
Sex Differences in Desiccation Tolerance Varies by Colony in the Mesic Liverwort Plagiochila porelloides



Abstract
:1. Introduction
2. Material & Methods
2.1. Study Organism, Sampling Conditions and Field Characteristics
2.2. Desiccation Tolerance Assay and Recovery of Photosystem II
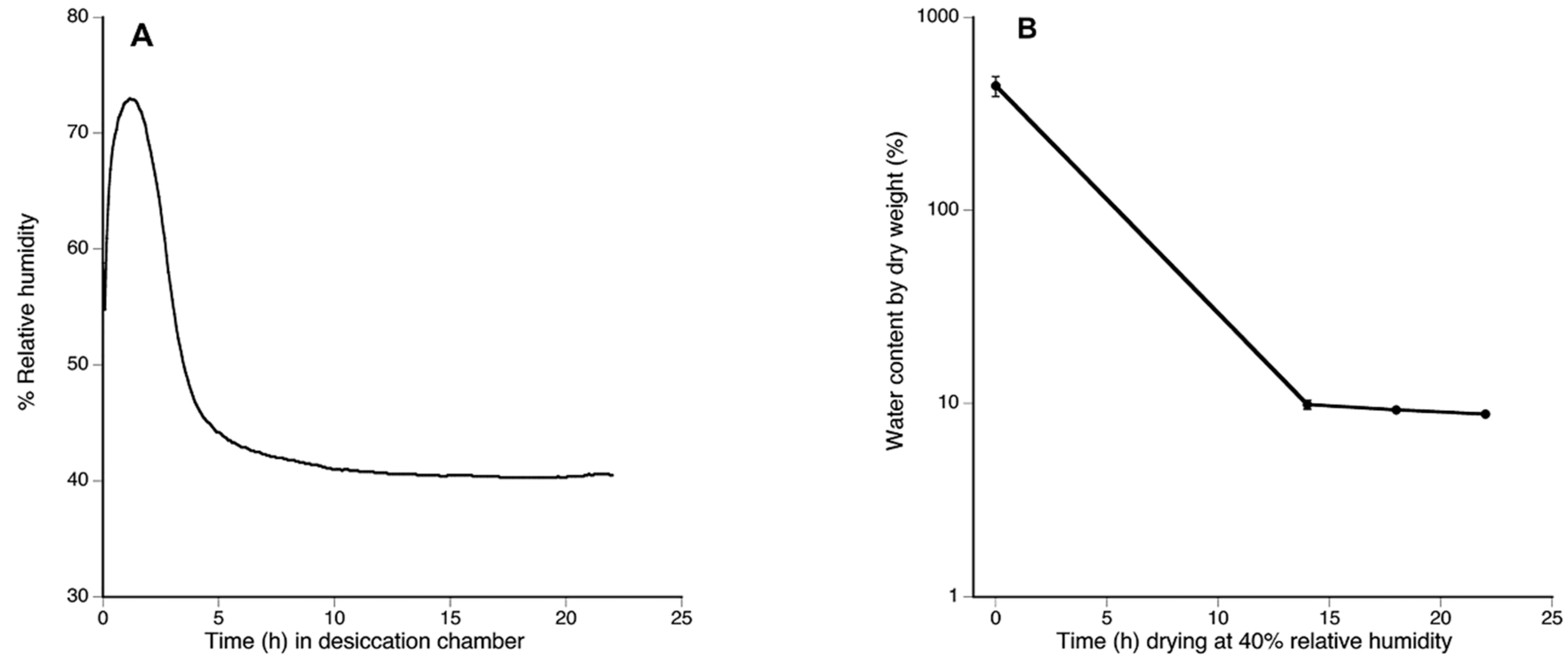
2.3. Evaluation of Water Content by Dry Weight
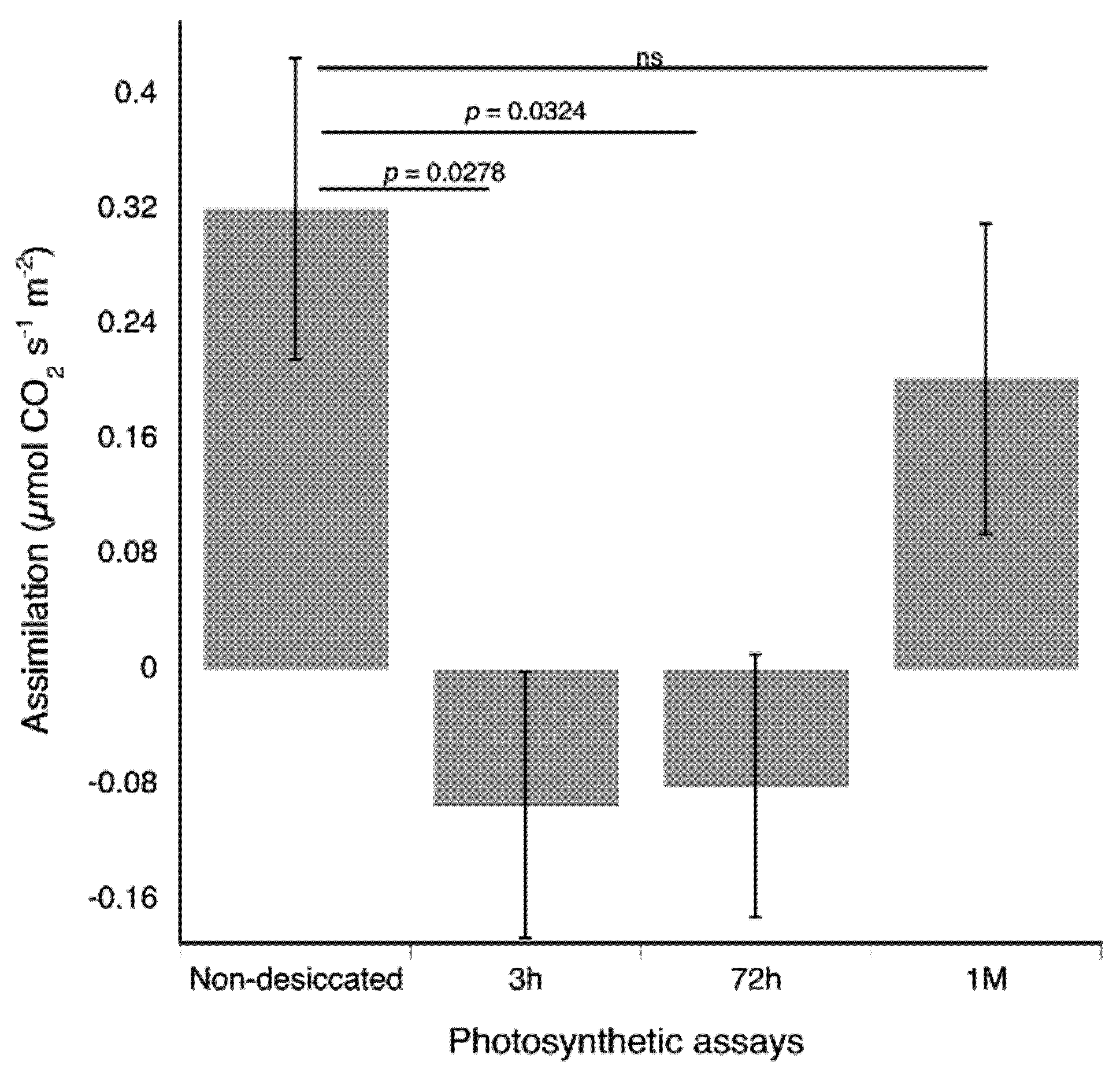
2.4. Gas Exchange Responses to Desiccation
2.5. Statistical Analyses
2.5.1. Desiccation Tolerance Assay
2.5.2. Evaluation of Water Content by Dry Weight
2.5.3. Gas Exchange in the Light in Response to Desiccation
2.5.4. Gas Exchange in the Dark in Response to Desiccation
3. Results
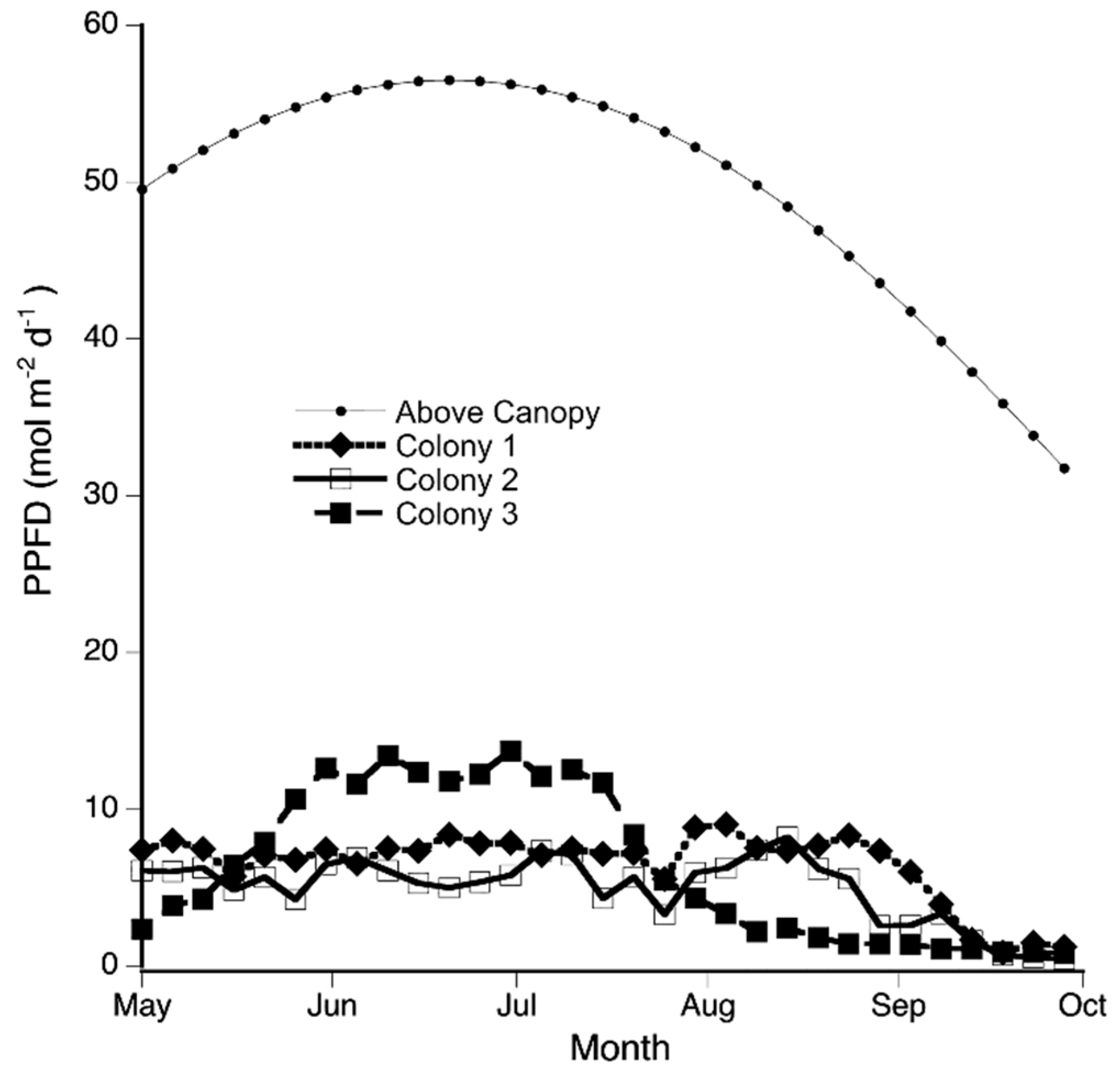
3.1. Field Environmental Conditions
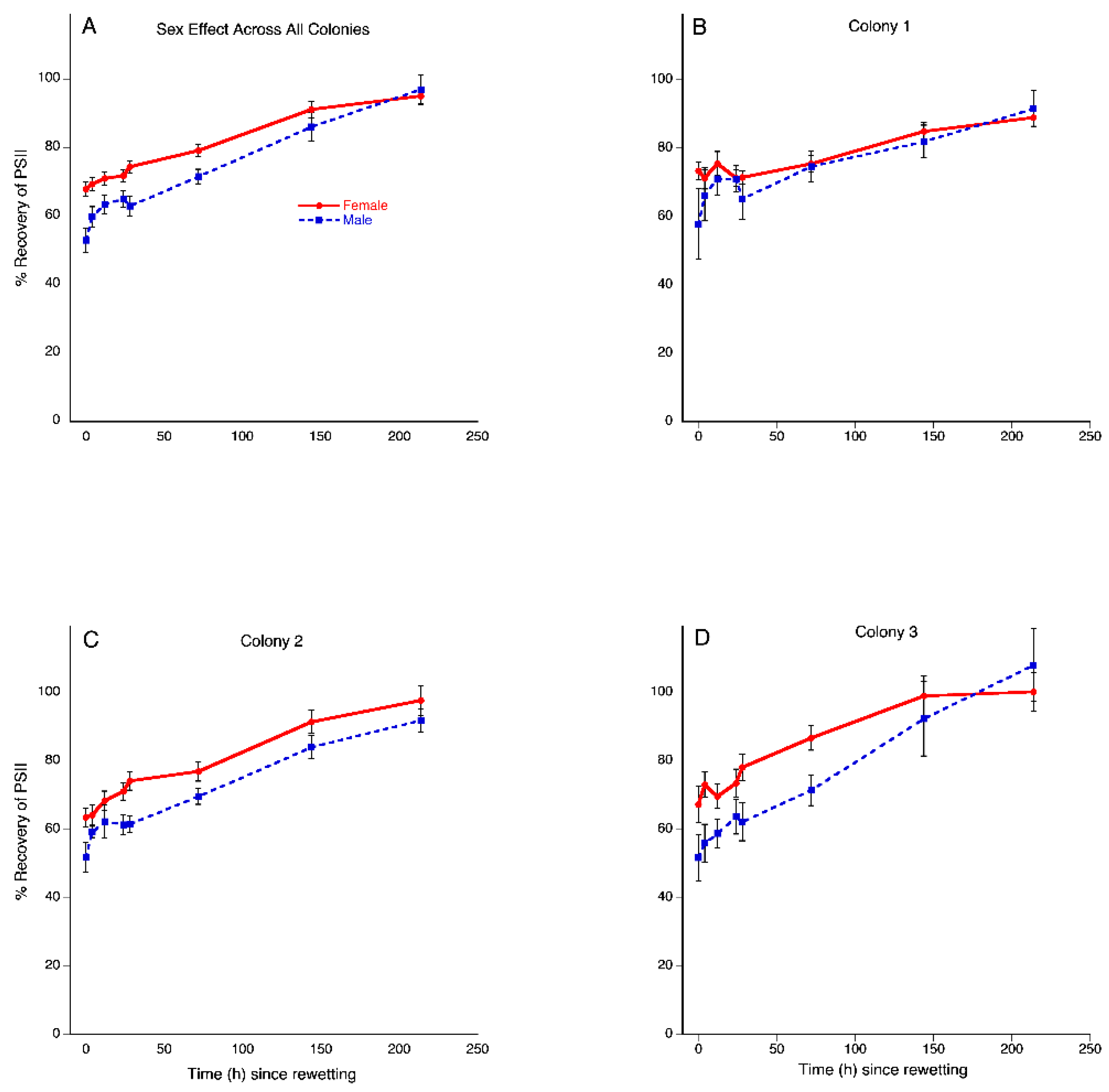
3.2. Desiccation Tolerance Recovery
3.2.1. Main Effects
3.2.2. Time × Colony Interaction Effect
3.2.3. Time × Sex Interaction Effect
3.2.4. Time × Colony × Sex Interaction Effect
3.3. Water Content by Dry Weight
3.4. Gas Exchange Responses to Desiccation
4. Discussion
4.1. Colony Specific Responses and Environmental Associations
4.2. Sex Differences on Water Stress Tolerance
4.3. Physiological Responses to Recovery on Rehydration Processes
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data availability statement
Acknowledgments
Conflicts of Interest
References
- Osakabe, Y.; Kawaoka, A.; Nishikubo, N.; Osakabe, K. Responses to environmental stresses in woody plants: Key to survive and longevity. J. Plant Res. 2011, 125, 1–10. [Google Scholar] [CrossRef]
- Smirnoff, N. Plant Stress Physiology. In Encyclopedia of Life Science, 1st ed.; John Wiley and Sons: Chichester, UK, 2014; Volume 26, pp. 1–11. [Google Scholar]
- Alpert, P. The limits and frontiers of desiccation-tolerant life. Integr. Comp. Biol. 2005, 45, 685–695. [Google Scholar] [CrossRef] [Green Version]
- Alpert, P.; Oliver, M.J. Drying without dying. In Desiccation and Survival in Plants: Drying without Dying; Black, M., Pritchard, H.W., Eds.; CABI Publishing: Wallingford, UK, 2002; pp. 3–43. [Google Scholar]
