
Dwarfing and the Underlying Morphological Changes of Poa alpigena Plants in Response to Overgrazing Conditions

Abstract
:1. Introduction
2. Results
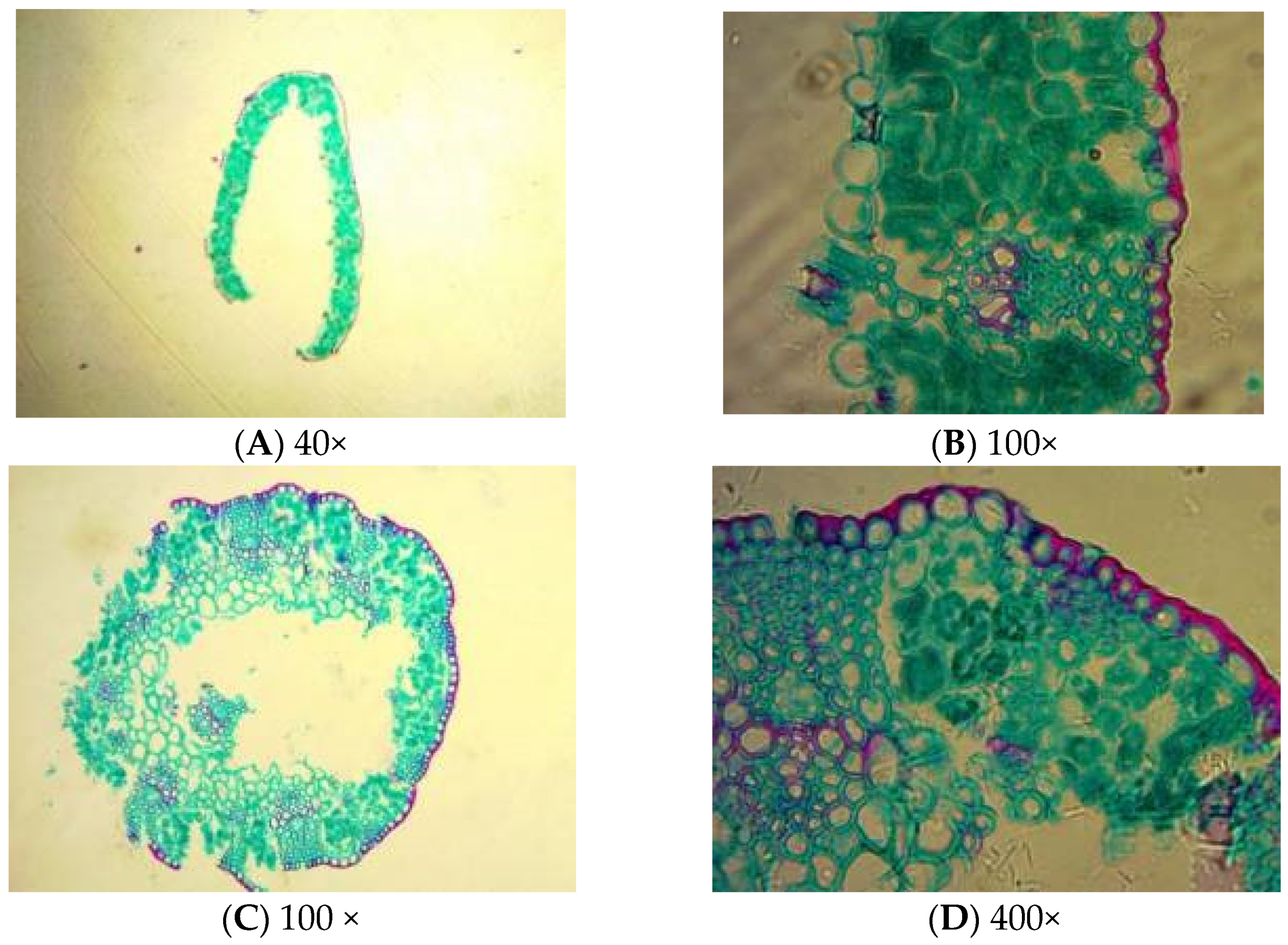
2.1. Poa alpigena Stem and Leaf Structures
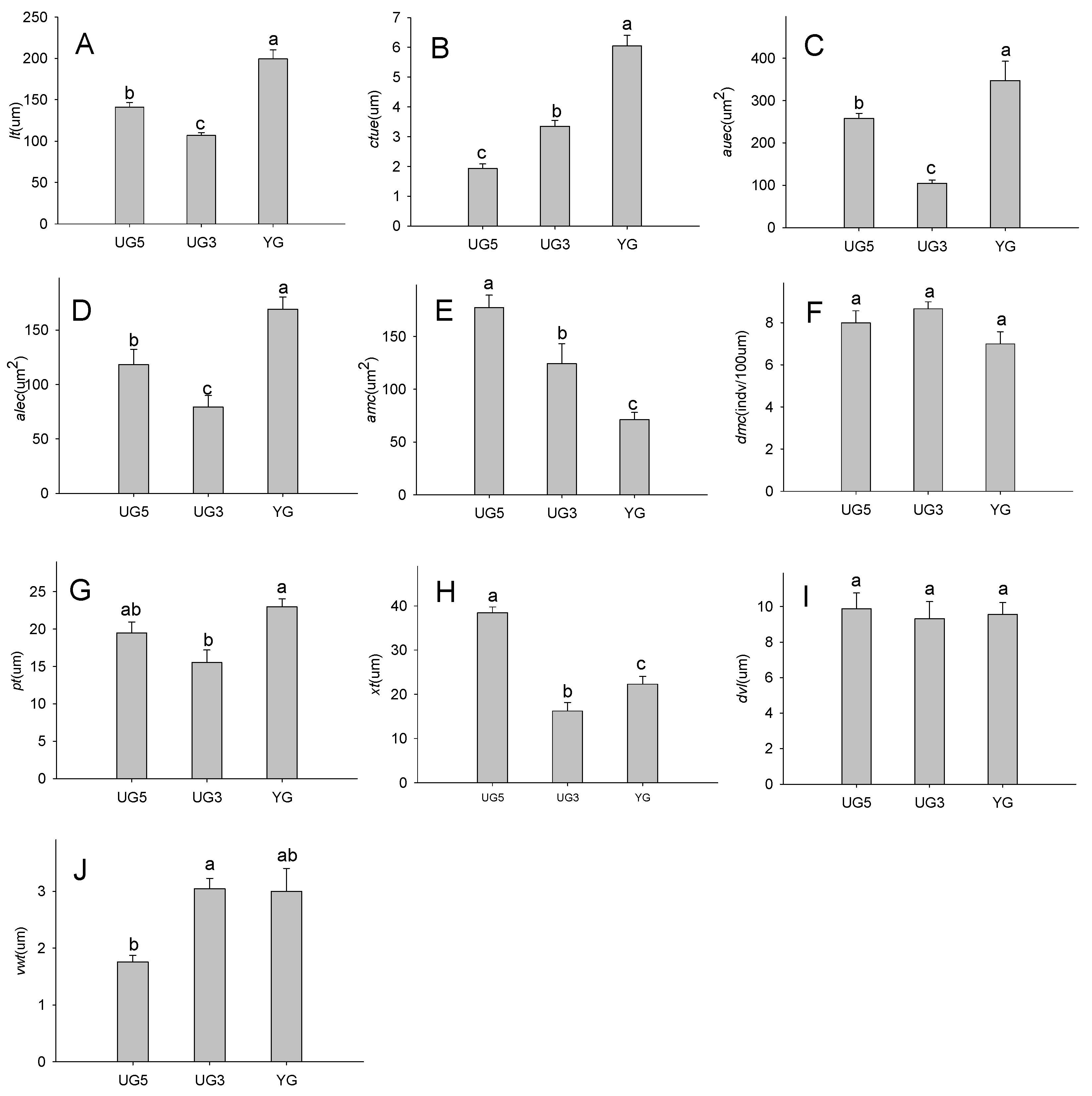
2.2. Effects of Overgrazing and Enclosure on Leaf Structures
2.2.1. Effects of Overgrazing and Enclosure on Leaf Thickness
2.2.2. Effects of Overgrazing and Enclosure on the Epidermal Cuticle
2.2.3. Effects of Overgrazing and Enclosure on Mesophyll Cells
2.2.4. Effects of Overgrazing and Enclosure on Phloem and Xylem Thickness
2.2.5. Effects of Overgrazing and Enclosure on Vessels
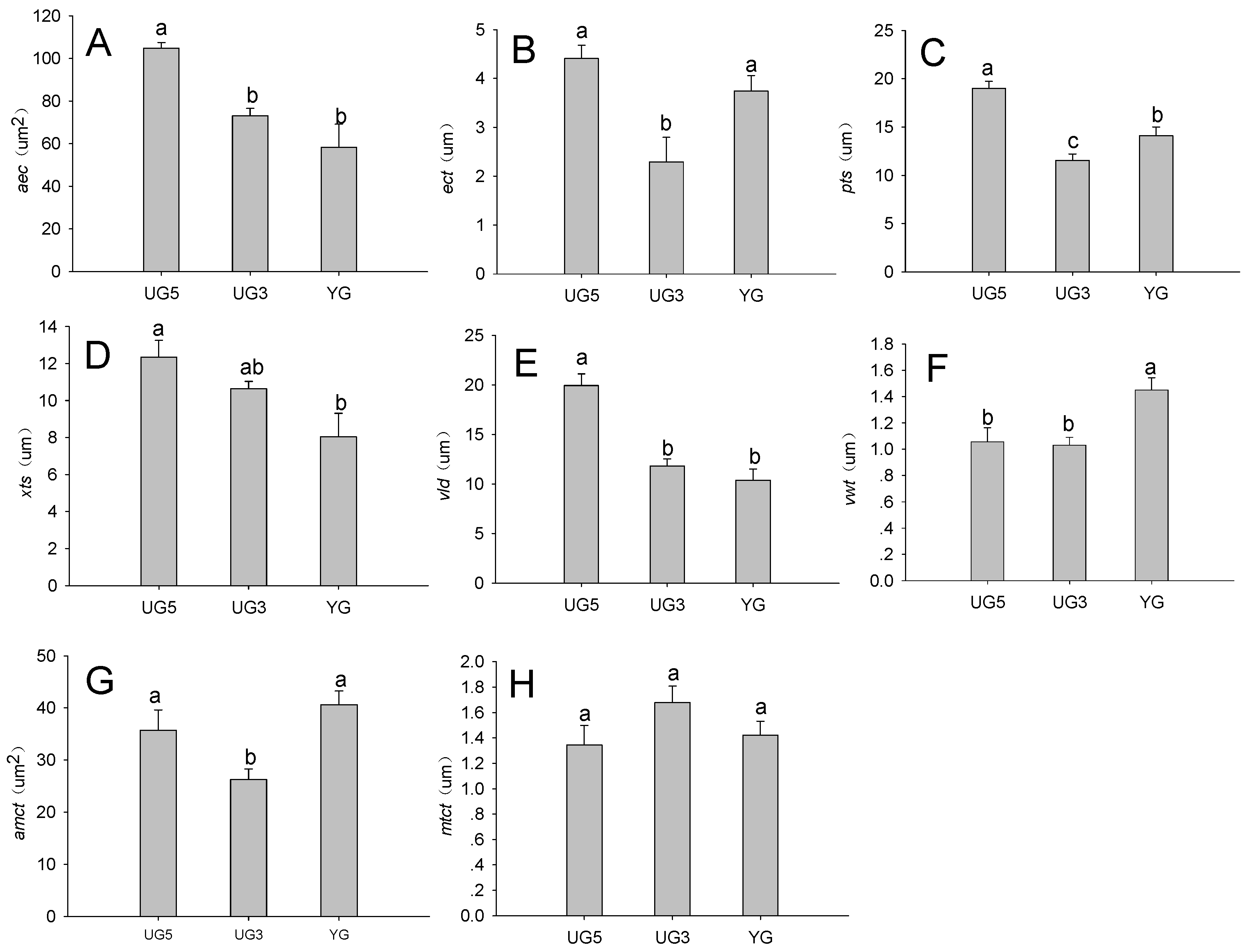
2.3. Effect of Overgrazing and Enclosure on Poa alpigena Stem Structures
2.3.1. Effects of Overgrazing on Stem Epidermis
2.3.2. Effect of Overgrazing on Stem Vascular Bundle Features
2.3.3. Effects of Overgrazing on Stem Vessel Features
2.3.4. Effects of Overgrazing on Stem Mechanical Tissue Features
2.4. Plasticity of Leaf and Stem Structures
3. Discussion
3.1. Effects of Overgrazing and Enclosure on Leaf Structures
3.2. Effects of Overgrazing on Stem Structures
3.3. Adaptability of Stem and Leaf Structures to Overgrazing
4. Conclusions
5. Materials and Methods
5.1. Research Overview
5.2. Plot Conditions and Sampling
5.3. Plots Sampling
5.4. Sample Preparation for Microscopic Analyses
5.5. Microscopy
5.6. Data Analysis
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Saito, K.; Azoma, K.; Sokei, Y. Genotypic adaptation of rice to lowland hydrology in West Africa. Field Crops Res. 2010, 119, 290–298. [Google Scholar] [CrossRef]
- Gell, G.; Petrik, K.; Balázs, E. A unique nucleotide sequence variant in the coat protein region of the genome of a Maize dwarf mosaic virus isolate. Acta Phytopathol. Entomol. Hung. 2011, 46, 11–15. [Google Scholar] [CrossRef]
