
Micro-RNA-Regulated SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE (SPL) Gene Expression and Cytokinin Accumulation Distinguish Early-Developing Male and Female Inflorescences in Oil Palm (Elaeis guineensis)

Abstract
:1. Introduction
2. Results
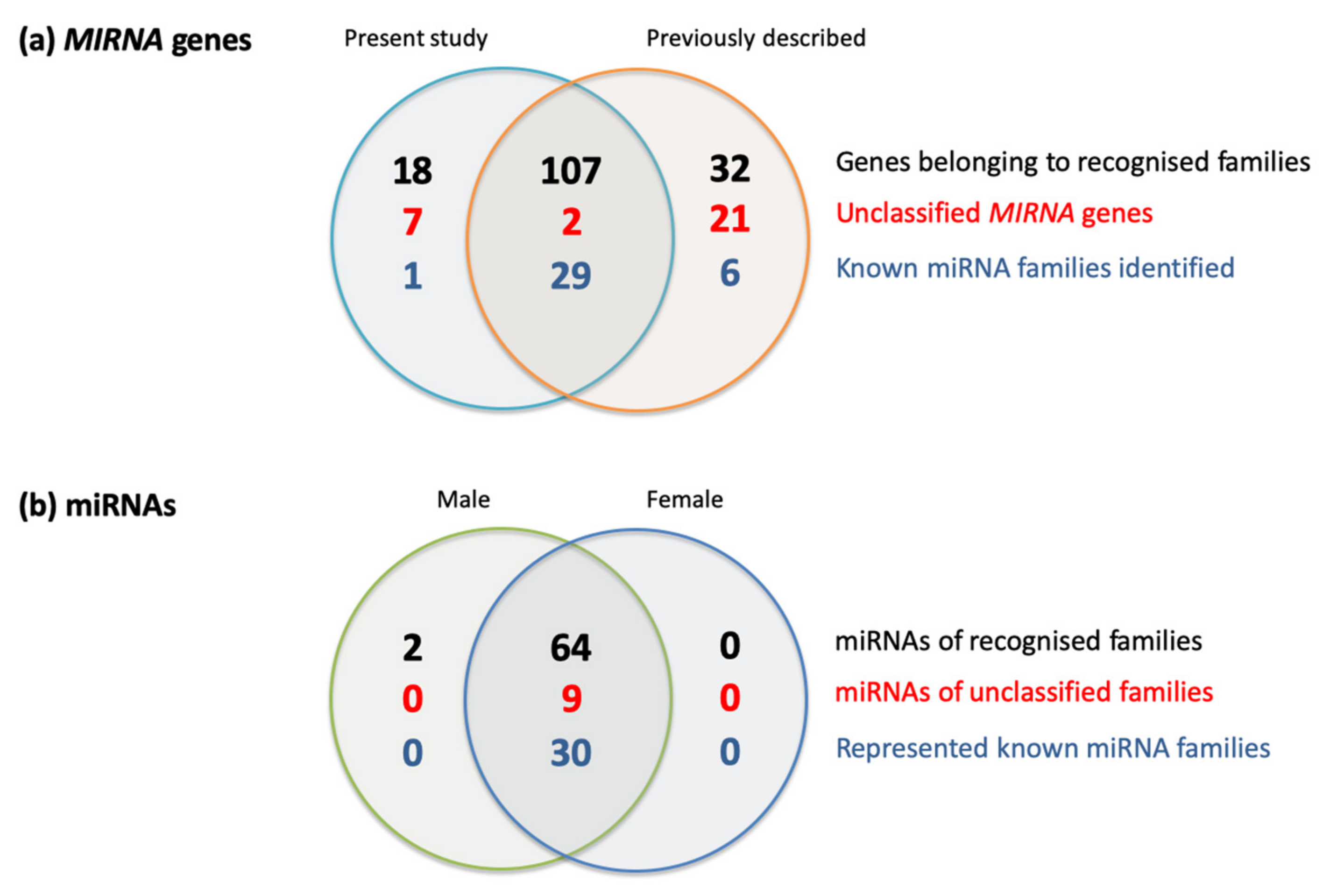
2.1. Characterisation of miRNAs Accumulating in the Immature Oil Palm Inflorescence
2.2. Gene Expression Patterns Distinguish Early Developing Male and Female Oil Palm Inflorescences
2.3. Key Regulatory Genes Displaying Sex-Dependent Expression Are miRNA-Regulated
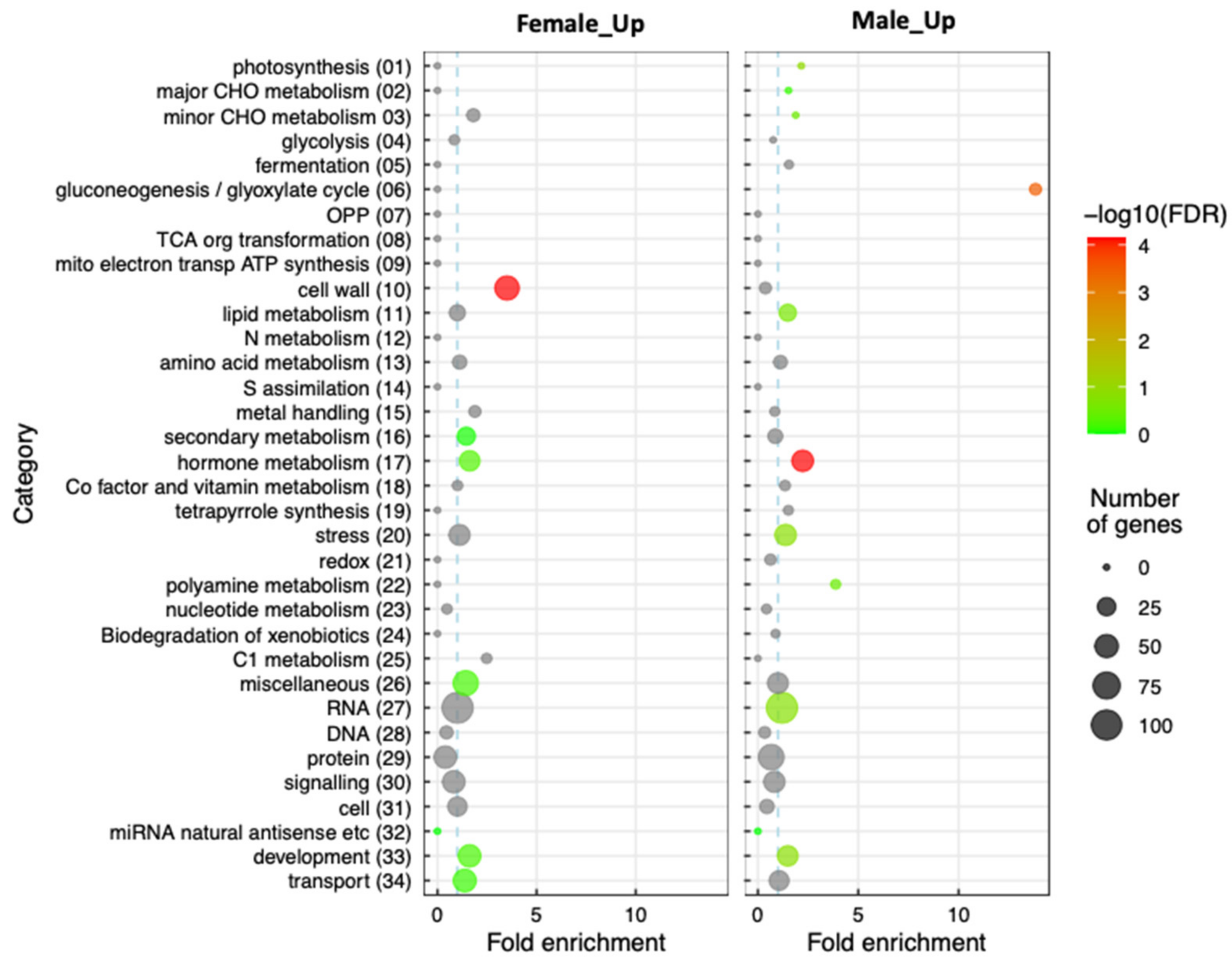
2.4. Gene Enrichment Analysis Reveals the Importance of Hormonal Regulation during Inflorescence Sexual Differentiation
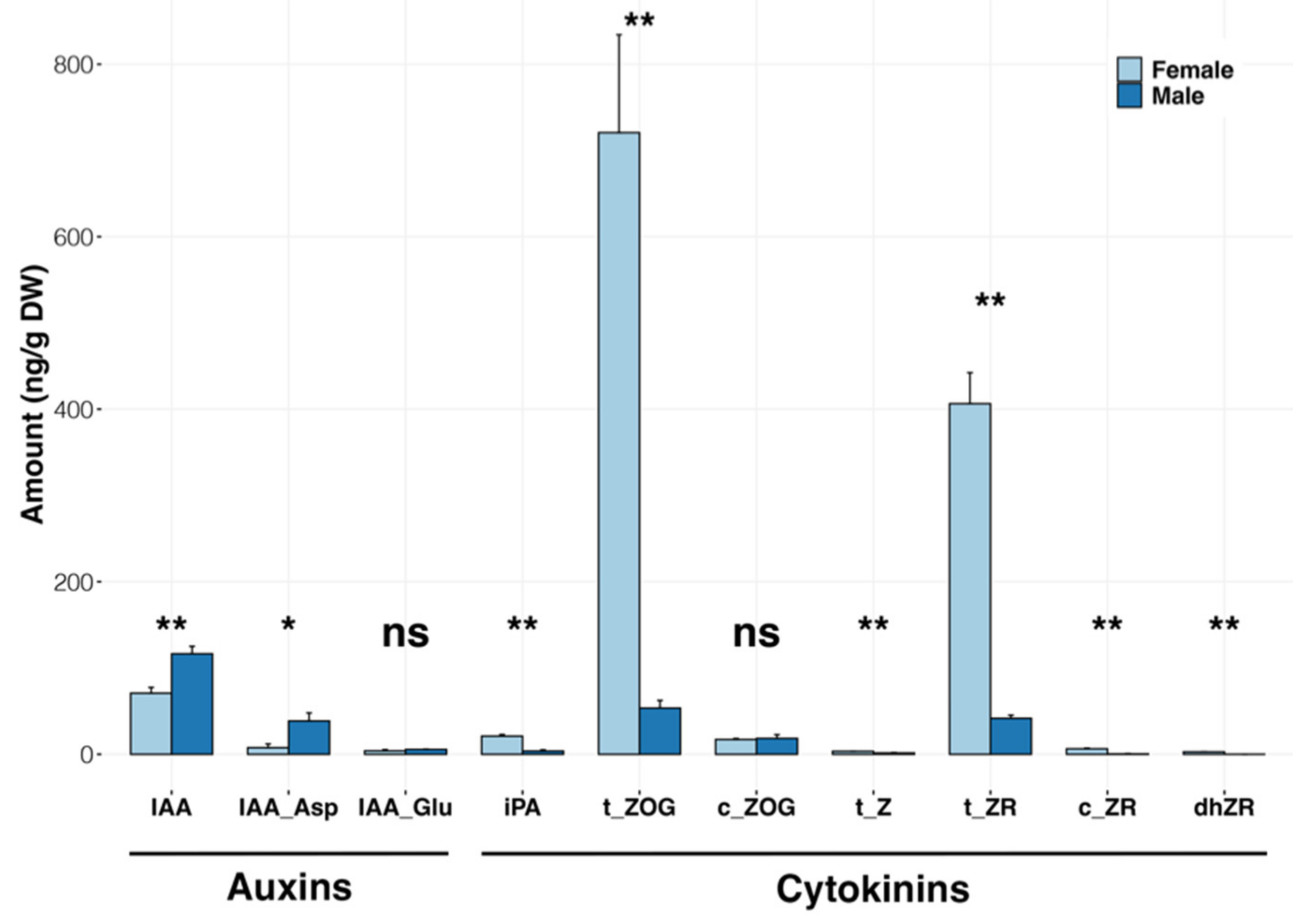
2.5. Analysis of Hormone Composition in Immature Male and Female Inflorescences
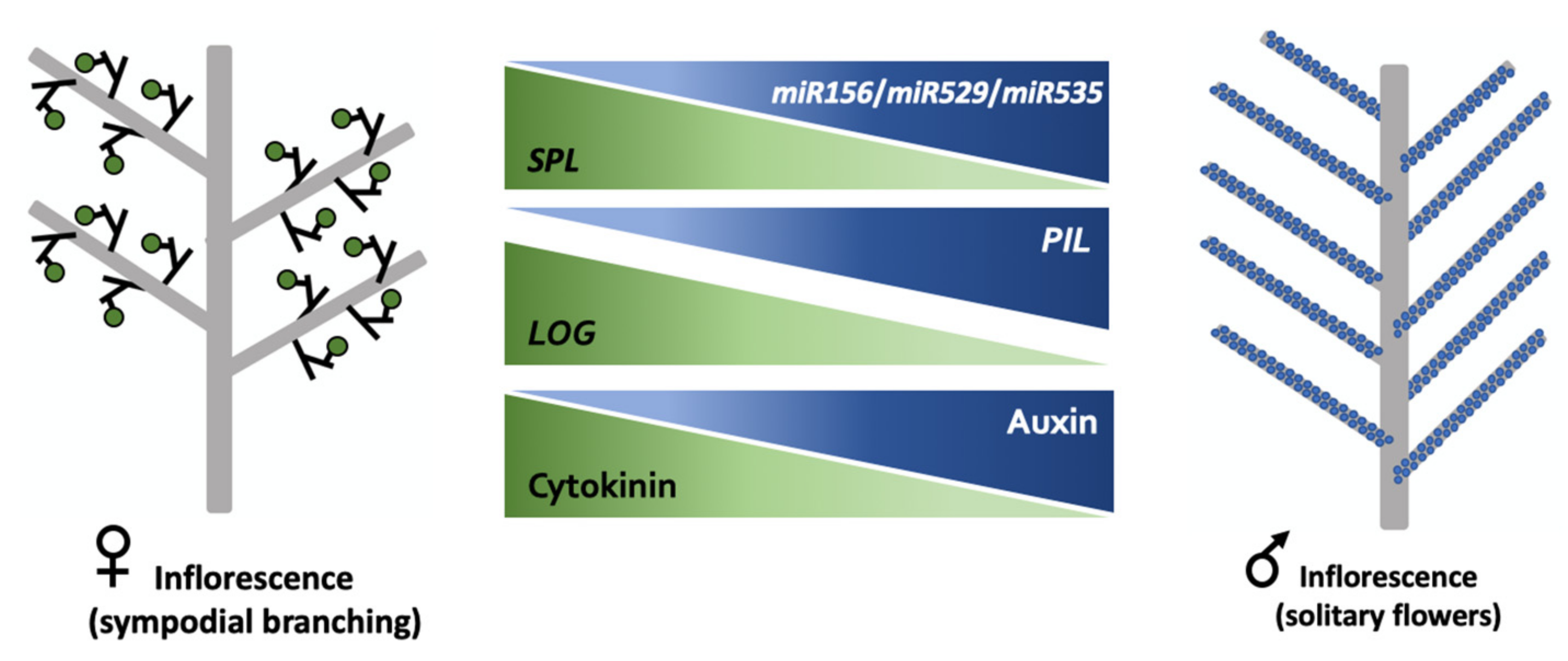
3. Discussion
3.1. Post-Transcriptional miRNA-mRNA Modules Are Likely to Be Involved in Sex-Dependent Regulatory Processes in the Developing Oil Palm Inflorescence
3.2. Global Expression and Biochemical Profiles Indicate a Key Role for Hormones in Oil Palm Inflorescence Sexual Differentiation
4. Materials and Methods
4.1. Plant Material
4.2. Extraction of Small RNAs from Oil Palm Inflorescence and Analysis of miRNAs
4.3. Extraction of Total RNAs and Analysis of Differential Gene Expression
4.4. Functional Annotation of Genes and Identification of Enriched Categories
4.5. Analysis of Molecular Phylogeny
4.6. Analysis of Hormone Composition in Immature Oil Palm Inflorescences
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hill, C.B.; Li, C. Genetic Architecture of Flowering Phenology in Cereals and Opportunities for Crop Improvement. Front. Plant Sci. 2016, 7, 1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Yang, X.; Li, G. Molecular Insights into Inflorescence Meristem Specification for Yield Potential in Cereal Crops. Int. J. Mol. Sci. 2021, 22, 3508. [Google Scholar] [CrossRef]
- Wils, C.R.; Kaufmann, K. Gene-regulatory networks controlling inflorescence and flower development in Arabidopsis thaliana. Biochim. Biophys. Acta Gene Regul. Mech. 2017, 1860, 95–105. [Google Scholar] [CrossRef]
