Google Scholar] [CrossRef] [Green Version]
- Bairu, M.W.; Stirk, W.A.; Dolezal, K.; Van Staden, J. Optimizing the micropropagation protocol for the endangered Aloe polyphylla: Can meta-topolin and its derivatives serve as replacement for benzyladenine and zeatin? Plant Cell Tissue Organ Cult. 2007, 90, 15–23. [Google Scholar] [CrossRef]
- Bairu, M.W.; Stirk, W.A.; Doležal, K.; van Staden, J. The role of topolins in micropropagation and somaclonal variation of banana cultivars ‘Williams’ and ‘Grand Naine’ (Musa spp. AAA). Plant Cell Tissue Organ Cult. 2008, 95, 373–379. [Google Scholar] [CrossRef]
- Valero-Aracama, C.; Kane, M.E.; Wilson, S.B.; Philman, N.L. Substitution of benzyladenine with meta-topolin during shoot multiplication increases acclimatization of difficult- and easy-to-acclimatize sea oats (Uniola paniculata L.) genotypes. Plant Growth Regul. 2010, 60, 43. [Google Scholar] [CrossRef]
- Dobránszky, J.; Jámbor-Benczúr, E.; Reményi, M.L.; Magyar-Tábori, K.; Hudák, I.; Kiss, E.; Galli, Z. Effects of aromatic cytokinins on structural characteristics of leaves and their post-effects on subsequent shoot regeneration from in vitro apple leaves of ’Royal Gala’. Int. J. Hortic. Sci. 2005, 11, 41–46. [Google Scholar] [CrossRef]
- Čatský, J.; Pospíšilová, J.; Kamínek, M.; Gaudinová, A.; Pulkrábek, J.; Zahradníček, J. Seasonal changes in sugar beet photosynthesis as affected by exogenous cytokininN6-(m-hydroxybenzyl)adenosine. Biol. Plant. 1996, 38, 511–518. [Google Scholar] [CrossRef]
- Palavan-Ünsal, N.; Çağ, S.; Çetin, E.; Büyüktunçer, D. Retardation of senescence by meta-topolin in wheat leaves. J. Cell Mol. Biol. 2002, 1, 101–108. [Google Scholar]
- Wojtania, A. Effect of Meta-topolin on in vitro propagation of Pelargonium × hortorum and Pelargonium × hederaefolium cultivars. Acta Soc. Bot. Pol. 2011, 79, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Bogaert, I.; Van Cauter, S.; Werbrouck, S.; Dolezal, K. New aromatic cytokinins can make the difference. Acta Hortic. 2004, 725, 265–270. [Google Scholar] [CrossRef]
- Bairu, M.; Jain, N.; Stirk, W.; Doležal, K.; Van Staden, J. Solving the problem of shoot-tip necrosis in Harpagophytum procumbens by changing the cytokinin types, calcium and boron concentrations in the medium. S. Afr. J. Bot. 2009, 75, 122–127. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Whiting, M.D. Improving ‘Bing’ sweet cherry fruit quality with plant growth regulators. Sci. Hortic. 2011, 127, 341–346. [Google Scholar] [CrossRef]
- Murch, S.J.; Saxena, P.K. Molecular fate of thidiazuron and its effects on auxin transport in hypocotyls tissues of Pelargonium × hortorum Bailey. Plant Growth Regul. 2001, 35, 269–275. [Google Scholar] [CrossRef]
- Guo, B.; Abbasi, B.H.; Zeb, A.; Xu, L.L.; Wei, Y.H. Thidiazuron: A multi-dimensional plant growth regulator. Afr. J. Biotechnol. 2011, 10, 8984–9000. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, N.; Anis, M. Rapid clonal multiplication of a woody tree, Vitex negundo L. through axillary shoots proliferation. Agrofor. Syst. 2007, 71, 195–200. [Google Scholar] [CrossRef]
- Dhavala, A.; Rathore, T.S. Micropropagation of Embelia ribes Burm f. through proliferation of adult plant axillary shoots. In Vitr. Cell. Dev. Biol.-Plant 2010, 46, 180–191. [Google Scholar] [CrossRef]
- Singh, M.; Jaiswal, V.; Jaiswal, U. Thidiazuron-induced anatomical changes and direct shoot morphogenesis in Dendrocalamus strictus Nees. Can. J. Pure Appl. Sci. 2014, 8, 2901–2904. [Google Scholar]
- Murthy, B.N.S.; Murch, S.J.; Saxena, P.K. Thidiazuron-induced somatic embryogenesis in intact seedlings of peanut (Arachis hypogaea): Endogenous growth regulator levels and significance of cotyledons. Physiol. Plant. 1995, 94, 268–276. [Google Scholar] [CrossRef]
- Zhang, C.G.; Li, W.; Mao, Y.F.; Zhao, D.L.; Dong, W.; Guo, G.Q. Endogenous Hormonal Levels in Scutellaria baicalensis Calli Induced by Thidiazuron. Russ. J. Plant Physiol. 2005, 52, 345–351. [Google Scholar] [CrossRef]
- Preece, J.E.; Imel, M.R. Plant regeneration from leaf explants of Rhododendron ‘P.J.M. Hybrids’. Sci. Hortic. 1991, 48, 159–170. [Google Scholar] [CrossRef]
- Sudha, C.; Seeni, S. In vitro multiplication and field establishment of Adhatoda beddomei C. B. Clarke, a rare medicinal plant. Plant Cell Rep. 1994, 13, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Faisal, M.; Ahmad, N.; Anis, M. Shoot multiplication in Rauvolfia tetraphylla L. using thidiazuron. Plant Cell Tissue Organ Cult. 2005, 80, 187–190. [Google Scholar] [CrossRef]
- Hussain, S.A.; Ahmad, N.; Anis, M. Synergetic effect of TDZ and BA on minimizing the post-exposure effects on axillary shoot proliferation and assessment of genetic fidelity in Rauvolfia tetraphylla (L.). Rend. Lincei Sci. Fis. Nat. 2018, 29, 109–115. [Google Scholar] [CrossRef]
- Lu, C.-Y. The use of thidiazuron in tissue culture. In Vitr. Cell. Dev. Biol.-Plant 1993, 29, 92–96. [Google Scholar] [CrossRef]
- Singh, S.K.; Syamal, M.M. A short pre-culture soak in thidiazuron or forchlorfenuron improves axillary shoot proliferation in rose micropropagation. Sci. Hortic. 2001, 91, 169–177. [Google Scholar] [CrossRef]
- Ahmad, N.; Siddique, I.; Anis, M. Improved plant regeneration in Capsicum annuum L. from nodal segments. Biol. Plant. 2006, 50, 701–704. [Google Scholar] [CrossRef]
- Faisal, M.; Anis, M. Thidiazuron induced high frequency axillary shoot multiplication in Psoralea corylifolia. Biol. Plant. 2006, 50, 437–440. [Google Scholar] [CrossRef]
- Huetteman, C.A.; Preece, J.E. Thidiazuron: A potent cytokinin for woody plant tissue culture. Plant Cell Tissue Organ Cult. 1993, 33, 105–119. [Google Scholar] [CrossRef]