- Peng, Z.; Li, X.; Yang, Z.; Liao, M. A new reduced height gene found in the tetraploid semi-dwarf wheat landrace Aiganfanmai. Genet. Mol. Res. 2011, 10, 2349–2357. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Liu, X.-D.; Chi, X.-J.; Wu, C.-A.; Li, Y.-Z.; Song, L.-L.; Liu, X.-M.; Wang, Y.-F.; Wang, F.-W.; Zhang, C. Dwarf apple MbDREB1 enhances plant tolerance to low temperature, drought, and salt stress via both ABA-dependent and ABA-independent pathways. Planta 2011, 233, 219–229. [Google Scholar] [CrossRef]
- Leeds, T.; Notter, D.; Leymaster, K.; Mousel, M.; Lewis, G. Evaluation of Columbia, USMARC-Composite, Suffolk, and Texel rams as terminal sires in an extensive rangeland production system: I. Ewe productivity and crossbred lamb survival and preweaning growth. J. Anim. Sci. 2012, 90, 2931–2940. [Google Scholar] [CrossRef]
- Wang, W.; Liang, C.Z.; Liu, Z.L.; Hao, D.Y. Analysis of the plant individual behaviour during the degradation and restoring succession in steppe community. Acta Phytoecol. Sin. 2000, 24, 268–274. [Google Scholar]
- Wang, W.; Liang, C.Z.; Liu, Z.L.; Hao, D.Y. Mechanism of Degradation Succession in Leymus Chinensis + Stipa grandis Steppe Community. Acta Phytoecol. Sin. 2000, 24, 468–472. [Google Scholar]
- He, T.; Wu, X.M.; Zhang, G.N.; Wang, X.R.; Jia, J.F. Characteristics of Starch Grains in Chloroplast of Five Alpine Plants. J. Wuhan Bot. Res. 2005, 23, 545–548. [Google Scholar]
- Yang, Y.; Sun, H. Advances in the Functional Ecology of Alpine and Arctic Plants. Acta Bot. Yunnanica 2006, 28, 43–53. [Google Scholar]
- He, T.; Wu, X.M.; Zhang, W.J.; Liu, J.X. Study of correlative location between chloroplast and mitochondria in alpine plants. Acta Bot. Boreali-Occident. Sin. 2004, 24, 728–731. [Google Scholar]