- Axtell, M.J.; Meyers, B.C. Revisiting Criteria for Plant MicroRNA Annotation in the Era of Big Data. Plant Cell 2018, 30, 272–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, K.; Chen, X. Biogenesis, turnover, and mode of action of plant microRNAs. Plant Cell 2013, 25, 2383–2399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achkar, N.P.; Cambiagno, D.A.; Manavella, P.A. miRNA Biogenesis: A Dynamic Pathway. Trends Plant Sci. 2016, 21, 1034–1044. [Google Scholar] [CrossRef]
- Yu, Y.; Jia, T.; Chen, X. The ‘how’ and ‘where’ of plant microRNAs. New Phytol. 2017, 216, 1002–1017. [Google Scholar] [CrossRef] [Green Version]
- Chuck, G.; Meeley, R.; Irish, E.; Sakai, H.; Hake, S. The maize tasselseed4 microRNA controls sex determination and meristem cell fate by targeting Tasselseed6/indeterminate spikelet1. Nat. Genet. 2007, 39, 1517–1521. [Google Scholar] [CrossRef]
- Zheng, Y.; Jagadeeswaran, G.; Gowdu, K.; Wang, N.; Li, S.; Ming, R.; Sunkar, R. Genome-Wide Analysis of MicroRNAs in Sacred Lotus, Nelumbo nucifera (Gaertn). Trop. Plant Biol. 2013, 6, 117–130. [Google Scholar] [CrossRef]
- Miura, K.; Ikeda, M.; Matsubara, A.; Song, X.-J.; Ito, M.; Asano, K.; Matsuoka, M.; Kitano, H.; Ashikari, M. OsSPL14 promotes panicle branching and higher grain productivity in rice. Nat. Genet. 2010, 42, 545–549. [Google Scholar] [CrossRef]
- Jiao, Y.; Wang, Y.; Xue, D.; Wang, J.; Yan, M.; Liu, G.; Dong, G.; Zeng, D.; Lu, Z.; Zhu, X.; et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nat. Genet. 2010, 42, 541–544. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Wei, M.; Li, Y.; Tao, H.; Wu, H.; Chen, Z.; Li, C.; Xu, J.H. MiR529a controls plant height, tiller number, panicle architecture and grain size by regulating SPL target genes in rice (Oryza sativa L.). Plant Sci. 2021, 302, 110728. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Shen, Y.; Li, H.; Yang, J.; Cai, X.; Zheng, G.; Zhu, Y.; Jia, B.; Sun, X. The multiple roles of OsmiR535 in modulating plant height, panicle branching and grain shape. Plant Sci. 2019, 283, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Yue, E.; Cao, H.; Liu, B. OsmiR535, a Potential Genetic Editing Target for Drought and Salinity Stress Tolerance in Oryza sativa. Plants 2020, 9, 1337. [Google Scholar] [CrossRef] [PubMed]
- Dransfield, J.; Baker, W.J.; Harley, M.M.; Asmussen, C.; Lewis, C.E. Genera Palmarum: The Evolution & Classification of Palms; Kew Publishing: London, UK, 2008. [Google Scholar]
- Nadot, S.; Alapetite, E.; Baker, W.J.; Tregear, J.W.; Barfod, A.S. The palm family (Arecaceae): A microcosm of sexual system evolution. Bot. J. Linn. Soc. 2016, 182, 376–388. [Google Scholar] [CrossRef] [Green Version]