- Farrant, J.M.; Lehner, A.; Cooper, K.; Wiswedel, S. Desiccation tolerance in the vegetative tissues of the fern Mohria caffrorum is seasonally regulated. Plant J. 2009, 57, 65–79. [Google Scholar] [CrossRef]
- Zhang, Q.; Bartels, D. Molecular responses to dehydration and desiccation in desiccation-tolerant angiosperm plants. J. Exp. Bot. 2018, 69, 3211–3222. [Google Scholar] [CrossRef] [PubMed]
- Bewley, J.D. Physiological aspects of desiccation tolerance. Ann. Rev. Plant Physiol. 1979, 30, 195–238. [Google Scholar] [CrossRef]
- Proctor, M.C.F.; Pence, V. Vegetative Tissues: Bryophytes, vascular resurrection plants and vegetative propagules. In Desiccation and Survival in Plants: Drying without Dying; Black, M., Pritchard, H.W., Eds.; CABI Publishing: Wallingford, UK, 2002; pp. 207–238. [Google Scholar]
- Stark, L.R. Ecology of desiccation tolerance in bryophytes: A conceptual framework and methodology. Bryologist 2017, 120, 130–165. [Google Scholar] [CrossRef]
- Oliver, M.J. Biochemical and molecular mechanisms of desiccation tolerance in bryophytes. In Bryophyte Biology; Shaw, J.A., Goffinet, B., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 269–298. [Google Scholar]
- Vanderpoorten, A.; Goffinet, B. Introduction to Bryophyte Biology; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Lüttge, U.; Beck, E.; Bartels, D. Plant Desiccation Tolerance; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Vitt, D.H.; Crandall-Stotler, B.J.; Wood, A.J. Survival in a dry world through avoidance and tolerance. In Plant Ecology and Evolution in Harsh Environments; Rajakaruna, N., Boyd, N.R., Harris, T., Eds.; Nova Science Publishers: New York, NY, USA, 2014; pp. 267–295. [Google Scholar]
- Oliver, M.J.; Farrant, J.M.; Hilhorst, H.W.M.; Mundree, S.; Williams, B.; Bewley, J.D. Desiccation Tolerance: Avoiding Cellular Damage During Drying and Rehydration. Annu. Rev. Plant Biol. 2020, 71, 435–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marks, R.A.; Farrant, J.M.; Mcletchie, D.N.; VanBuren, R. Unexplored dimensions of variability in vegetative desiccation tolerance. Am. J. Bot. 2021, 108, 346–358. [Google Scholar] [CrossRef]