- Wang, X.L.; Wang, J. Morphology and Structure of Plants; Lanzhou University Press: Lanzhou, China, 1989; pp. 105–148. [Google Scholar]
- Jiang, G.C.; Jia, X.H. Comparson of the Microstructures of Vegetative Organs of Three Species of Kobresia Growing in Different Altitude. J. Henan Univ. 1999, 1, 63–68. [Google Scholar]
- Wu, K.; Zhou, X.Y. Effects of Environmental Stresses on Plant Ultrastructure. ShanDong For. Sci. Technol. 2007, 3, 79–83. [Google Scholar]
- Zheng, X.P.; Zhang, C.L.; Chen, G.C. Investigaion on the Daptation of Photosynthetic Carbon Metabolism Pathway to Environment of Phragmites Communis in Hexi Corridor of Gansu Province. Acta Phytoecol. Geobot. Sin. 1993, 17, 1–8. [Google Scholar]
- Li, Z.L.; Li, R.A. Anatomical Observation of Assimilating Branches of Nine Xerophytes in Gansu. Acta Bot. Sin. 1981, 23, 181–185. [Google Scholar]
- He, J.S.; Chen, W.L. Morphological and Anatomical Features of Quercus Section Suber and Its Adaptation to the Ecological Environment. Acta Phytoecol. Sin. 1994, 3, 219–227. [Google Scholar]
- Boardman, N.K. Comparative photosynthesis of sun and shade plants. Ann. Rev. Plant Physiol. 1977, 28, 355–377. [Google Scholar] [CrossRef]
- Gao, Q. Adaptability Research of Morphological Structure to the Dese. Master’s Thesis, Inner Mongolia University, Hohhot, China, May 2008. [Google Scholar]
- Wylic, R.B. Relations between tissue organization and vascularization in leaves of certain tropical and subtropical dicotyledons. Am. J. Bot. 1946, 33, 721–725. [Google Scholar]
- Chabot, B.F.; Jurid, T.W. Influence of instantaneous and integrated light flux density on leaf anatomy and photosynthesis. Am. J. Bot. 1979, 66, 721–725. [Google Scholar] [CrossRef]