- Adam, H.; Jouannic, S.; Escoute, J.; Duval, Y.; Verdeil, J.L.; Tregear, J.W. Reproductive developmental complexity in the African oil palm (Elaeis guineensis). Am. J. Bot. 2005, 92, 1836–1852. [Google Scholar] [CrossRef]
- Durand-Gasselin, T.; Noiret, J.M.; Kouamé, R.K.; Cochard, B.; Adon, B. Availability of quality pollen for improved oil palm (Elaeis guineensis Jacq.) seed production. Plant. Rech. Développement 1999, 6, 264–276. [Google Scholar]
- Adam, H.; Collin, M.; Richaud, F.; Beulé, T.; Cros, D.; Omore, A.; Nodichao, L.; Nouy, B.; Tregear, J.W. Environmental regulation of sex determination in oil palm: Current knowledge and insights from other species. Ann. Bot. 2011, 108, 1529–1537. [Google Scholar] [CrossRef] [Green Version]
- Van Heel, W.A.; Breure, C.J.; Menendez, T. The early development of inflorescences and flowers of oil palm (Elaeis guineensis Jacq.) seen through the scanning electron microscope. Blumea 1987, 32, 67–78. [Google Scholar]
- Chen, C.; Li, J.; Feng, J.; Liu, B.; Feng, L.; Yu, X.; Li, G.; Zhai, J.; Meyers, B.C.; Xia, R. sRNAanno-a database repository of uniformly annotated small RNAs in plants. Hortic. Res. 2021, 8, 45. [Google Scholar] [CrossRef]
- Dai, X.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39, W155–W159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preston, J.C.; Hileman, L.C. Functional Evolution in the Plant SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE (SPL) Gene Family. Front. Plant Sci. 2013, 4, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adam, H.; Jouannic, S.; Morcillo, F.; Richaud, F.; Duval, Y.; Tregear, J.W. MADS box genes in oil palm (Elaeis guineensis): Patterns in the evolution of the SQUAMOSA, DEFICIENS, GLOBOSA, AGAMOUS and SEPALLATA subfamilies. J. Mol. Evol. 2006, 62, 15–31. [Google Scholar] [CrossRef]
- Aceto, S.; Sica, M.; De Paolo, S.; D’Argenio, V.; Cantiello, P.; Salvatore, F.; Gaudio, L. The analysis of the inflorescence miRNome of the orchid Orchis italica reveals a DEF-like MADS-box gene as a new miRNA target. PLoS ONE 2014, 9, e97839. [Google Scholar] [CrossRef]
- Li, X.; Jin, F.; Jin, L.; Jackson, A.; Ma, X.; Shu, X.; Wu, D.; Jin, G. Characterization and comparative profiling of the small RNA transcriptomes in two phases of flowering in Cymbidium ensifolium. BMC Genom. 2015, 16, 622. [Google Scholar] [CrossRef] [Green Version]