- Freeman, D.C.; McArthur, E.D. A comparison of twig water stress between males and females of six species of desert shrubs. For. Sci. 1982, 28, 304–308. [Google Scholar]
- Sinclair, J.; Emlen, J.; Freeman, D.C. Biased Sex Ratios in Plants: Theory and Trends. Bot. Rev. 2012, 78, 63–86. [Google Scholar] [CrossRef]
- Juvany, M.; Munné-Bosch, S. Sex-related differences in stress tolerance in dioecious plants: A critical appraisal in a physiological context. J. Exp. Bot. 2015, 66, 6083–6092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delph, L.F. Sexual Dimorphism in Life History. In Gender and Sexual Dimorphism in Flowering Plants; Geber, M.A., Dawson, T.E., Delph, L.F., Eds.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 149–173. [Google Scholar]
- Newton, M.E. Sex-ratio differences in Mnium hornum Hedw. and M. undulatum Sw. in relation to spore germination and vegetative regeneration. Ann. Bot. 1972, 36, 163–178. [Google Scholar] [CrossRef]
- Stieha, C.R.; Middleton, A.R.; Stieha, J.K.; Trott, S.H.; Mcletchie, D.N. The dispersal process of asexual propagules and the contribution to population persistence in Marchantia inflexa (Marchantiaceae). Am. J. Bot. 2014, 101, 348–356. [Google Scholar] [CrossRef]
- Marks, R.A.; Burton, J.F.; McLetchie, D.N. Sex differences and plasticity in dehydration tolerance: Insight from a tropical liverwort. Ann. Bot. 2016, 118, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Slate, M.; Rosenstiel, T.N.; Eppley, S.M. Sex-specif.f.fic morphological and physiological differences in the moss Ceratodon purpureus (Dicranales). Ann. Bot. 2017, 120, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Marks, R.A.; Pike, B.; McLetchie, D.N. Water stress tolerance tracks environmental exposure and exhibits a fluctuating sexual dimorphism in a tropical liverwort. Oecologia 2019, 191, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Alpert, P. The discovery, scope and puzzle of desiccation tolerance in plants. Plant Ecol. 2000, 151, 5–17. [Google Scholar] [CrossRef]