- Lin, F.Z.; Lin, G.Z. The photosynthesis procedure with Dinghushan canopy leaf structure properties analysis. J. Collect. China South-East Inst. Bot. Res. 1989, 5, 101–108. [Google Scholar]
- Zhong, Z.C. The Evergreen Broadleaf Forests Ecological Procedure Research; South-West Normal University Press: Chongqing, China, 1992. [Google Scholar]
- Shi, H.X.; Hou, X.Y.; Shi, S.L.; Wu, X.H.; Yang, T.T.; Li, P. Poa alpigena response traits affected by grazing and enclosuers in an alpine meadow on the Qinghai-Tiber Plateau. Acta Ecol. Sin. 2016, 36, 3601–3608. [Google Scholar]
- Wang, W.C.; Chen, J.R. The Vegetation Collection of China; Science Technology Press: Beijing, China, 1990. [Google Scholar]
- Wang, X.L.; Wang, J. The Vegetation Appearance Patterns and Correlated Environment Structure; Lanzhou University Press: Lanzhou, China, 1989. [Google Scholar]
- Li, H.; Li, B.; Ding, W.C.; Shi, S.L.; Li, T.Y. Effect of polyethylene glycol(PEG) stress on the leaf blade microstructure and ultrastructure of two kinds of Agropyron cristatum. Heilongjiang Anim. Sci. Vet. Med. 2015, 6, 28–31. [Google Scholar]
- Zhang, H.X.; Liu, G.H.; Cui, X.P. Affection of aridity to anatomical structure of leave of Ulmus pumila L. var. sabulosa. Bull. Bot. Res. 2005, 25, 39–44. [Google Scholar]
- Zhao, X.Y.; Wang, S.P. Responses of the anatomical characteristics of plant leaf to long-term grazing under different stocking rates in Inner Mongolia steppe. Acta Ecol. Sin. 2009, 29, 2906–2918. [Google Scholar]
- Fan, C.M.; Liao, C.Y.; Liao, C.Y.; Li, P.Y.; Sun, C.Z.; Xu, X.M. A study of the effects of different grazing intensities on soil physical properties of grassland and forest floor—For example Hilly and Gully regions on the loss Platesu. Sci. Agric. Sin. 2006, 39, 1501–1506. [Google Scholar]