- Sabana, A.A.; Rajesh, M.K.; Antony, G. Dynamic changes in the expression pattern of miRNAs and associated target genes during coconut somatic embryogenesis. Planta 2020, 251, 79. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, T.; Wang, R.; Zhao, Y. Recent advances in auxin research in rice and their implications for crop improvement. J. Exp. Bot. 2018, 69, 255–263. [Google Scholar] [CrossRef]
- Li, S.B.; Xie, Z.Z.; Hu, C.G.; Zhang, J.Z. A Review of Auxin Response Factors (ARFs) in Plants. Front. Plant Sci. 2016, 7, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohse, M.; Nagel, A.; Herter, T.; May, P.; Schroda, M.; Zrenner, R.; Tohge, T.; Fernie, A.R.; Stitt, M.; Usadel, B. Mercator: A fast and simple web server for genome scale functional annotation of plant sequence data. Plant Cell Environ. 2014, 37, 1250–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnot, T.; Gillard, M.B.; Nagel, D.H. A Simple Protocol for Informative Visualization of Enriched Gene Ontology Terms. Bio Protocol 2019, 9, e3429. [Google Scholar] [CrossRef]
- Lu, Z.; Yu, H.; Xiong, G.; Wang, J.; Jiao, Y.; Liu, G.; Jing, Y.; Meng, X.; Hu, X.; Qian, Q.; et al. Genome-wide binding analysis of the transcription activator ideal plant architecture1 reveals a complex network regulating rice plant architecture. Plant Cell 2013, 25, 3743–3759. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yu, H.; Xiong, G.; Lu, Z.; Jiao, Y.; Meng, X.; Liu, G.; Chen, X.; Wang, Y.; Li, J. Tissue-Specific Ubiquitination by IPA1 INTERACTING PROTEIN1 Modulates IPA1 Protein Levels to Regulate Plant Architecture in Rice. Plant Cell 2017, 29, 697–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; He, G.; Li, Y.; Yang, Z.; Liu, T.; Xie, X.; Kong, X.; Sun, J. PIL transcription factors directly interact with SPLs and repress tillering/branching in plants. New Phytol. 2021, 233, 1414–1425. [Google Scholar] [CrossRef] [PubMed]
- Adam, H.; Jouannic, S.; Morcillo, F.; Orieux, Y.; Duval, Y.; Tregear, J.W. Functional characterization of MADS box genes involved in the determination of oil palm flower structure. J. Exp. Bot. 2007, 58, 1245–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong-Abdullah, M.; Ordway, J.M.; Jiang, N.; Ooi, S.E.; Kok, S.Y.; Sarpan, N.; Azimi, N.; Hashim, A.T.; Ishak, Z.; Rosli, S.K.; et al. Loss of Karma transposon methylation underlies the mantled somaclonal variant of oil palm. Nature 2015, 525, 533–537. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yuan, Z.; Meng, Q.; Huang, G.; Perin, C.; Bureau, C.; Meunier, A.C.; Ingouff, M.; Bennett, M.J.; Liang, W.; et al. Dynamic Regulation of Auxin Response during Rice Development Revealed by Newly Established Hormone Biosensor Markers. Front. Plant Sci. 2017, 8, 256. [Google Scholar] [CrossRef] [Green Version]
