
Turf Quality and Physiological Responses to Summer Stress in Four Creeping Bentgrass Cultivars in a Subtropical Zone



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Measurements of Turf Quality and Photosynthetic Parameters
2.3. Measurements of Water Status and Osmolyte Contents
2.4. Measurements of Oxidative Damage and Membrane Stability
2.5. Experimental Design and Statistical Analysis
3. Results
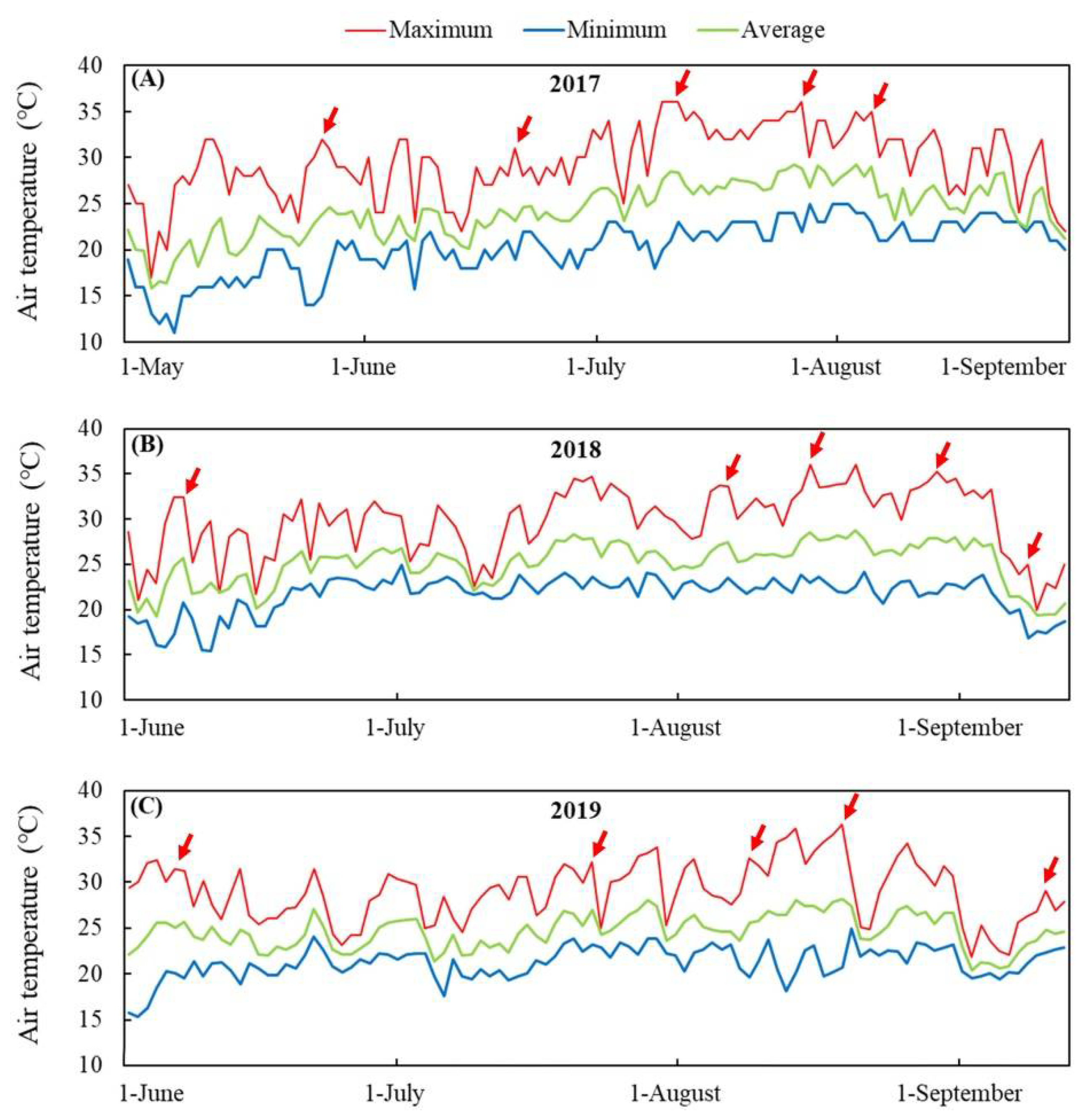
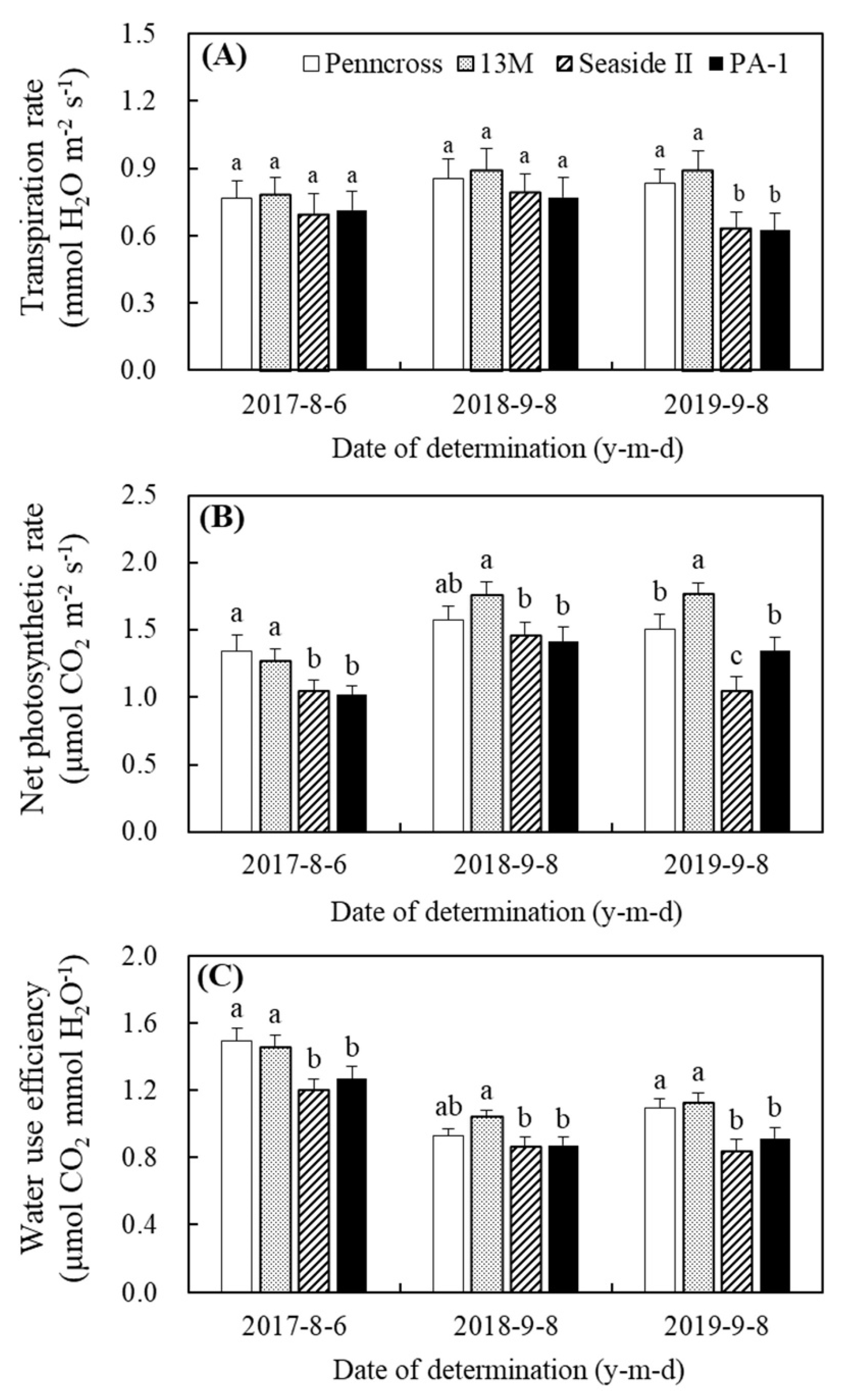
3.1. Turf Quality and Photosynthetic Parameters Affected by Summer Stress
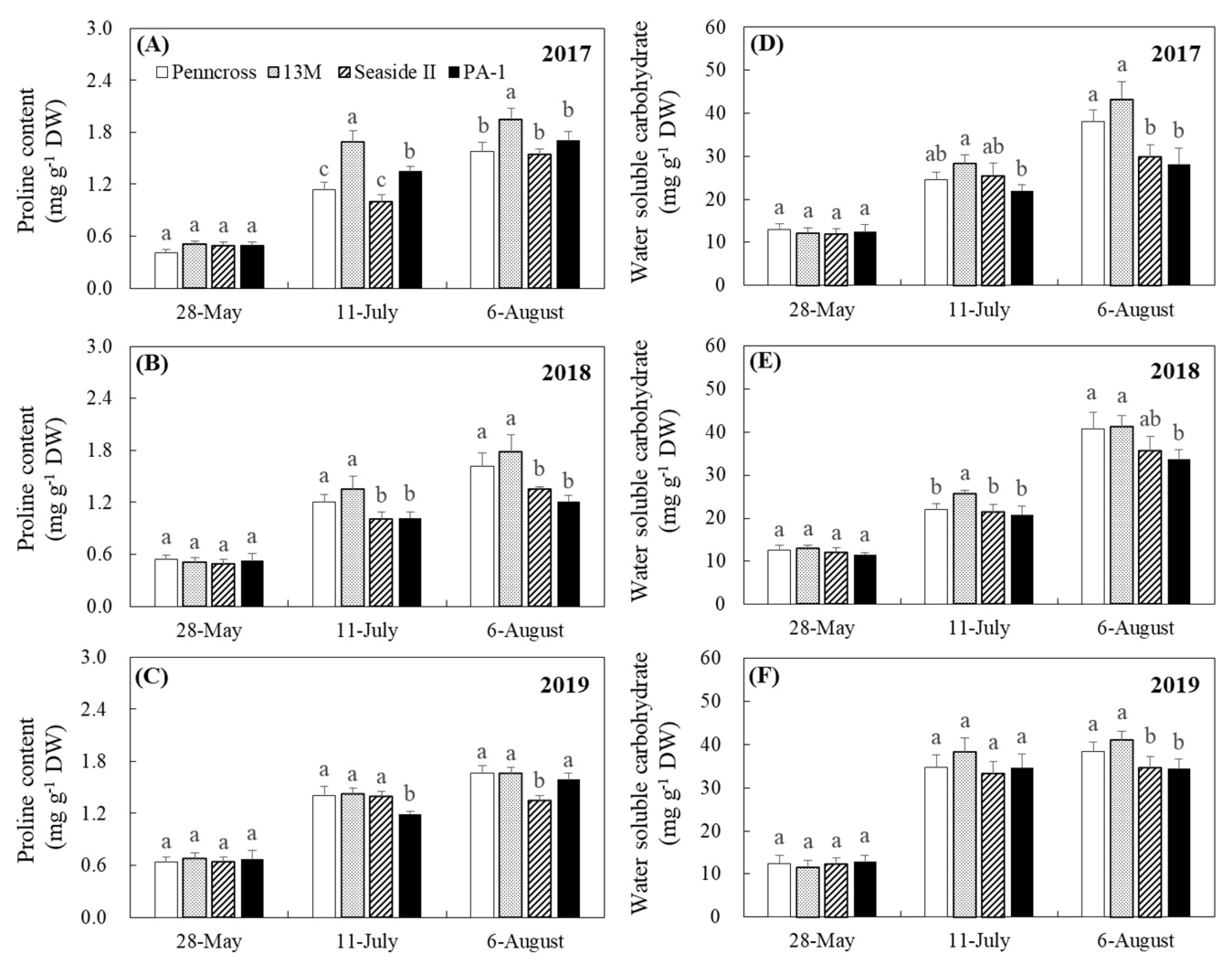
3.2. Water Status and Osmolytes Affected by Summer Stress
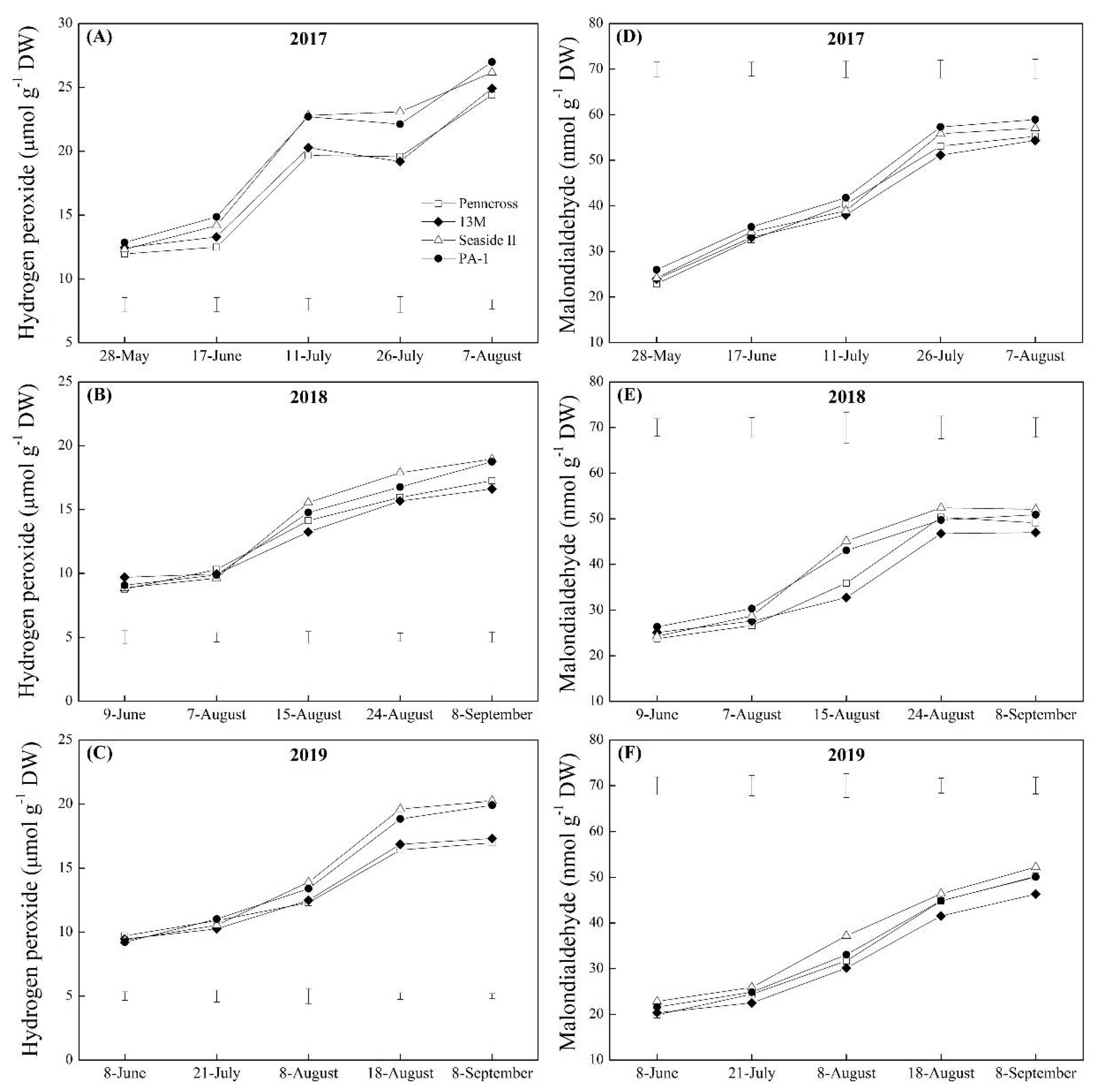
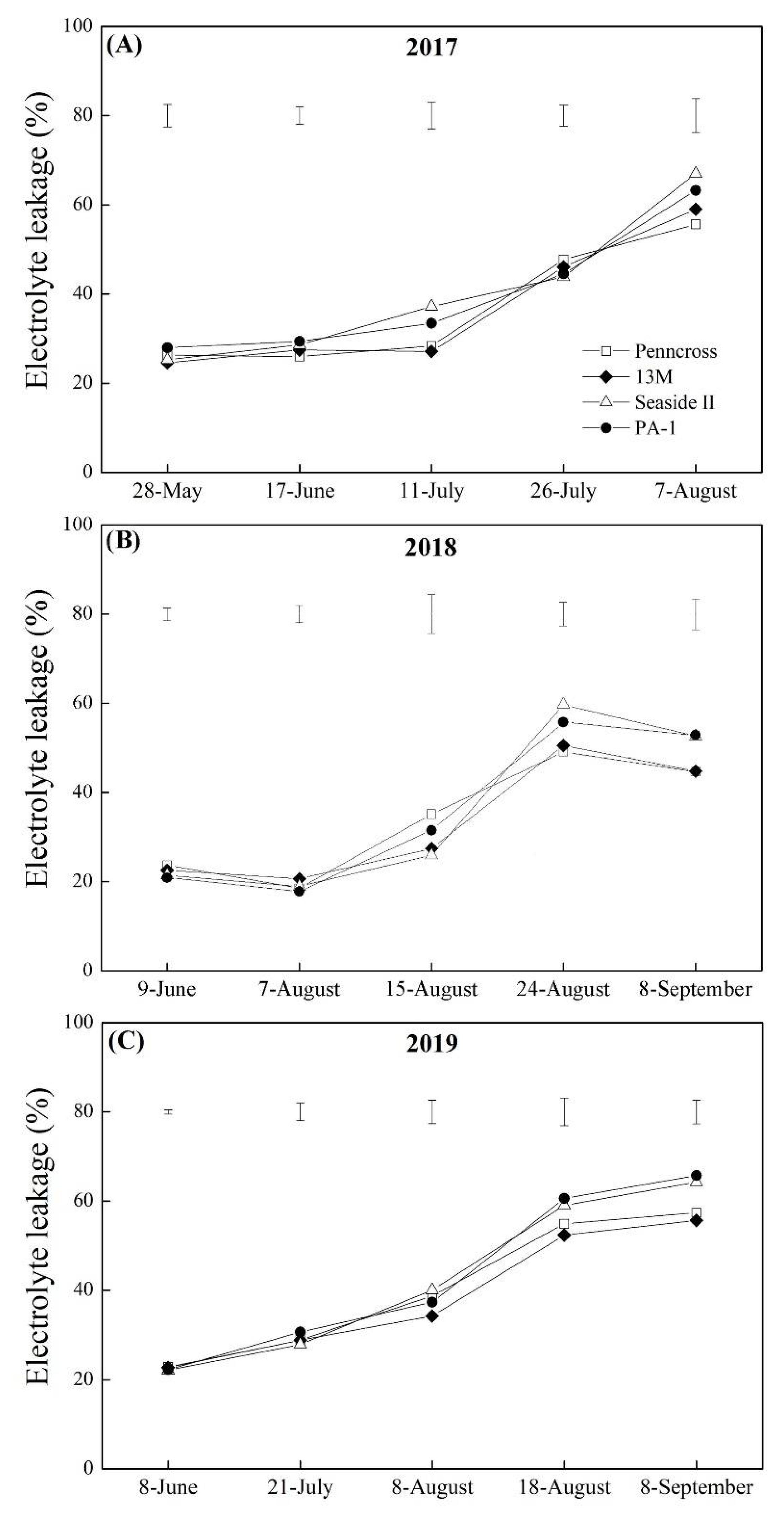
3.3. Oxidative Damage and Membrane Stability Affected by Summer Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fry, J.; Huang, B. Applied Turfgrass Science and Physiology; John Wiley Sons: Hoboken, NJ, USA, 2004. [Google Scholar]
- Xu, Q.; Huang, B. Seasonal changes in carbohydrate accumulation for two creeping bentgrass cultivars. Crop Sci. 2003, 43, 266–271. [Google Scholar] [CrossRef]
- Miller, G.L.; Brotherton, M.A. Creeping bentgrass summer decline as influenced by climatic conditions and cultural practices. Agron. J. 2020, 112, 3500–3512. [Google Scholar] [CrossRef]