- Yin, G.M.; Zhao, J.H.; Xue, Y.L. Effects of grazing pattern on the anatomical structure characteristics of Cleistengenes songorica leaves. Anim. Husb. Feed Sci. 2014, 35, 28–30. [Google Scholar]
- Chen, Y.; Chen, Y.J.; Zhou, Y.; Yan, Q.W.; Li, X. Stem structure associated with drought resistance in different clover cultivars. Acta Agrestia Sin. 2012, 4, 689–691. [Google Scholar]
- Yu, X.Z.; He, X.; Zhang, T. Response of leaf structures of 8 plants to grazing prohibition in degraded grassland of Inner Mongolia. Acta Ecol. Sin. 2007, 27, 1638–1645. [Google Scholar]
- Zhen, W.J.; Xun, L.Y.; Wang, X.L. The salty pressure response for the vegetation structure analysis. Acta Pratacult. Sin. 1993, 2, 78–80. [Google Scholar]
- Li, J.C.; Zhao, Y.E.; Kang, B.W.; Yao, Z.C. Anatomical observation of assimilatine branches of some species in Yanchi county, NingXia. Acta Bot. Boreali-Occident. Sinica. 1989, 9, 191–196. [Google Scholar]
- Wang, X.L.; Ma, J.A. Study on leaf—Structure and the diversity of xerophytes ecology adaptation. Acta Ecol. Sin. 1999, 19, 787–792. [Google Scholar]
- Shi, G.R.; Tang, Y.; Zhang, Z. Leaf anatomy of dominant plant species in the successional communities of XiangShan Moutnain, HuaiBei, China. Chin. J. Plant Ecol. 2006, 30, 314–322. [Google Scholar]
- Li, X.L.; Hou, X.Y.; Wu, X.H.; Ji, L.; Chen, H.J.; Liu, Z.Y.; Ding, Y. Plastic responses of stem and leaf functional traits in Leymus chinensis to long-term grazing in a meadow steppe. Chin. J. Plant Ecol. 2014, 5, 440–451. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| lt | uect | auec | alec | amc | dmc | pt | xt | dvl | vwt |
|---|---|---|---|---|---|---|---|---|---|
| 0.46 | 0.68 | 0.53 | 0.70 | 0.60 | 0.19 | 0.33 | 0.58 | 0.06 | 0.42 |