- Fasolo, F.; Zimmerman, R.H.; Fordham, I. Adventitions shoot formation on excised leaves of in vitro grown shoots of apple cultivars. Plant Cell Tissue Organ Cult. 1989, 16, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Vengadesan, G.; Ganapathi, A.; Prem Anand, R.; Ramesh Anbazhagan, V. In vitro propagation of Acacia sinuata (Lour.) Merr. via cotyledonary nodes. Agrofor. Syst. 2002, 55, 9–15. [Google Scholar] [CrossRef]
- Thomas, T. Thidiazuron Induced Multiple Shoot Induction and Plant Regeneration from Cotyledonary Explants of Mulberry. Biol. Plant. 2003, 46, 529–533. [Google Scholar] [CrossRef]
- Patri, S.; Bhatnagar, S.; Bhojwani, S.S. Preliminary investigations on micropropagation of a leguminous timber tree: Pterocarpus santalinus. Phytomorphology 1988, 38, 41–45. [Google Scholar]
- Anuradha, M.; Pullaiah, T. In vitro seed culture and induction of enhanced axillary branching in Pterocarpus santalinus and Pterocarpus marsupium: A method for rapid multiplication. Phytomorphology 1999, 49, 157–163. [Google Scholar]
- Loc, N.H.; Duc, D.T.; Kwon, T.H.; Yang, M.S. Micropropagation of zedoary (Curcuma zedoaria Roscoe): A valuable medicinal plant. Plant Cell Tissue Organ Cult. 2005, 81, 119–122. [Google Scholar] [CrossRef]
- Nayak, P.; Behera, P.R.; Manikkannan, T. High frequency plantlet regeneration from cotyledonary node cultures of Aegle marmelos (L.) Corr. In Vitr. Cell. Dev. Biol.-Plant 2007, 43, 231–236. [Google Scholar] [CrossRef]
- Purkayastha, J.; Sugla, T.; Paul, A.; Solleti, S.; Sahoo, L. Rapid In vitro multiplication and plant regeneration from nodal explants of Andrographis paniculata: A valuable medicinal plant. In Vitr. Cell. Dev. Biol.-Plant 2008, 44, 442–447. [Google Scholar] [CrossRef]
- Perveen, S.; Varshney, A.; Anis, M.; Aref, I.M. Influence of cytokinins, basal media and pH on adventitious shoot regeneration from excised root cultures of Albizia lebbeck. J. For. Res. 2011, 22, 47–52. [Google Scholar] [CrossRef]
- Phulwaria, M.; Rai, M.K.; Harish; Gupta, A.K.; Ram, K.; Shekhawat, N.S. An improved micropropagation of Terminalia bellirica from nodal explants of mature tree. Acta Physiol. Plant. 2011, 34, 299–305. [Google Scholar] [CrossRef]
- Varshney, A.; Anis, M.; Rasheed, M.; Khan, P.; Aref, I. Assessment of changes in physiological and biochemical behaviors in Grey-haired Acacia tree (Acacia gerrardii)—An important plant of arid region. Int. J. Sci. Res. Eng. Res. 2013, 4, 1134–1156. [Google Scholar]
- Gururaj, H.B.; Giridhar, P.; Ravishankar, G.A. Micropropagation of Tinospora cordifolia (Willd.) Miers ex Hook. F & Thoms—A multipurpose medicinal plant. Curr. Sci. 2007, 92, 23–26. [Google Scholar]
- Chaudhary, H.; Sood, N. Purification and partial characterization of lectins from In vitro cultures of Ricinus communis. Plant Tissue Cult. Biotechnol. 2008, 18, 89–102. [Google Scholar] [CrossRef]
- Harisharanraj, R.; Suresh, K.; Saravanababu, S. Rapid clonal propagation of Rauvolfia tetraphylla L. Acad. J. Plant Sci. 2009, 2, 195–198. [Google Scholar]
- Ozudogru, E.A.; Kaya, E.; Kirdok, E.; Issever-Ozturk, S. In vitro propagation from young and mature explants of thyme (Thymus vulgaris and T. longicaulis) resulting in genetically stable shoots. In Vitr. Cell. Dev. Biol.-Plant 2011, 47, 309–320. [Google Scholar] [CrossRef]
- Siddique, I.; Anis, M.; Aref, I.M. In vitro Adventitious Shoot Regeneration via Indirect Organogenesis from Petiole Explants of Cassia angustifolia Vahl.—A Potential Medicinal Plant. Appl. Biochem. Biotechnol. 2010, 162, 2067–2074. [Google Scholar] [CrossRef]
- Parveen, S.; Shahzad, A. Factors affecting In vitro plant regeneration from cotyledonary node explant of Senna sophera (L.) Roxb. a highly medicinal legume. Afr. J. Biotechnol. 2014, 13, 413–422. [Google Scholar] [CrossRef]
- Naaz, A.; Hussain, S.A.; Naz, R.; Anis, M.; Alatar, A.A. Successful plant regeneration system via de novo organogenesis in Syzygium cumini (L.) Skeels: An important medicinal tree. Agrofor. Syst. 2019, 93, 1285–1295. [Google Scholar] [CrossRef]
- Ferguson, B.J.; Beveridge, C.A. Roles for Auxin, Cytokinin, and Strigolactone in Regulating Shoot Branching. Plant Physiol. 2009, 149, 1929–1944. [Google Scholar] [CrossRef] [Green Version]
- Müller, D.; Leyser, O. Auxin, cytokinin and the control of shoot branching. Ann. Bot. 2011, 107, 1203–1212. [Google Scholar] [CrossRef] [Green Version]
- Anis, M.; Varshney, A.; Siddique, I. In vitro clonal propagation of Balanites aegyptiaca (L.) Del. Agrofor. Syst. 2010, 78, 151–158. [Google Scholar] [CrossRef]
- Faisal, M.; Alatar, A.A.; Ahmad, N.; Anis, M.; Hegazy, A.K. An Efficient and Reproducible Method for in vitro Clonal Multiplication of Rauvolfia tetraphylla L. and Evaluation of Genetic Stability using DNA-Based Markers. Appl. Biochem. Biotechnol. 2012, 168, 1739–1752. [Google Scholar] [CrossRef] [PubMed]
- Siddique, I.; Javed, S.B.; Al-Othman, M.R.; Anis, M. Stimulation of In vitro organogenesis from epicotyl explants and successive micropropagation round in Cassia angustifolia Vahl.: An important source of sennosides. Agrofor. Syst. 2013, 87, 583–590. [Google Scholar] [CrossRef]
- Danova, K.; Motyka, V.; Todorova, M.; Trendafilova, A.; Krumova, S.; Dobrev, P.; Andreeva, T.; Oreshkova, T.; Taneva, S.; Evstatieva, L. Effect of Cytokinin and Auxin Treatments on Morphogenesis, Terpenoid Biosynthesis, Photosystem Structural Organization, and Endogenous Isoprenoid Cytokinin Profile in Artemisia alba Turra In vitro. J. Plant Growth Regul. 2018, 37, 403–418. [Google Scholar] [CrossRef]
- Nordström, A.; Tarkowski, P.; Tarkowska, D.; Norbaek, R.; Astot, C.; Dolezal, K.; Sandberg, G. Auxin regulation of cytokinin biosynthesis in Arabidopsis thaliana: A factor of potential importance for auxin-cytokinin-regulated development. Proc. Natl. Acad. Sci. USA 2004, 101, 8039–8044. [Google Scholar] [CrossRef] [Green Version]