- Weijers, D.; Wagner, D. Transcriptional Responses to the Auxin Hormone. Annu. Rev. Plant Biol. 2016, 67, 539–574. [Google Scholar] [CrossRef]
- Sims, K.; Abedi-Samakush, F.; Szulc, N.; Macias Honti, M.G.; Mattsson, J. OsARF11 Promotes Growth, Meristem, Seed, and Vein Formation during Rice Plant Development. Int. J. Mol. Sci. 2021, 22, 4089. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Yu, Y.; Wang, C.Y.; Li, Z.Y.; Liu, Q.; Xu, J.; Liao, J.Y.; Wang, X.J.; Qu, L.H.; Chen, F.; et al. Overexpression of microRNA OsmiR397 improves rice yield by increasing grain size and promoting panicle branching. Nat. Biotechnol. 2013, 31, 848–852. [Google Scholar] [CrossRef]
- Swetha, C.; Basu, D.; Pachamuthu, K.; Tirumalai, V.; Nair, A.; Prasad, M.; Shivaprasad, P.V. Major Domestication-Related Phenotypes in Indica Rice Are Due to Loss of miRNA-Mediated Laccase Silencing. Plant Cell 2018, 30, 2649–2662. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Dong, H.; Wang, M.; Liu, Q. Evolutionary divergence of function and expression of laccase genes in plants. J. Genet. 2020, 99, 23. [Google Scholar] [CrossRef] [PubMed]
- Chuck, G.; Meeley, R.; Hake, S. Floral meristem initiation and meristem cell fate are regulated by the maize AP2 genes ids1 and sid1. Development 2008, 135, 3013–3019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, M.R.; Lou, H.; Aubert, M.K.; Wilkinson, L.G.; Little, A.; Houston, K.; Pinto, S.C.; Shirley, N.J. Exploring the Role of Cell Wall-Related Genes and Polysaccharides during Plant Development. Plants 2018, 7, 42. [Google Scholar] [CrossRef] [Green Version]
- Landschutze, V.; Willmitzer, L.; Muller-Rober, B. Inhibition of flower formation by antisense repression of mitochondrial citrate synthase in transgenic potato plants leads to a specific disintegration of the ovary tissues of flowers. EMBO J. 1995, 14, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Malone, S.; Chen, Z.-H.; Bahrami, A.R.; Walker, R.P.; Gray, J.E.; Leegood, R.C. Phospho enol pyruvate Carboxykinase in Arabidopsis: Changes in Gene Expression, Protein and Activity during Vegetative and Reproductive Development. Plant Cell Physiol. 2007, 48, 441–450. [Google Scholar] [CrossRef]