- Krishnan, S.; Ma, Y.; Merewitz, E. Leaf trimming and high temperature regulation of phytohormones and polyamines in creeping bentgrass leaves. J. Am. Soc. Hortic. Sci. 2016, 141, 66–75. [Google Scholar] [CrossRef]
- Fu, J.; Dernoeden, P.H. Creeping bentgrass putting green turf responses to two irrigation practices: Quality, chlorophyll, canopy temperature, and thatch–mat. Crop Sci. 2009, 49, 1071–1078. [Google Scholar] [CrossRef] [Green Version]
- McCullough, P.E.; Liu, H.; McCarty, L.B.; Toler, J.E. Trinexapac-ethyl application regimens influence growth, quality, and performance of bermuda grass and creeping bentgrass putting greens. Crop Sci. 2007, 47, 2138–2144. [Google Scholar] [CrossRef]
- Li, Z.; Tang, M.; Hassan, M.J.; Zhang, Y.; Han, L.; Peng, Y. Adaptability to high temperature and stay-green genotypes associated with variations in antioxidant, chlorophyll metabolism, and γ-aminobutyric acid accumulation in creeping bentgrass species. Front. Plant Sci. 2021, 12, 2341. [Google Scholar] [CrossRef]
- Jespersen, D.; Ma, X.; Bonos, S.A.; Belanger, F.C.; Raymer, P.; Huang, B. Association of SSR and candidate gene markers with genetic variations in summer heat and drought performance for creeping bentgrass. Crop Sci. 2018, 58, 2644–2656. [Google Scholar] [CrossRef]