- Ioio, R.D.; Nakamura, K.; Moubayidin, L.; Perilli, S.; Taniguchi, M.; Morita, M.T.; Aoyama, T.; Costantino, P.; Sabatini, S. A Genetic Framework for the Control of Cell Division and Differentiation in the Root Meristem. Science 2008, 322, 1380–1384. [Google Scholar] [CrossRef] [Green Version]
- Rashotte, A.M.; Carson, S.D.B.; To, J.P.C.; Kieber, J.J. Expression profiling of cytokinin action in Arabidopsis. Plant Physiol. 2003, 132, 1998–2011. [Google Scholar] [CrossRef] [Green Version]
- Kaur, K.; Verma, B.; Kant, U. Plants obtained from the Khair tree (Acacia catechu Willd.) using mature nodal segments. Plant Cell Rep. 1998, 17, 427–429. [Google Scholar] [CrossRef]
- Barrueto Cid, L.P.; Machado, A.C.M.G.; Carvalheira, S.B.R.C.; Brasileiro, A.C.M. Plant regeneration from seedling explants of Eucalyptus grandis × E. urophylla. Plant Cell Tissue Organ Cult. 1999, 56, 17–23. [Google Scholar] [CrossRef]
- Tiwari, S.K.; Kashyap, M.K.; Ujjaini, M.M.; Agrawal, A.P. In vitro propagation of Lagerstromia parviflora Roxb. from adult tree. Indian J. Exp. Biol. 2002, 40, 212–215. [Google Scholar]
- Siddique, I.; Anis, M. Direct plant regeneration from nodal explants of Balanites aegyptiaca L. (Del.): A valuable medicinal tree. New For. 2009, 37, 53–62. [Google Scholar] [CrossRef]
- Devi, Y. Rapid cloning of elite teak (Tectona grandis Linn.) by In vitro multiple shoot production. Indian J. Exp. Biol. 1994, 32, 668–671. [Google Scholar]
- Dhar, U.; Upreti, J. In vitro regeneration of a mature leguminous liana (Bauhinia vahlii Wight & Arnott). Plant Cell Rep. 1999, 18, 664–669. [Google Scholar] [CrossRef]
- Husain, M.K.; Anis, M. In vitro proliferation of shoots of Melia azedarach L. from mature trees. In Biotechnology for a Better Future; D’Souza, L., Anuradha, M., Nivas, S., Hegde, S., Rajendra, K., Eds.; SAC: Mangalore, India, 2004; pp. 294–301. [Google Scholar]
- Giri, C.C.; Shyamkumar, B.; Anjaneyulu, C. Progress in tissue culture, genetic transformation and applications of biotechnology to trees: An overview. Trees 2004, 18, 115–135. [Google Scholar] [CrossRef]
- Fuentes, S.R.L.; Calheiros, M.B.P.; Manetti-Filho, J.; Vieira, L.G.E. The effects of silver nitrate and different carbohydrate sources on somatic embryogenesis in Coffea canephora. Plant Cell Tissue Organ Cult. 2000, 60, 5–13. [Google Scholar] [CrossRef]
- Sinha, R.K.; Majumdar, K.; Sinha, S. Somatic embryogenesis and plantlet regeneration from leaf explants of Sapindus mukorossi Gaertn.: A soapnut tree. Curr. Sci. 2000, 78, 620–623. [Google Scholar]
- Nehra, N.S.; Kartha, K.K. Meristem and Shoot Tip Culture: Requirements and Applications. In Plant Cell and Tissue Culture; Vasil, I.K., Thorpe, T.A., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 37–70. [Google Scholar]
- Sharma, A.K.; Prasad, R.N.; Chaturvedi, H.C. Clonal propagation of Bougainvillea glabra ‘Magnifica’ through shoot apex culture. Plant Cell Tissue Organ Cult. 1981, 1, 33–38. [Google Scholar] [CrossRef]
- Hamid, M.E. In vitro Propagation of Catha edulis. HortScience 1991, 26, 212. [Google Scholar] [CrossRef] [Green Version]
- Kaur, K.; Kant, U. Clonal propagation of Acacia catechu Willd. by shoot tip culture. Plant Growth Regul. 2000, 31, 143–145. [Google Scholar] [CrossRef]
- Balaraju, K.; Agastian, P.; Ignacimuthu, S.; Park, K. A rapid in vitro propagation of red sanders (Pterocarpus santalinus L.) using shoot tip explants. Acta Physiol. Plant. 2011, 33, 2501. [Google Scholar] [CrossRef]
- Al-Sulaiman, M.A.; Barakat, M.N. In vitro shoot multiplication of Ziziphus spina-christi by shoot tip culture. Afr. J. Biotechnol. 2010, 9, 850–857. [Google Scholar]
- Hussain, S.A.; Anis, M.; Alatar, A.A. Efficient In Vitro Regeneration System for Tecoma stans L., Using Shoot Tip and Assessment of Genetic Fidelity Among Regenerants. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2020, 90, 171–178. [Google Scholar] [CrossRef]
- Borges de Paiva Neto, V.; Ribeiro da Mota, T.; Campos Otoni, W. Direct organogenesis from hypocotyl-derived explants of annatto (Bixa orellana). Plant Cell Tissue Organ Cult. 2003, 75, 159–167. [Google Scholar] [CrossRef]
- Taniguchi, T.; Kurita, M.; Itahana, N.; Kondo, T. Somatic embryogenesis and plant regeneration from immature zygotic embryos of Hinoki cypress (Chamaecyparis obtusa Sieb. et Zucc.). Plant Cell Rep. 2004, 23, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Lara-Chavez, A.; Flinn, B.S.; Egertsdotter, U. Initiation of somatic embryogenesis from immature zygotic embryos of Oocarpa pine (Pinus oocarpa Schiede ex Schlectendal). Tree Physiol. 2011, 31, 539–554. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Wu, Y.; Shen, Y.; Mao, Y.; Liu, W.; Zhao, W.; Mu, Y.; Li, M.; Yang, M.; Wu, L. Plant regeneration through somatic embryogenesis and shoot organogenesis from immature zygotic embryos of Sapium sebiferum Roxb. Sci. Hortic. 2015, 197, 218–225. [Google Scholar] [CrossRef]
- Dunstan, D.I.; Tautorus, T.E.; Thorpe, T.A. Somatic embryogenesis in woody plants. In In Vitro Embryogenesis in Plants; Thorpe, T.A., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995. [Google Scholar]
- Apóstolo, N.M.; Brutti, C.B.; Llorente, B.E. Leaf anatomy of Cynara scolymus L. in successive micropropagation stages. In Vitr. Cell. Dev. Biol.-Plant 2005, 41, 307–313. [Google Scholar] [CrossRef]
- Fatima, N.; Ahmad, N.; Ahmad, I.; Anis, M. Interactive Effects of Growth Regulators, Carbon Sources, pH on Plant Regeneration and Assessment of Genetic Fidelity Using Single Primer Amplification Reaction (SPARS) Techniques in Withania somnifera L. Appl. Biochem. Biotechnol. 2015, 177, 118–136. [Google Scholar] [CrossRef] [PubMed]