- Kieber, J.J. Cytokinins. Arab. Book 2002, 1, e0063. [Google Scholar] [CrossRef]
- Hirose, N.; Makita, N.; Kojima, M.; Kamada-Nobusada, T.; Sakakibara, H. Overexpression of a Type-A Response Regulator Alters Rice Morphology and Cytokinin Metabolism. Plant Cell Physiol. 2007, 48, 523–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashikari, M.; Sakakibara, H.; Lin, S.; Yamamoto, T.; Takashi, T.; Nishimura, A.; Angeles, E.R.; Qian, Q.; Kitano, H.; Matsuoka, M. Cytokinin Oxidase Regulates Rice Grain Production. Science 2005, 309, 741–745. [Google Scholar] [CrossRef]
- Duan, J.; Yu, H.; Yuan, K.; Liao, Z.; Meng, X.; Jing, Y.; Liu, G.; Chu, J.; Li, J. Strigolactone promotes cytokinin degradation through transcriptional activation of CYTOKININ OXIDASE/DEHYDROGENASE 9 in rice. Proc. Natl. Acad. Sci. USA 2019, 116, 14319–14324. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Liu, L.; Li, M.; Fang, S.; Shen, X.; Chu, J.; Zhang, Z. UNBRANCHED3 regulates branching by modulating cytokinin biosynthesis and signaling in maize and rice. New Phytol. 2017, 214, 721–733. [Google Scholar] [CrossRef] [Green Version]
- Kurepa, J.; Shull, T.E.; Smalle, J.A. Antagonistic activity of auxin and cytokinin in shoot and root organs. Plant Direct 2019, 3, e00121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotov, A.A.; Kotova, L.M.; Romanov, G.A. Signaling network regulating plant branching: Recent advances and new challenges. Plant Sci. 2021, 307, 110880. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shi, X.a.; Chang, Z.; Ding, Y.; Ding, C. Auxin Efflux Transporters OsPIN1c and OsPIN1d Function Redundantly in Regulating Rice (Oryza sativa L.) Panicle Development. Plant Cell Physiol 2021, pcab172. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.; Wang, L.; Li, Y.; Chen, L.; Li, S.; Du, X.; Wang, B.; Yan, J.; Li, J.; Xie, D. Rice DWARF14 acts as an unconventional hormone receptor for strigolactone. J. Exp. Bot. 2018, 69, 2355–2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarra, R.; Jin, L.; Zhao, Z.; Cao, H. Progress in Tissue Culture and Genetic Transformation of Oil Palm: An Overview. Int. J. Mol. Sci. 2019, 20, 5353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeap, W.-C.; Norkhairunnisa Che Mohd, K.; Norfadzilah, J.; Muad, M.R.; Appleton, D.R.; Harikrishna, K. An Efficient Clustered Regularly Interspaced Short Palindromic Repeat (CRISPR)/CRISPR-Associated Protein 9 Mutagenesis System for Oil Palm (Elaeis guineensis). Front. Plant Sci. 2021, 12, 773656. [Google Scholar] [CrossRef]