- Giordani, P.; Incerti, G.; Rizzi, G.; Rellini, I.; Nimis, P.L.; Modenesi, P. Functional traits of cryptogams in Mediterranean ecosystems are driven bywater, light and substrate interactions. J. Veg. Sci. 2013, 25, 778–792. [Google Scholar] [CrossRef]
- Ekwealor, J.T.B.; Clark, T.A.; Dautermann, O.; Russell, A.; Ebrahimi, S.; Stark, L.R.; Niyogi, K.K.; Mishler, B.D. Natural UV exposure alters photosynthetic biology and improves recovery from desiccation in a desert moss. J. Exp. Bot. 2021, 11, 4161–4179. [Google Scholar] [CrossRef]
- Söderström, L.; Hagborg, A.; Von Konrat, M. Notes on early land plants today. 69. Circumscription of Plagiochilaceae (Marchantiophyta) with a preliminary infrageneric subdivision of Plagiochila. Phytotaxa 2015, 208, 75–91. [Google Scholar] [CrossRef] [Green Version]
- Söderström, L.; Hagborg, A.; von Konrat, M.; Bartholomew-Began, S.; Bell, D.; Briscoe, L.; Brown, E.; Cargill, D.C.; Costa, D.P.; Crandall-Stotler, B.J.; et al. World checklist of hornworts and liverworts. PhytoKeys 2016, 59, 1–828. [Google Scholar] [CrossRef] [Green Version]
- Clausen, E. Hepatics and humidity, a study on the occurrence of hepatics in a Danish tract and the influence of relative humidity on their distribution. Dansk Bot. Ark. 1952, 15, 1–80. [Google Scholar]
- Dilks, T.J.K.; Proctor, M.C.F. The pattern of recovery of bryophytes after desiccation. J. Bryol. 1974, 8, 97–115. [Google Scholar] [CrossRef]
- Dilks, T.J.K.; Proctor, M.C.F. Seasonal variation in desiccation tolerance in some British bryophytes. J. Bryol. 1976, 9, 239–247. [Google Scholar] [CrossRef]
- Gupta, R.K. The Physiology of the Desiccation Resistance in Bryophytes: Nature of Organic Compounds Leaked from Desiccated Liverwort, Plagiochila asplenioides. Biochem. Physiol. Pflanzen 1976, 170, 389–395. [Google Scholar] [CrossRef]
- Wood, A.J. The nature and distribution of vegetative desiccation-tolerance in hornworts, liverworts and mosses. Bryologist 2007, 110, 163–177. [Google Scholar] [CrossRef]