- Hiregoudar, L.V.; Kumar, H.G.; Murthy, H.N. In vitro culture of Feronia limonia (L.) Swingle from hypocotyl and internodal explants. Biol. Plant. 2005, 49, 41–45. [Google Scholar] [CrossRef]
- Prakash, E.; Khan, P.S.S.V.; Rao, T.J.V.S.; Meru, E.S. Micropropagation of red sanders (Pterocarpus santalinus L.) using mature nodal explants. J. For. Res. 2006, 11, 329–335. [Google Scholar] [CrossRef]
- Tripathi, M.; Kumari, N. Micropropagation of a tropical fruit tree Spondias mangifera Willd. through direct organogenesis. Acta Physiol. Plant. 2010, 32, 1011–1015. [Google Scholar] [CrossRef]
- Shekhawat, M.S.; Shekhawat, N.S. Micropropagation of Arnebia hispidissima (Lehm). DC. and production of alkannin from callus and cell suspension culture. Acta Physiol. Plant. 2010, 33, 1445–1450. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, T.-t.; Zhang, J. Effect of subculture times on genetic fidelity, endogenous hormone level and pharmaceutical potential of Tetrastigma hemsleyanum callus. Plant Cell Tissue Organ Cult. 2015, 122, 67–77. [Google Scholar] [CrossRef]
- Malik, K.; Saxena, P. Regeneration in Phaseolus vulgaris L.: High-frequency induction of direct shoot formation in intact seedlings by N6-benzylaminopurine and thidiazuron. Planta 1992, 186, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Bhuyan, A.K.; Pattnaik, S.; Chand, P.K. Micropropagation of Curry Leaf Tree [Murraya koenigii (L.) Spreng.] by axillary proliferation using intact seedlings. Plant Cell Rep. 1997, 16, 779–782. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.M.; Chandrasekhar, T.; Gopal, G.R. Micropropagation of Sterculia urens Roxb., an endangered tree species from intact seedlings. Afr. J. Biotechnol. 2008, 7, 95–101. [Google Scholar]
- Williams, E.G.; Maheswaran, G. Somatic Embryogenesis: Factors Influencing Coordinated Behaviour of Cells as an Embryogenic Group. Ann. Bot. 1986, 57, 443–462. [Google Scholar] [CrossRef] [Green Version]
- Chaturani, G.; Subasinghe, S.; Jayatilleke, M.; Mapalana, K.; Lanka, S. In vitro establishment, germination and growth performance of Red sandal wood (Pterocarpus santalinus L.). Trop. Agric. Res. Ext. 2006, 9, 116–130. [Google Scholar]
- Yang, L.; Li, Y.; Shen, H. Somatic embryogenesis and plant regeneration from immature zygotic embryo cultures of mountain ash (Sorbus pohuashanensis). Plant Cell Tissue Organ Cult. 2012, 109, 547–556. [Google Scholar] [CrossRef]
- Ashok Kumar, H.G.; Murthy, H.N.; Paek, K.Y. Somatic embryogenesis and plant regeneration in Gymnema sylvestre. Plant Cell Tissue Organ Cult. 2002, 71, 85–88. [Google Scholar] [CrossRef]
- Junaid, A.; Mujib, A.; Bhat, M.A.; Sharma, M.P. Somatic embryo proliferation, maturation and germination in Catharanthus roseus. Plant Cell Tissue Organ Cult. 2006, 84, 325–332. [Google Scholar] [CrossRef]
- Rout, G.R.; Samantaray, S.; Das, P. Somatic embryogenesis and plant regeneration from callus culture of Acacia catechu—A multipurpose leguminous tree. Plant Cell Tissue Organ Cult. 1995, 42, 283–285. [Google Scholar] [CrossRef]
- Das, A.B.; Rout, G.R.; Das, P. In vitro somatic embryogenesis from callus culture of the timber yielding tree Hardwickia binata Roxb. Plant Cell Rep. 1995, 15, 147–149. [Google Scholar] [CrossRef]
- Garg, L.; Bhandari, N.N.; Rani, V.; Bhojwani, S.S. Somatic embryogenesis and regeneration of triploid plants in endosperm cultures of Acacia nilotica. Plant Cell Rep. 1996, 15, 855–858. [Google Scholar] [CrossRef]
- Austin Burns, J.; Wetzstein, H.Y. Embryogenic cultures of the leguminous tree Albizia julibrissin and recovery of plants. Plant Cell Tissue Organ Cult. 1998, 54, 55–59. [Google Scholar] [CrossRef]
- Xie, D.Y.; Hong, Y. Regeneration of Acacia mangium through somatic embryogenesis. Plant Cell Rep. 2001, 20, 34–40. [Google Scholar] [CrossRef]
- De Klerk, G.-J. Rooting of microcuttings: Theory and practice. In Vitr. Cell. Dev. Biol.-Plant 2002, 38, 415–422. [Google Scholar] [CrossRef]
- Wiszniewska, A.; Nowak, B.; Kołton, A.; Sitek, E.; Grabski, K.; Dziurka, M.; Długosz-Grochowska, O.; Dziurka, K.; Tukaj, Z. Rooting response of Prunus domestica L. microshoots in the presence of phytoactive medium supplements. Plant Cell Tissue Organ Cult. 2016, 125, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Ilczuk, A.; Jacygrad, E. The effect of IBA on anatomical changes and antioxidant enzyme activity during the in vitro rooting of smoke tree (Cotinus coggygria Scop.). Sci. Hortic. 2016, 210, 268–276. [Google Scholar] [CrossRef]
- Yan, Y.-H.; Li, J.-L.; Zhang, X.-Q.; Yang, W.-Y.; Wan, Y.; Ma, Y.-M.; Zhu, Y.-Q.; Peng, Y.; Huang, L.-K. Effect of naphthalene acetic acid on adventitious root development and associated physiological changes in stem cutting of Hemarthria compressa. PLoS ONE 2014, 9, e90700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naija, S.; Elloumi, N.; Jbir, N.; Ammar, S.; Kevers, C. Anatomical and biochemical changes during adventitious rooting of apple rootstocks MM 106 cultured in vitro. Comptes Rendus Biol. 2008, 331, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-Y.; Guo, Z.-R.; Zhang, R.; Li, Y.-R.; Cao, L.; Liang, Y.-W.; Huang, L.-B. Auxin Type, Auxin Concentration, and Air and Substrate Temperature Difference Play Key Roles in the Rooting of Juvenile Hardwood Pecan Cuttings. HortTechnology 2015, 25, 209–213. [Google Scholar] [CrossRef] [Green Version]
- Elmongy, M.S.; Cao, Y.; Zhou, H.; Xia, Y. Root Development Enhanced by Using Indole-3-butyric Acid and Naphthalene Acetic Acid and Associated Biochemical Changes of In Vitro Azalea Microshoots. J. Plant Growth Regul. 2018, 37, 813–825. [Google Scholar] [CrossRef]
- Ludwig-Müller, J. Indole-3-butyric acid in plant growth and development. Plant Growth Regul. 2000, 32, 219–230. [Google Scholar] [CrossRef]