| vld | vwt | aec | ect | xts | pts | mtct | amtc |
|---|---|---|---|---|---|---|---|
| 0.41 | 0.29 | 0.44 | 0.48 | 0.35 | 0.39 | 0.20 | 0.35 |
| Plot | Latitude | Grazing Intensity | Land-Use-Type | Dominant Species | |||
|---|---|---|---|---|---|---|---|
| Longitude | Latitude | Altitude (m) | Area (ha) | ||||
| YG | 33°0′27.72″ | 97°18′13.32″ | 4279.9 | Heavy grazing | 66.67 | Annual grazing from January to December | Kobresia pygmea, Kobresia kansuensis |
| UG3 | 33°21′38.25″ | 97°18′6.65″ | 4277.4 | No grazing | 6.67 | Fenced and no grazing in 2011 | Elymus natans, Poa crymophila, Kobresia pygmea, Kobresia kansuensis |
| UG5 | 33°21′30.24″ | 97°18′10.08″ | 4198.1 | No grazing | 13.33 | Fenced and no grazing in 2009 | Elymus natans, Poa crymophila, Kobresia pygmea |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, H.; Wu, X.; Wang, H.; Ha, S.; Yang, T.; Liu, W. Dwarfing and the Underlying Morphological Changes of Poa alpigena Plants in Response to Overgrazing Conditions. Plants 2022, 11, 336. https://doi.org/10.3390/plants11030336
Shi H, Wu X, Wang H, Ha S, Yang T, Liu W. Dwarfing and the Underlying Morphological Changes of Poa alpigena Plants in Response to Overgrazing Conditions. Plants. 2022; 11(3):336. https://doi.org/10.3390/plants11030336
Chicago/Turabian StyleShi, Hongxiao, Xinhong Wu, Hai Wang, Sibagen Ha, Tingting Yang, and Wenhui Liu. 2022. "Dwarfing and the Underlying Morphological Changes of Poa alpigena Plants in Response to Overgrazing Conditions" Plants 11, no. 3: 336. https://doi.org/10.3390/plants11030336
APA StyleShi, H., Wu, X., Wang, H., Ha, S., Yang, T., & Liu, W. (2022). Dwarfing and the Underlying Morphological Changes of Poa alpigena Plants in Response to Overgrazing Conditions. Plants, 11(3), 336. https://doi.org/10.3390/plants11030336