- Söderström, L.; Rycroft, D.S.; Cole, W.J.; Wei, S. Plagiochila porelloides (Plagiochilaceae, Hepaticae) form Changbai Mountain, new to China, with chemical characterization and chromosomes measurements. Bryobrothera 1999, 5, 195–201. [Google Scholar]
- Cronberg, N. Absence of genetic variation in populations of the liverwort Plagiochila porelloides from northern Greece and southern Scandinavia. Lindbergia 2000, 25, 20–24. [Google Scholar]
- Sim-Sim, M.; Carvalho, S.; Fontinha, S.; Lobo, C.; Garcia, C. Plagiochila porelloides (Torr. ex Nees) Lindenb. in mainland Portugal and Madera archipelago. New records and the threatened status. Port. Acta Biol. 2003, 21, 231–238. [Google Scholar]
- Stotler, R.E.; Cradall-Stotler, B.J. A Checklist of the Liverworts and Hornworts of North America. Bryologist 1997, 8, 405–428. [Google Scholar] [CrossRef]
- Stotler, R.E.; Cradall-Stotler, B.J. A Synopsis of the liverwort flora of north America north and Mexico. Ann. Mo. Bot. Gard. 2017, 102, 574–709. [Google Scholar] [CrossRef]
- Schuster, R.M. The Hepaticae and Anthocerotae of North America East of the Hundredth Meridian. Vol. IV; Columbia University Press: New York, NY, USA, 1980. [Google Scholar]
- Mägdefrau, K. Life-forms of Bryophytes. In Bryophyte Ecology; Smith, A.J.E., Ed.; Springer: Dordrecht, The Netherlands, 1982; pp. 45–58. [Google Scholar]
- Gradstein, S.R.; Churchill, S.P.; Salazar-Allen, N. Guide to the Bryophytes of Tropical America; The New York Botanical Garden Press: New York, NY, USA, 2001. [Google Scholar]
- Greenspan, L. Humidity Fixed Points of Binary Saturated Aqueous Solutions. J. Res. Natl. Bur. Stand. 1977, 81, 89–96. [Google Scholar] [CrossRef]
- Proctor, M.C.F.; Oliver, M.J.; Wood, A.J.; Stark, L.R.; Cleavitt, N.L.; Mishler, B.D. Desiccation-tolerance in bryophytes: A review. Bryologist 2007, 110, 595–621. [Google Scholar] [CrossRef]