- Xu, Q.; Huang, B. Morphological and physiological characteristics associated with heat tolerance in creeping bentgrass. Crop Sci. 2001, 41, 127–133. [Google Scholar] [CrossRef]
- Huang, B.; Dacosta, M.; Jiang, Y. Research advances in mechanisms of turfgrass tolerance to abiotic stresses: From physiology to molecular biology. Crit. Rev. Plant Sci. 2014, 33, 141–189. [Google Scholar] [CrossRef]
- Yancey, P.H. Organic osmolytes as compatible, metabolic and counteracting cytoprotectants in high osmolarity and other stresses. J. Exp. Biol. 2005, 208, 2819–2830. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Liu, T.; Liang, L.; Li, Z.; Hassan, M.J.; Peng, Y.; Wang, D. Enhanced photosynthesis, carbohydrates, and energy metabolism associated with chitosan-induced drought tolerance in creeping bentgrass. Crop Sci. 2020, 60, 1064–1076. [Google Scholar] [CrossRef]
- Rossi, S.; Chapman, C.; Yuan, B.; Huang, B. Improved heat tolerance in creeping bentgrass by γ-aminobutyric acid, proline, and inorganic nitrogen associated with differential regulation of amino acid metabolism. Plant Growth Regul. 2021, 93, 231–242. [Google Scholar] [CrossRef]