- Bahariah, B.; Masani, M.Y.A.; Rasid, O.A.; Parveez, G.K.A. Multiplex CRISPR/Cas9-mediated genome editing of the FAD2 gene in rice: A model genome editing system for oil palm. J. Genet. Eng. Biotechnol. 2021, 19, 86. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Fallmann, J.; Videm, P.; Bagnacani, A.; Batut, B.; Doyle, M.A.; Klingstrom, T.; Eggenhofer, F.; Stadler, P.F.; Backofen, R.; Grüning, B. The RNA workbench 2.0: Next generation RNA data analysis. Nucleic Acids Res. 2019, 47, W511–W515. [Google Scholar] [CrossRef] [Green Version]
- Afgan, E.; Baker, D.; van den Beek, M.; Blankenberg, D.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Eberhard, C.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2016 update. Nucleic Acids Res. 2016, 44, W3–W10. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Ge, S.X.; Son, E.W.; Yao, R. iDEP: An integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinform. 2018, 19, 534. [Google Scholar] [CrossRef] [Green Version]
- Klie, S.; Nikoloski, Z. The choice between Mapman and Gene Ontology for automated gene function prediction in plant science. Front. Genet. 2012, 3, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed]
- Chiwocha, S.D.; Abrams, S.R.; Ambrose, S.J.; Cutler, A.J.; Loewen, M.; Ross, A.R.; Kermode, A.R. A method for profiling classes of plant hormones and their metabolites using liquid chromatography-electrospray ionization tandem mass spectrometry: An analysis of hormone regulation of thermodormancy of lettuce (Lactuca sativa L.) seeds. Plant J. 2003, 35, 405–417. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA_ID | Sequence | M/F Ratio of miRNA | Type of Target | DEG mRNA Target | M/F Ratio of DEGs |
|---|---|---|---|---|---|
| miR156d | TGACAGAAGAGAGTGAGCACC | ∞ | SBP | EgSPL14-1 (LOC105050210) EgSPL14-2 (LOC105061255) EgSPL143-3 (LOC105050485) | 0.1806 0.3275 0.4332 |
| miR529a | AGAAGAGAGAGAGTACAGCCT | 8.2980 | SBP | EgSPL14-1 (LOC105050210) EgSPL14-2 (LOC105061255) EgSPL14-3 (LOC105050485) | 0.1806 0.3275 0.4332 |
| miR156a | CTGACAGAAGAGAGTGAGCAC | 8.1495 | SBP | EgSPL14-1 (LOC105050210) EgSPL14-2 (LOC105061255) EgSPL14-3 (LOC105050485) | 0.1806 0.3275 0.4332 |
| miR535a | TGACAACGAGAGAGAGCACGC | 7.2660 | SBP | EgSPL14-1 (LOC105050210) EgSPL14-2 (LOC105061255) EgSPL14-3 (LOC105050485) | 0.1806 0.3275 0.4332 |