- Schonbeck, M.W.; Bewley, J.D. Responses of the moss Tortula ruralis to desiccation treatments. II. Variations in desiccation tolerance. Botany 1981, 59, 2707–2712. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, J. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd ed.; W.H. Freeman and Company: New York, NY, USA, 1995. [Google Scholar]
- Stark, L.R.; Greenwood, J.L.; Brinda, J.C.; Oliver, M.M.J. Physiological history may mask the inherent inducible desiccation tolerance strategy of the desert moss Crossidium crassinerve. Plant Biol. 2014, 16, 935–946. [Google Scholar] [CrossRef] [PubMed]
- Brinda, J.C.; Stark, L.R.; Clarck, T.A.; Greenwood, J.L. Embryos of a moss can be hardened to desiccation tolerance: Effects of rate of drying on the timeline of recovery and dehardening in Aloina ambigua (Pottiaceae). Ann. Bot. 2016, 117, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Stark, L.R.; Oliver, M.J.; Mishler, B.D.; McLetchie, D.N. Generational differences in response to desiccation stress in the desert moss Tortula inermis. Ann. Bot. 2007, 99, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Proctor, M.C.F. Physiological Ecology. In Bryophyte Biology; Shaw, J.A., Goffinet, B., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 237–268. [Google Scholar]
- Haig, D. Living together and living apart: The sexual lives of bryophytes. Philos. Trans. R. Soc. B 2016, 371, 20150535. [Google Scholar] [CrossRef] [Green Version]
- Stark, L.R.; Mishler, B.D.; McLetchie, D.N. The cost of realized sexual reproduction: Assessing patterns of reproductive allocation and sporophyte abortion in a desert moss. Am. J. Bot. 2000, 87, 1599–1608. [Google Scholar] [CrossRef]
- Stark, L.R.; Nichols, L.; McLetchie, D.N.; Bonine, M.L. Do the sexes of the desert moss Syntrichia caninervis differ in desiccation tolerance: A leaf regeneration assay. Int. J. Plant Sci. 2005, 166, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.; Kollar, L.; McLetchie, D.N. Does selection for gamete dispersal and capture lead to a sex difference in clump water-holding capacity? Am. J. Bot. 2016, 103, 1449–1457. [Google Scholar] [CrossRef] [Green Version]