- Rajeswari, V.; Paliwal, K. In vitro propagation of Albizia odoratissima L.F. (Benth.) from cotyledonary node and leaf nodal explants. In Vitr. Cell. Dev. Biol.-Plant 2006, 42, 399–404. [Google Scholar] [CrossRef]
- Werbrouck, S.P.O.; van der Jeugt, B.; Dewitte, W.; Prinsen, E.; Van Onckelen, H.A.; Debergh, P.C. The metabolism of benzyladenine in Spathiphyllum floribundum ‘Schott Petite’ in relation to acclimatisation problems. Plant Cell Rep. 1995, 14, 662–665. [Google Scholar] [CrossRef]
- Khanam, M.N.; Javed, S.B.; Anis, M.; Alatar, A.A. meta-Topolin induced In vitro regeneration and metabolic profiling in Allamanda cathartica L. Ind. Crops Prod. 2020, 145, 111944. [Google Scholar] [CrossRef]
- Khanam, M.N.; Anis, M. Organogenesis and efficient in vitro plantlet regeneration from nodal segments of Allamanda cathartica L. using TDZ and ultrasound assisted extraction of quercetin. Plant Cell Tissue Organ Cult. 2018, 134, 241–250. [Google Scholar] [CrossRef]
- Hussain, S.A.; Ahmad, N.; Anis, M.; Hakeem, K.R. Development of an efficient micropropagation system for Tecoma stans (L.) Juss. ex Kunth using thidiazuron and effects on phytochemical constitution. In Vitr. Cell. Dev. Biol.-Plant 2019, 55, 442–453. [Google Scholar] [CrossRef]
- Du, Y.; Scheres, B. Lateral root formation and the multiple roles of auxin. J. Exp. Bot. 2018, 69, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Ilina, E.L.; Kiryushkin, A.S.; Semenova, V.A.; Demchenko, N.P.; Pawlowski, K.; Demchenko, K.N. Lateral root initiation and formation within the parental root meristem of Cucurbita pepo: Is auxin a key player? Ann. Bot. 2018, 122, 873–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Smet, I. Lateral root initiation: One step at a time. New Phytol. 2012, 193, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Lloret, P.G.; Casero, P.J. Lateral root initiation. In Plant Roots—The Hidden Half, 3rd ed.; Waisel, Y., Eshel, A., Kafkafi, U., Eds.; Marcel Dekker Inc.: New York, NY, USA, 2002; pp. 127–156. [Google Scholar]
- Demchenko, N.P.; Demchenko, K.N. Resumption of DNA Synthesis and Cell Division in Wheat Roots as Related to Lateral Root Initiation. Russ. J. Plant Physiol. 2001, 48, 755–764. [Google Scholar] [CrossRef]
- Vieten, A.; Vanneste, S.; Wisniewska, J.; Benkova, E.; Benjamins, R.; Beeckman, T.; Luschnig, C.; Friml, J. Functional redundancy of PIN proteins is accompanied by auxin-dependentcross-regulation of PIN expression. Development 2005, 132, 4521–4531. [Google Scholar] [CrossRef] [Green Version]
- De Klerk, G.-J.; Brugge, J.T.; Marinova, S. Effectiveness of indoleacetic acid, indolebutyric acid and naphthaleneacetic acid during adventitious root formation in vitro in Malus ‘Jork 9′. Plant Cell Tissue Organ Cult. 1997, 49, 39–44. [Google Scholar] [CrossRef]
- Thomas, T.D.; Puthur, J.T. Thidiazuron induced high frequency shoot organogenesis in callus from Kigelia pinnata L. Bot. Bull. Acad. Sin 2004, 45, 307–313. [Google Scholar]
- Pandey, S.; Singh, M.; Jaiswal, U.; Jaiswal, V.S. Shoot initiation and multiplication from a mature tree of Terminalia arjuna Roxb. In Vitr. Cell. Dev. Biol.-Plant 2006, 42, 389–393. [Google Scholar] [CrossRef]
- James, D.J. Adventitious root formation ‘in vitro’ in apple rootstocks (Malus pumila) I. Factors affecting the length of the auxin-sensitive phase in M.9. Physiol. Plant. 1983, 57, 149–153. [Google Scholar] [CrossRef]
- Zanol, G.; Fortes, G.d.L.; Campos, A.; Da Silva, J.; Centellas, A. In vitro rooting and peroxidase activity of apple rootstocks cv.” Marubakaido” treated with indolebutyric acid and floroglucinol. Rev. Bras. De Fisiol. Veg. 1998, 10, 65–68. [Google Scholar]
- Hammatt, N.; Grant, N.J. Micropropagation of mature British wild cherry. Plant Cell Tissue Organ Cult. 1997, 47, 103–110. [Google Scholar] [CrossRef]
- De Klerk, G.-J.; van der Krieken, W.; De Jong, J.C. Review the formation of adventitious roots: New concepts, new possibilities. In Vitr. Cell. Dev. Biol.-Plant 1999, 35, 189–199. [Google Scholar] [CrossRef]
- Borkowska, B. Morphological and physiological characteristics of micropropagated strawberry plants rooted in vitro or ex vitro. Sci. Hortic. 2001, 89, 195–206. [Google Scholar] [CrossRef]
- Sharma, S.; Gambhir, G.; Srivastava, D.K. In vitro differentiation and plant regeneration from root and other explants of juvenile origin in pea (Pisum sativum L.). Legume Res. 2017, 40, 1020–1027. [Google Scholar] [CrossRef] [Green Version]
- Bellamine, J.; Penel, C.; Greppin, H.; Gaspar, T. Confirmation of the role of auxin and calcium in the late phases of adventitious root formation. Plant Growth Regul. 1998, 26, 191–194. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Z. Micropropagation of Camptotheca acuminata decaisne from axillary buds, shoot tips, and seed embryos in a tissue culture system. In Vitr. Cell. Dev. Biol.-Plant 2001, 37, 84–88. [Google Scholar] [CrossRef]
- Yan, H.; Liang, C.; Yang, L.; Li, Y. In vitro and ex vitro rooting of Siratia grosvenorii, a traditional medicinal plant. Acta Physiol. Plant. 2010, 32, 115–120. [Google Scholar] [CrossRef]
- Tiwari, S.K.; Tiwari, K.P.; Siril, E.A. An improved micropropagation protocol for teak. Plant Cell Tissue Organ Cult. 2002, 71, 1–6. [Google Scholar] [CrossRef]
- Siddique, I.; Anis, M.; Jahan, A. Rapid multiplication of Nyctanthes arbor-tristis L. through In vitro axillary shoot proliferation. World J. Agri. Sci 2006, 2, 188–192. [Google Scholar]
- Xu, J.; Wang, Y.; Zhang, Y.; Chai, T. Rapid In vitro multiplication and ex vitro rooting of Malus zumi (Matsumura) Rehd. Acta Physiol. Plant. 2007, 30, 129–132. [Google Scholar] [CrossRef]
- Varshney, A.; Anis, M. Improvement of shoot morphogenesis in vitro and assessment of changes of the activity of antioxidant enzymes during acclimation of micropropagated plants of Desert Teak. Acta Physiol. Plant. 2012, 34, 859–867. [Google Scholar] [CrossRef]