- Xu, Y.; Du, H.; Huang, B. Identification of metabolites associated with superior heat tolerance in thermal bentgrass through metabolic profiling. Crop Sci. 2013, 53, 1626–1635. [Google Scholar] [CrossRef]
- Rossi, S.; Chapman, C.; Huang, B. Suppression of heat-induced leaf senescence by γ-aminobutyric acid, proline, and ammonium nitrate through regulation of chlorophyll degradation in creeping bentgrass. Environ. Exp. Bot. 2020, 177, 104116. [Google Scholar] [CrossRef]
- Liang, L.L.; Cao, Y.Q.; Wang, D.; Peng, Y.; Zhang, Y.; Li, Z. Spermine alleviates heat-induced senescence in creeping bentgrass by regulating water and oxidative balance, photosynthesis, and heat shock proteins. Biol. Plant. 2021, 65, 184–192. [Google Scholar] [CrossRef]
- Li, Z.; Yu, J.; Peng, Y.; Huang, B. Metabolic pathways regulated by γ-aminobutyric acid (GABA) contributing to heat tolerance in creeping bentgrass (Agrostis stolonifera). Sci. Rep. 2016, 6, 30338. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Huang, B. Antioxidant metabolism associated with summer leaf senescence and turf quality decline for creeping bentgrass. Crop Sci. 2004, 44, 553–560. [Google Scholar] [CrossRef]