- Stark, L.R.; Greenwood, J.L.; Brinda, J.C.; Oliver, M.J. The desert moss Pterygoneurum lamellatum (Pottiaceae) exhibits an inducible ecological strategy of desiccation tolerance: Effects of rate of drying on shoot damage and regeneration. Am. J. Bot. 2013, 100, 1522–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leigh, A.; Nicotra, A.B. Sexual dimorphism in reproductive allocation and water use efficiency in Maireana pyramidata (Chenopodiaceae), a dioecious, semi-arid shrub. Aust. J. Bot. 2003, 51, 509–514. [Google Scholar] [CrossRef]
- Li, C.; Ren, J.; Luo, J.; Lu, R. Sex-specific physiological and growth responses to water stress in Hippophae rhamnoides L. population. Acta Physiol. Plant 2004, 26, 123–129. [Google Scholar] [CrossRef]
- Álvarez-Cansino, L.; Zunzunegui, M.; Díaz-Barradas, M.C.; Esquivias, M.P. Physiological performance and xylem water isotopic composition underlie gender-specific responses in the dioecious shrub Corema album. Physiol. Plant 2010, 140, 32–45. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, S.; Zhao, H.; Korpelainen, H.; Li, C. Sex-related adaptive responses to interaction of drought and salinity in Populus yunnanensis. Plant Cell Environ. 2010, 33, 1767–1778. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, F.; Peng, S.; Ma, W.; Korpelainen, H.; Li, C. Comparative physiological, ultrastructural and proteomic analyses reveal sexual differences in the responses of Populus cathayana under drought stress. Proteomics 2010, 10, 2661–2677. [Google Scholar] [CrossRef]
- Mishler, B.D.; Oliver, M.J. Putting Physcomitrella Patens on the tree of life: The evolution and ecology of mosses. In The Moss Physcomitrella Patens; Knight, C.D., Perroud, P.F., Cove, D.J., Eds.; Blackwell Publishing: Singapore, 2009; pp. 1–15. [Google Scholar]
- Proctor, M.C.F.; Smirnoff, N. Rapid recovery of photosystems on rewetting desiccation-tolerant mosses: Chlorophyll fluorescence and inhibitor experiments. J. Exp. Bot. 2000, 51, 1695–1704. [Google Scholar] [CrossRef]
- Proctor, M.C.F. Patterns of desiccation tolerance and recovery in bryophytes. J. Plant Growth Regul. 2001, 35, 147–156. [Google Scholar] [CrossRef]