- Alam, N.; Anis, M.; Javed, S.B.; Alatar, A.A. Stimulatory effect of copper and zinc sulphate on plant regeneration, glutathione-S-transferase analysis and assessment of antioxidant activities in Mucuna pruriens L. (DC). Plant Cell Tissue Organ Cult. 2020, 141, 155–166. [Google Scholar] [CrossRef]
- Amâncio, S.; Rebordão, J.P.; Chaves, M.M. Improvement of acclimatization of micropropagated grapevine: Photosynthetic competence and carbon allocation. Plant Cell Tissue Organ Cult. 1999, 58, 31–37. [Google Scholar] [CrossRef]
- Kadleček, P.; Tichá, I.; Čapková, V.; Schäfer, C. Acclimatization of Micropropagated Tobacco Plantlets. In Photosynthesis: Mechanisms and Effects: Volume I–V, Proceedings of the XIth International Congress on Photosynthesis, Budapest, Hungary, 17–22 August 1998; Garab, G., Ed.; Springer: Dordrecht, The Netherlands, 1998; pp. 3853–3856. [Google Scholar]
- Pospisilova, J.; Ticha, I.; Kadlecek, P.; Haisel, D.; Plzakova, S. Acclimatization of Micropropagated Plants to Ex Vitro Conditions. Biol. Plant. 1999, 42, 481–497. [Google Scholar] [CrossRef]
- Yang, Y.-J.; Tong, Y.-G.; Yu, G.-Y.; Zhang, S.-B.; Huang, W. Photosynthetic characteristics explain the high growth rate for Eucalyptus camaldulensis: Implications for breeding strategy. Ind. Crops Prod. 2018, 124, 186–191. [Google Scholar] [CrossRef]
- Lavanya, M.; Venkateshwarlu, B.; Devi, B.P. Acclimatization of neem microshoots adaptable to semi-sterile conditions. Indian J. Biotechnol. 2009, 8, 218–222. [Google Scholar]
- Faisal, M.; Anis, M. Effect of light irradiations on photosynthetic machinery and antioxidative enzymes during ex vitro acclimatization of Tylophora indica plantlets. J. Plant Interact. 2010, 5, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Nayak, S.A.; Kumar, S.; Satapathy, K.; Moharana, A.; Behera, B.; Barik, D.P.; Acharya, L.; Mohapatra, P.K.; Jena, P.K.; Naik, S.K. In vitro plant regeneration from cotyledonary nodes of Withania somnifera (L.) Dunal and assessment of clonal fidelity using RAPD and ISSR markers. Acta Physiol. Plant. 2013, 35, 195–203. [Google Scholar] [CrossRef]
- Javed, S.B.; Alatar, A.A.; Anis, M.; Faisal, M. Synthetic seeds production and germination studies, for short term storage and long distance transport of Erythrina variegata L.: A multipurpose tree legume. Ind. Crops Prod. 2017, 105, 41–46. [Google Scholar] [CrossRef]
- Goyal, A.K.; Pradhan, S.; Basistha, B.C.; Sen, A. Micropropagation and assessment of genetic fidelity of Dendrocalamus strictus (Roxb.) nees using RAPD and ISSR markers. 3 Biotech 2015, 5, 473–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Khateeb, W.; Bahar, E.; Lahham, J.; Schroeder, D.; Hussein, E. Regeneration and assessment of genetic fidelity of the endangered tree Moringa peregrina (Forsk.) Fiori using Inter Simple Sequence Repeat (ISSR). Physiol. Mol. Biol. Plants 2013, 19, 157–164. [Google Scholar] [CrossRef]
- Agarwal, T.; Gupta, A.K.; Patel, A.K.; Shekhawat, N.S. Micropropagation and validation of genetic homogeneity of Alhagi maurorum using SCoT, ISSR and RAPD markers. Plant Cell Tissue Organ Cult. 2015, 120, 313–323. [Google Scholar] [CrossRef]
- Dangi, B.; Khurana-Kaul, V.; Kothari, S.L.; Kachhwaha, S. Micropropagtion of Terminalia bellerica from nodal explants of mature tree and assessment of genetic fidelity using ISSR and RAPD markers. Physiol. Mol. Biol. Plants 2014, 20, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Adhikari, S.; Dey, T.; Ghosh, P. RAPD and ISSR based evaluation of genetic stability of micropropagated plantlets of Morus alba L. variety S-1. Meta Gene 2016, 7, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Kuzminsky, E.; Alicandri, E.; Agrimi, M.; Vettraino, A.M.; Ciaffi, M. Data set useful for the micropropagation and the assessment of post-vitro genetic fidelity of veteran trees of P. orientalis L. Data Brief 2018, 20, 1532–1536. [Google Scholar] [CrossRef] [PubMed]
- Antony Ceasar, S.; Lenin Maxwell, S.; Bhargav Prasad, K.; Karthigan, M.; Ignacimuthu, S. Highly efficient shoot regeneration of Bacopa monnieri (L.) using a two-stage culture procedure and assessment of genetic integrity of micropropagated plants by RAPD. Acta Physiol. Plant. 2010, 32, 443–452. [Google Scholar] [CrossRef]
- Gupta, A.K.; Harish; Rai, M.K.; Phulwaria, M.; Agarwal, T.; Shekhawat, N.S. In vitro Propagation, Encapsulation, and Genetic Fidelity Analysis of Terminalia arjuna: A Cardioprotective Medicinal Tree. Appl. Biochem. Biotechnol. 2014, 173, 1481–1494. [Google Scholar] [CrossRef]
- Saeed, T.; Shahzad, A.; Ahmad, N.; Parveen, S. High frequency conversion of non-embryogenic synseeds and assessment of genetic stability through ISSR markers in Gymnema sylvestre. Plant Cell Tissue Organ Cult. 2018, 134, 163–168. [Google Scholar] [CrossRef]
- Jiao, L.; Yin, Y.; Cheng, Y.; Jiang, X. DNA barcoding for identification of the endangered species Aquilaria sinensis: Comparison of data from heated or aged wood samples. Holzforschung 2014, 68, 487–494. [Google Scholar] [CrossRef]
- Hajibabaei, M.; Singer, G.A.C.; Hebert, P.D.N.; Hickey, D.A. DNA barcoding: How it complements taxonomy, molecular phylogenetics and population genetics. Trends Genet. 2007, 23, 167–172. [Google Scholar] [CrossRef]
- Lee, S.Y.; Ng, W.L.; Mahat, M.N.; Nazre, M.; Mohamed, R. DNA Barcoding of the Endangered Aquilaria (Thymelaeaceae) and Its Application in Species Authentication of Agarwood Products Traded in the Market. PLoS ONE 2016, 11, e0154631. [Google Scholar] [CrossRef]
- Chen, S.; Yao, H.; Han, J.; Liu, C.; Song, J.; Shi, L.; Zhu, Y.; Ma, X.; Gao, T.; Pang, X.; et al. Validation of the ITS2 Region as a Novel DNA Barcode for Identifying Medicinal Plant Species. PLoS ONE 2010, 5, e8613. [Google Scholar] [CrossRef] [PubMed]