| miR156b | TTGACAGAAGATAGAGAGCAC | 7.2342 | SBP | EgSPL14-1 (LOC105050210) EgSPL14-2 (LOC105061255) EgSPL14-3 (LOC105050485) | 0.1806 0.3275 0.4332 |
| miR5179 | TTTTGCTCAAGACCGCGCAAC | 4.3972 | DEF | EgDEF1 (ID105033334) | 0.0307 |
| miR167b | TGAAGCTGCCAGCATGATCTA | 3.5564 | ARF | LOC105034684 LOC105055363 | 0.3410 0.3620 |
| miR167a | TGAAGCTGCCAGCATGATCT | 3.2110 | ARF | LOC105055363 | 0.3620 |
| miR164a | TGGAGAAGCAGGGCACGTGCA | 2.6633 | NAC | LOC105058561 LOC105056634 | 3.3287 22.0167 |
| miR397b | TCATTGAGTGCAGCGTTGATG | 2.4699 | Laccase | LOC105036954 | 14.0474 |
| miR397a | TCATCGAGTGCAGCGTTGATG | 0.4993 | Laccase | LOC105036954 | 14.0474 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tregear, J.W.; Richaud, F.; Collin, M.; Esbelin, J.; Parrinello, H.; Cochard, B.; Nodichao, L.; Morcillo, F.; Adam, H.; Jouannic, S. Micro-RNA-Regulated SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE (SPL) Gene Expression and Cytokinin Accumulation Distinguish Early-Developing Male and Female Inflorescences in Oil Palm (Elaeis guineensis). Plants 2022, 11, 685. https://doi.org/10.3390/plants11050685
Tregear JW, Richaud F, Collin M, Esbelin J, Parrinello H, Cochard B, Nodichao L, Morcillo F, Adam H, Jouannic S. Micro-RNA-Regulated SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE (SPL) Gene Expression and Cytokinin Accumulation Distinguish Early-Developing Male and Female Inflorescences in Oil Palm (Elaeis guineensis). Plants. 2022; 11(5):685. https://doi.org/10.3390/plants11050685
Chicago/Turabian StyleTregear, James W., Frédérique Richaud, Myriam Collin, Jennifer Esbelin, Hugues Parrinello, Benoît Cochard, Leifi Nodichao, Fabienne Morcillo, Hélène Adam, and Stefan Jouannic. 2022. "Micro-RNA-Regulated SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE (SPL) Gene Expression and Cytokinin Accumulation Distinguish Early-Developing Male and Female Inflorescences in Oil Palm (Elaeis guineensis)" Plants 11, no. 5: 685. https://doi.org/10.3390/plants11050685
APA StyleTregear, J. W., Richaud, F., Collin, M., Esbelin, J., Parrinello, H., Cochard, B., Nodichao, L., Morcillo, F., Adam, H., & Jouannic, S. (2022). Micro-RNA-Regulated SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE (SPL) Gene Expression and Cytokinin Accumulation Distinguish Early-Developing Male and Female Inflorescences in Oil Palm (Elaeis guineensis). Plants, 11(5), 685. https://doi.org/10.3390/plants11050685