- Zhang, X.; Ervin, E.H. Impact of seaweed extract-based cytokinins and zeatin riboside on creeping bentgrass heat tolerance. Crop Sci. 2008, 48, 364–370. [Google Scholar] [CrossRef]
- Beard, J.B. Turf Management for Golf Courses, 2nd ed.; Ann Arbor Press: Chelsea, MI, USA, 2002. [Google Scholar]
- Amnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar]
- Barrs, H.; Weatherley, P. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Blum, A. Osmotic adjustment and growth of barley genotypes under drought stress. Crop Sci. 1989, 29, 230–233. [Google Scholar] [CrossRef]
- Burgess, P.; Huang, B. Effects of sequential application of plant growth regulators and osmoregulants on drought tolerance of creeping bentgrass (Agrostis stolonifera). Crop Sci. 2013, 54, 837–844. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Jan, B.; Roel, M. An improved colorimetric method to quantify sugar content of plant tissue. J. Exp. Bot. 1993, 44, 1627–1629. [Google Scholar]
- Li, Z.; Peng, Y.; Huang, B. Physiological effects of γ-aminobutyric acid application on improving heat and drought tolerance in creeping bentgrass. J. Am. Soc. Hortic. Sci. 2016, 141, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Dhindsa, R.; Plumb-Dhindsa, P.; Thorpe, T. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Blum, A.; Ebercon, A. Cell membrane stability as a measure of drought and heat tolerance in wheat. Crop Sci. 1981, 21, 43–47. [Google Scholar] [CrossRef]