- Timme, R.E.; Kuehl, J.V.; Boore, J.L.; Jansen, R.K. A comparative analysis of the Lactuca and Helianthus (Asteraceae) plastid genomes: Identification of divergent regions and categorization of shared repeats. Am. J. Bot. 2007, 94, 302–312. [Google Scholar] [CrossRef]
- Shaw, J.; Lickey, E.B.; Schilling, E.E.; Small, R.L. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: The tortoise and the hare III. Am. J. Bot. 2007, 94, 275–288. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Dong, W.; Liu, B.; Xu, C.; Yao, X.; Gao, J.; Corlett, R.T. Comparative analysis of complete chloroplast genome sequences of two tropical trees Machilus yunnanensis and Machilus balansae in the family Lauraceae. Front. Plant Sci. 2015, 6, 662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, T.; Teng, R.; Shao, Q.; Wang, H.; Zhang, W.; Li, M.; Zhang, L. DNA barcodes for the identification of Anoectochilus roxburghii and its adulterants. Planta 2015, 242, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Pei, N.; Chen, B.; Kress, W.J. Advances of Community-Level Plant DNA Barcoding in China. Front. Plant Sci. 2017, 8, 225. [Google Scholar] [CrossRef] [Green Version]
- Li, D.-Z.; Gao, L.-M.; Li, H.-T.; Wang, H.; Ge, X.-J.; Liu, J.-Q.; Chen, Z.-D.; Zhou, S.-L.; Chen, S.-L.; Yang, J.-B.; et al. Comparative analysis of a large dataset indicates that internal transcribed spacer (ITS) should be incorporated into the core barcode for seed plants. Proc. Natl. Acad. Sci. USA 2011, 108, 19641–19646. [Google Scholar] [CrossRef] [Green Version]
- Campbell, M.M.; Brunner, A.M.; Jones, H.M.; Strauss, S.H. Forestry’s fertile crescent: The application of biotechnology to forest trees. Plant Biotechnol. J. 2003, 1, 141–154. [Google Scholar] [CrossRef]
- Yang, M.; Xie, X.; Zheng, C.; Zhang, F.; He, X.; Li, Z. Agrobacterium tumefaciens-mediated genetic transformation of Acacia crassicarpa via organogenesis. Plant Cell Tissue Organ Cult. 2008, 95, 141–147. [Google Scholar] [CrossRef]
- Thangjam, R.; Sahoo, L. In vitro regeneration and Agrobacterium tumefaciens-mediated genetic transformation of Parkia timoriana (DC.) Merr.: A multipurpose tree legume. Acta Physiol. Plant. 2012, 34, 1207–1215. [Google Scholar] [CrossRef]
- Gorpenchenko, T.Y.; Kiselev, K.V.; Bulgakov, V.P.; Tchernoded, G.K.; Bragina, E.A.; Khodakovskaya, M.V.; Koren, O.G.; Batygina, T.B.; Zhuravlev, Y.N. The Agrobacterium rhizogenes rolC-gene-induced somatic embryogenesis and shoot organogenesis in Panax ginseng transformed calluses. Planta 2006, 223, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Jube, S.; Borthakur, D. Development of an Agrobacterium-mediated transformation protocol for the tree-legume Leucaena leucocephala using immature zygotic embryos. Plant Cell Tissue Organ Cult. 2009, 96, 325–333. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Parts | Extract Preparation | Technique * | Bioactive Compound | References |
|---|---|---|---|---|
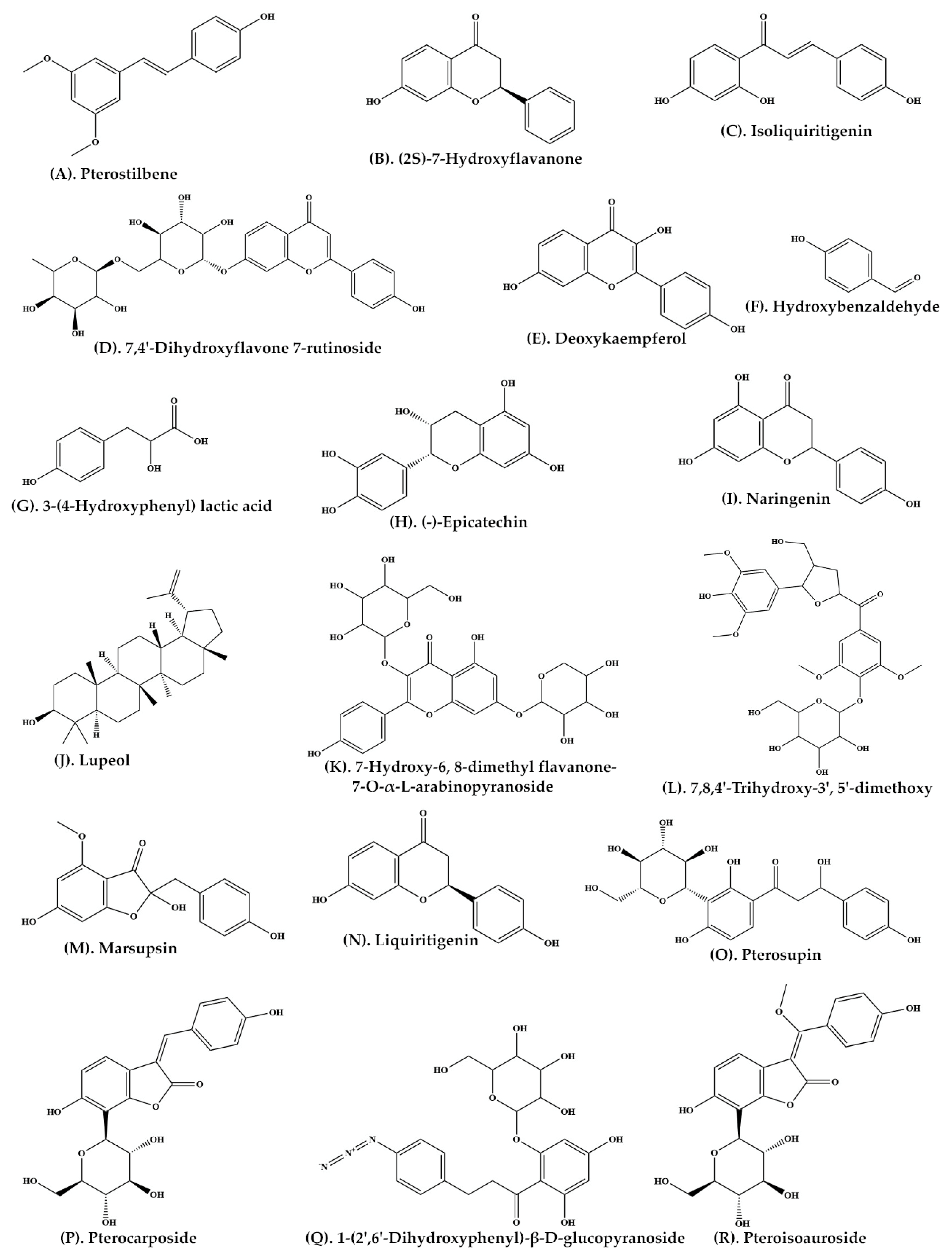
| Heartwood | Ethyl acetate | C-SG | Pterostilbene (Figure 2A) (2S)-7-Hydroxyflavanone (Figure 2B) Isoliquiritigenin (Figure 2C) 7,4′-Dihydroxyflavone 7-rutinoside (Figure 2D) 5-Deoxykaempferol (Figure 2E) p-Hydroxybenzaldehyde (Figure 2F) 3-(4-Hydroxyphenyl) lactic acid (Figure 2G) | [34] |
| Bark | Ethanolic extract | C-SG | (−)-Epicatechin (Figure 2H) | [35] |
| P. marsupium extract | Ethyl acetate | C-SG | Naringenin (Figure 2I) Lupeol (Figure 2J) | [36] |
| Roots | Ethanolic extract | C-SG | 7-Hydroxy-6, 8-dimethyl flavanone-7-O-α-L-arabinopyranoside (Figure 2K) 7,8,4′-Trihydroxy-3′, 5′-dimethoxy flavanone-4′-O-β-D-glucopyranoside (Figure 2L) | [37] |
| Heartwood | Ethyl acetate | Thin Layer Chromatography | Marsupsin (Figure 2M) Liquiritigenin (Figure 2N) | [31] |
| Heartwood | Ethyl acetate | C-SG | Pterosupin (Figure 2O) | [32] |
| Heartwood | Aqueous extract | C-SG | Pterocarposide (Figure 2P) | [38] |
| Heartwood | Aqueous extract | Coulman chromatography over Sephadex LH-20 | 1-(2′,6′-Dihydroxyphenyl)-β-D-glucopyranoside (Figure 2Q) | [39] |
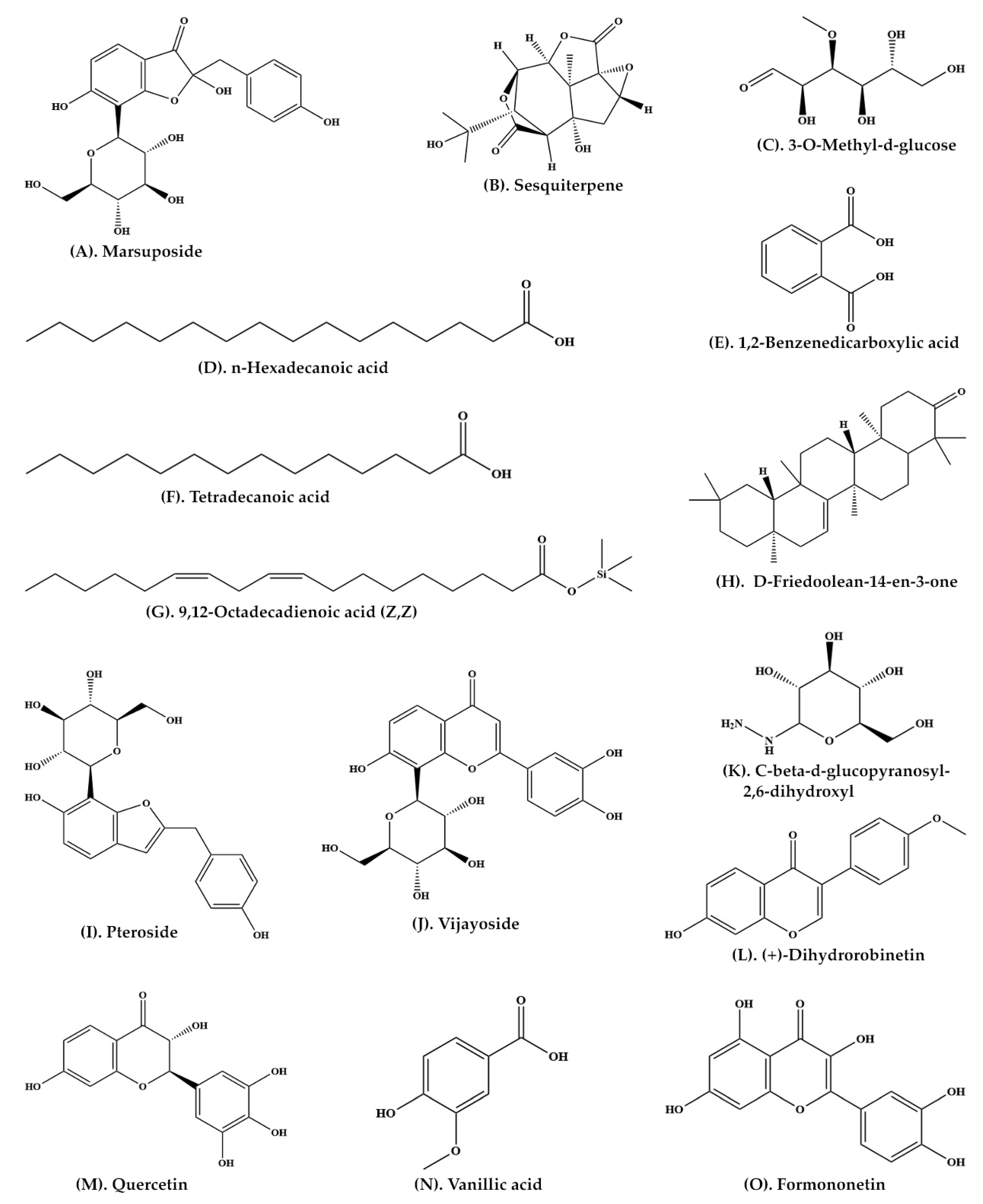
| Heartwood | Aqueous extract | C-SG | Pteroisoauroside (Figure 2R) Marsuposide (Figure 3A) Sesquiterpene (Figure 3B) | [30] |
| Leaves | Methanolic extract | UV-spectrophotometer | Phenolics | [40] |
| Wood and bark | Ethanolic extract | GC-MS | 3-O-Methyl-d-glucose (Figure 3C) n-Hexadecanoic acid (Figure 3D) 1,2-Benzenedicarboxylic acid (Figure 3E) Tetradecanoic acid (Figure 3F) 9,12-Octadecadienoic acid (Z,Z) (Figure 3G) D-Friedoolean-14-en-3-one (Figure 3H) | [33] |
| Apical stem bark | Methanolic extract | Followed standard protocols | Alkaloids Glycosides Flavonoids Terpenoids | [41] |
| Heartwood | Ethanolic extract | C-SG | Pteroside (Figure 3I) Vijayoside (Figure 3J) C-β-D-Glucopyranosyl-2,6-dihydroxyl benzene (Figure 3K) | [42] |
| Heartwood | Ethanolic extract | C-SG and HPLC | (+)-Dihydrorobinetin (Figure 3L) | [43] |
| Heartwood | Methanolic extract | LC-MS-MS | Pterosupol Quercetin (Figure 3M) Vanillic acid (Figure 3N) Formononetin (Figure 3O) | [21] |
| Heartwood | Methanolic extract | HPLC and FTIR | Liquiritigenin | [44] |
| S.N. | Extracts/Bioactive Compound | Potential Activities | References |
|---|---|---|---|
| 1 | (−)-Epicatechin (Figure 2H) | No effect on central nervous system Cardiac stimulant activity Anti-diabetic | [35] |
| 2 | Flavonoids | Anti-hyperlipidemic | [31] |
| 3 | Phenolics | Anti-hyperglycemic | [32] |
| 4 | Pterostilbene (Figure 2A) | Cyclooxygenase-2 (COX-2) inhibition | [52] |
| 5 | Pterostilbene and 3,5-hydroxypterostilbene | Induce apoptosis in tumor cells | [53] |