- Oliver, M.J. Influence of protoplasmic water loss on the control of protein synthesis in the desiccation-tolerant moss Tortula ruralis: Ramifications for a repair-based mechanism of desiccation tolerance. Plant Phys. 1991, 97, 1501–1511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinshiri, H.N.; Proctor, M.C.F. The effect of desiccation on subsequent assimilation and respiration of the bryophytes Anomodon viticulosus and Porella platyphylla. New Phytol. 1971, 70, 527–538. [Google Scholar] [CrossRef]
- Coe, K.K.; Belnap, J.; Sparks, J.P. Precipitation-driven carbon balance controls survivorship of desert biocrust mosses. Ecology 2012, 93, 1626–1636. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Colony | Canopy Openness (%) | PPFD ± se (mol m−2 d−1) | Red/Far Red Ratio | Sex Difference in Desiccation Tolerance |
|---|---|---|---|---|
| C1 | 6.54 | 6.50 ± 0.40 | 0.19 | ns |
| C2 | 6.49 | 4.95 ± 0.38 | 0.24 | ns |
| C3 | 11.30 | 6.35± 0.87 | 0.46 | F > M (p < 0.001) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva-e-Costa, J.d.C.; Luizi-Ponzo, A.P.; McLetchie, D.N. Sex Differences in Desiccation Tolerance Varies by Colony in the Mesic Liverwort Plagiochila porelloides. Plants 2022, 11, 478. https://doi.org/10.3390/plants11040478
Silva-e-Costa JdC, Luizi-Ponzo AP, McLetchie DN. Sex Differences in Desiccation Tolerance Varies by Colony in the Mesic Liverwort Plagiochila porelloides. Plants. 2022; 11(4):478. https://doi.org/10.3390/plants11040478
Chicago/Turabian StyleSilva-e-Costa, Juliana da C., Andrea P. Luizi-Ponzo, and David Nicholas McLetchie. 2022. "Sex Differences in Desiccation Tolerance Varies by Colony in the Mesic Liverwort Plagiochila porelloides" Plants 11, no. 4: 478. https://doi.org/10.3390/plants11040478
APA StyleSilva-e-Costa, J. d. C., Luizi-Ponzo, A. P., & McLetchie, D. N. (2022). Sex Differences in Desiccation Tolerance Varies by Colony in the Mesic Liverwort Plagiochila porelloides. Plants, 11(4), 478. https://doi.org/10.3390/plants11040478