- Xin, J.; Yan, X.; Jiang, T.; Gianfagna, T.; Huang, B. Suppression of shade- or heat-induced leaf senescence in creeping bentgrass through transformation with the IPT gene for cytokinin synthesis. J. Am. Soc. Hortic. Sci. 2009, 134, 602–609. [Google Scholar]
- Jespersen, D.; Zhang, J.; Huang, B. Chlorophyll loss associated with heat-induced senescence in bentgrass. Plant Sci. 2016, 249, 1–12. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, M.; Xu, W.; Wang, Y.; Huang, K.; Zhang, C.; Wen, J. Understanding the molecular mechanism of anther development under abiotic stresses. Plant Mol. Biol. 2021, 105, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Huang, B. Carbohydrate accumulation in relation to heat stress tolerance in two creeping bentgrass cultivars. J. Am. Soc. Hortic. Sci. 2000, 125, 442–447. [Google Scholar] [CrossRef] [Green Version]
- Wahid, A.; Close, T.J. Expression of dehydrins under heat stress and their relationship with water relations of sugarcane leaves. Biol. Plant. 2007, 51, 104–109. [Google Scholar] [CrossRef]
- Zeng, W.; Hassan, M.J.; Kang, D.; Peng, Y.; Li, Z. Photosynthetic maintenance and heat shock protein accumulation relating to γ-aminobutyric acid (GABA)-regulated heat tolerance in creeping bentgrass (Agrostis stolonifera). S. Afr. J. Bot. 2021, 141, 405–413. [Google Scholar] [CrossRef]
- Dobra, J.; Motyka, V.; Dobrev, P.; Malbeck, J.; Prasil, I.T.; Haisel, D.; Gaudinova, A.; Havlova, M.; Gubis, J.; Vankova, R. Comparison of hormonal responses to heat, drought and combined stress in tobacco plants with elevated proline content. J. Plant Physiol. 2010, 167, 1360–1370. [Google Scholar] [CrossRef]
- Batcho, A.A.; Sarwar, M.B.; Rashid, B.; Hassan, S.; Husnain, T. Heat shock protein gene identified from Agave sisalana (AsHSP70) confers heat stress tolerance in transgenic cotton (Gossypium hirsutum). Theor. Exp. Plant Physiol. 2021, 33, 141–156. [Google Scholar] [CrossRef]
- Belhadj Slimen, I.; Najar, T.; Ghram, A.; Dabbebi, H.; Ben Mrad, M.; Abdrabbah, M. Reactive oxygen species, heat stress and oxidative-induced mitochondrial damage. A review. Int. J. Hyperth. 2014, 30, 513–523. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Klimov, V.V.; Los, D.A.; Carpentier, R.; Mohanty, P. Heat stress: An overview of molecular responses in photosynthesis. Photosynth. Res. 2008, 98, 541. [Google Scholar] [CrossRef]