| 6 | 5,7,2-4 tetrahydroxy isoflavone 6-6 glucoside | Cardiotonic | [45] |
| 7 | Pterostilbene | Anti-cancerous Anti-inflammatory Analgesic | [54] |
| 8 | Phenolics | Anti-oxidant | [40] |
| 9 | Pterostilbene | Anti-cancerous Anti-proliferative | [55] |
| 10 | Bark extract | Anti-oxidant Analgesic | [56] |
| 11 | Extract of bark and wood | Anti-diabetic Anti-hyperlipidemic | [57] |
| 12 | Extract of apical stem bark | Anti-microbicidal | [41] |
| 13 | Phenolic-C-glycosides | Anti-diabetic | [42] |
| 14 | Pterostilbene | Novel telomerase inhibitor | [58] |
| 15 | Heartwood extract | Dipeptidyl peptidase-4 (DPP-4) inhibition activity | [59] |
| 16 | Heartwood extract | Anti-glycation Sorbitol accumulation Inhibition of aldose reductase | [60] |
| 17 | Pterostilbene | Inhibition of platelet aggregation | [61] |
| 18 | Heartwood extract | Reduction in body weight Anti-diabetic Anti-hyperlipidemic | [62] |
| 19 | (+)-Dihydrorobinetin (Figure 3L) | Radical scavenging activity | [43] |
| 20 | Heartwood extract | In vitro lipid lowering activity | [21] |
| 21 | Liquiritigenin (Figure 2N) | Hypoglycemic activity | [44] |
| 22 | Pterostilbene | Sun (UV rays) protective capacity | [63] |
| Explants | Source | Media Compositions (Multiplication) * | Culture Response | Media Compositions (Rhizogenesis) | Rooting Response | Plantlets Survival Rate | References |
|---|---|---|---|---|---|---|---|
| Shoot tip | AS/MT | MS + 0.2 mg·L−1 BAP | Ca-Dm | - | - | - | [96] |
| Aseptic seeds | - | MS basal medium | ISG (95–100%) | - | - | >68% | [26] |
| Nodal segment | 35-d-old-AS | MS + 0.2 mg·L−1 IBA | IO | MS + 0.2 mg·L−1 IBA | RF | ||
| Cotyledonary node | 20-d-old-AS | MS + 4.44 µM BA+ 0.26 µM NAA | SM (85%) | ½ MS + 9.84 µM IBA | RF (IVR) | 52% | [136] |
| Cotyledonary node | 18-d-old-AS | MS + 5.0 µM BA + 0.25 µM IAA | SM (75%) | 2-step-method: PT on ½ MS (liquid) + 200 µM IBA | MRI (40–50%) | - | [25] |
| FT on ½ MS (semi-solid) + 0.5 µM IBA | ER (IVR) | ||||||
| Aseptic seeds | - | ½ MS 0.25 mg·L−1 GA3 | ISG (80%) | - | - | - | [98] |
| Cotyledonary node | 18-d-old-AS | 2-step-method: SIM: MS + 0.4 µM TDZ | MSI (90%) | 2-step-method: PT on ½ MS (liquid) + 200 µM IBA | MRI (65%) | 70% | [137] |
| FT on SEM: MS + 5.0 µM BA | ES (90%) | FT on ½ MS + 0.5 µM IBA + 3.96 µM PG | ER (IVR) | ||||
| Nodal segment | 18-d-old-AS | MS + 4.0 µM BA + 0.5 µM IAA + 20 µM AdS | SM (85%) | 2-step-method: PT on ½ MS (liquid) + 100 µM IBA + 15.84 µM PG | MRI (70%) | 75% | [143] |
| FT on ½ MS (semi-solid) + 0.5 µM IBA | ER (IVR) | ||||||
| Hypocotyl | 12-d-old-AS | MS + 5.0 µM 2,4-D + 1.0 µM BA | Ca-Fm (90%) | ½ MS + 1.0 µM BA | SEG (56%) | 60% | [144] |
| MS + 0.5 µM BA + 0.1 µM NAA + 10 µM ABA | SEs (51%) | ||||||
| Aseptic seed | - | ½ MS basal medium | ISG (96%) | - | - | - | [116] |
| Cotyledonary node | 18-d-old-AS | MS + 1.0 mg·L−1 BAP + 0.5 mg·L−1 NAA | SM (70%) | - | - | - | |
| Aseptic seed | - | ½ MS basal medium | ISG (78.23%) | - | - | - | [19] |
| Immature zygotic embryo | Green fruits | MS + 3.0 mg·L−1 BA + 0.5 mg·L−1 IAA | SM (93.8%) | 2-step-method: PT on ½ MS (liquid) + 3.0 mg·L−1 IBA | MRI (70.8%) | 74% | [56] |
| FT on ½ MS basal medium | ER (IVR) | ||||||
| Immature cotyledon | 9-d-old-AS | 3-step-method: MS + 1.07 µM NAA | Ca-Fm (60.41%) | 2-step-method: | 95% | [145] | |
| FT on MS + 8.9 µM BAP + 1.07 µM NAA | MSI (60.41%) | PT on ½ MS (liquid) + 19.6 µM IBA | MRI (75%) | ||||
| FT on MS + 4.4 µM BAP | ES | FT on ½ MS + 2.85 µM IBA | ER (IVR) | ||||
| Nodal segment | 10-y-old-MT | 2-step-method: MS + 13.95 µM Kn + 568 µM AA + 260 µM CA + 605 µM AmS + 217 µM AdS | MSBB (64.44%) | ½ MS + 4.92 µM IBA | RF (42%) (IVR) | - | [146] |
| FT on MS + 9.3 µM Kn + 0.54 µM NAA + 568 µM AA + 260 µM CA + 605 µM AmS + 217 µM AdS | ES | ||||||
| Nodal segment | 4-w-old-AS | 2-step-method: PT on ½ MS (liquid) + 10.0 µM TDZ | MSBB (96%) | 2-step-method: PT on ½ MS (liquid) + 150 µM IBA | MRI (80%) | 75% | [147] |
| FT on MS (semisolid) + 5.0 µM mT + 1.0 µM NAA | ES (70%) | FT on ½ MS + 1.5 µM IBA | ER (IVR) | ||||
| Cotyledonary node | 20-d-old-AS | MS + 7.5 µM mT + 1.0 µM NAA | SM (85%) | 2-step-method: PT on ½ MS (liquid) + 100 µM IBA | MRI (75%) | 80% | [20] |
| FT on ½ MS + 1.0 µM IBA | ER (IVR) | ||||||
| Immature zygotic embryo | Green fruits | MS + 5.37 µM NAA | SEs (67.3%) | ½ MS + 5.8 µM GA3 | SEG (70%) | 78% | [148] |
| MS + 2.69 µM NAA + 4.4 µM BA + 3% Sucrose | |||||||
| Shoot tip | 7-d-old-AS | MS + 7.0 µM mT + 1.0 µM NAA | SM (80%) | 2-step-method: PT on ½ MS (liquid) + 250 µM IBA | MRI (67.7%) | 96.7% | [149] |
| FT on Soilrite | ER (EVR) | ||||||
| In vitro seedling | Seed | MS + 0.5 µM GA3 + 0.5 µM TDZ | SM (85%) | 2-step-method: PT on ½ MS (liquid) + 100 µM IBA | MRI (80%) | 86.7% | [150] |
| FT on ½ MS + 0.5 µM GA3 | ER (IVR) | ||||||
| Aseptic seeds | - | ½ MS 0.5 µM GA3 | ISG (91.3%) | - | - | - | [119] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, A.; Ahmad, N.; Anis, M.; Faisal, M.; Alatar, A.A.; Abdel-Salam, E.M.; Meena, R.P.; Sivanesan, I. Biotechnological Advances in Pharmacognosy and In Vitro Manipulation of Pterocarpus marsupium Roxb. Plants 2022, 11, 247. https://doi.org/10.3390/plants11030247
Ahmad A, Ahmad N, Anis M, Faisal M, Alatar AA, Abdel-Salam EM, Meena RP, Sivanesan I. Biotechnological Advances in Pharmacognosy and In Vitro Manipulation of Pterocarpus marsupium Roxb. Plants. 2022; 11(3):247. https://doi.org/10.3390/plants11030247
Chicago/Turabian StyleAhmad, Anees, Naseem Ahmad, Mohammad Anis, Mohammad Faisal, Abdulrahman A. Alatar, Eslam M. Abdel-Salam, Ram Pratap Meena, and Iyyakkannu Sivanesan. 2022. "Biotechnological Advances in Pharmacognosy and In Vitro Manipulation of Pterocarpus marsupium Roxb." Plants 11, no. 3: 247. https://doi.org/10.3390/plants11030247