- Hanif, S.; Saleem, M.F.; Sarwar, M.; Irshad, M.; Shakoor, A.; Wahid, M.A.; Khan, H.Z. Biochemically triggered heat and drought stress tolerance in rice by proline application. J. Plant Growth Regul. 2021, 40, 305–312. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Zeng, W.; Cheng, B.; Xu, J.; Han, L.; Peng, Y. Turf Quality and Physiological Responses to Summer Stress in Four Creeping Bentgrass Cultivars in a Subtropical Zone. Plants 2022, 11, 665. https://doi.org/10.3390/plants11050665
Li Z, Zeng W, Cheng B, Xu J, Han L, Peng Y. Turf Quality and Physiological Responses to Summer Stress in Four Creeping Bentgrass Cultivars in a Subtropical Zone. Plants. 2022; 11(5):665. https://doi.org/10.3390/plants11050665
Chicago/Turabian StyleLi, Zhou, Weihang Zeng, Bizhen Cheng, Jie Xu, Liebao Han, and Yan Peng. 2022. "Turf Quality and Physiological Responses to Summer Stress in Four Creeping Bentgrass Cultivars in a Subtropical Zone" Plants 11, no. 5: 665. https://doi.org/10.3390/plants11050665
APA StyleLi, Z., Zeng, W., Cheng, B., Xu, J., Han, L., & Peng, Y. (2022). Turf Quality and Physiological Responses to Summer Stress in Four Creeping Bentgrass Cultivars in a Subtropical Zone. Plants, 11(5), 665. https://doi.org/10.3390/plants11050665