
Light and Plant Growth Regulators on In Vitro Proliferation



Abstract
:1. Introduction
2. Effects of Spectral Quality of Light on In Vitro Proliferation
2.1. Red Light Effects
2.1.1. Red Light Effects on Shoot Proliferation
2.1.2. Red Light Effects on Shoot Morphology
2.2. Blue Light Effects
2.2.1. Blue Light Effects on Shoot Proliferation
2.2.2. Blue Light Effects on Plantlet Morphology
2.3. Combined Blue and Red Light Effects
2.3.1. Blue and Red Light Effects on Shoot Proliferation
2.3.2. Blue and Red Light Effects on Plantlet Morphology
2.4. White Light Effects
2.4.1. White Light Effects on Shoot Proliferation
2.4.2. White Light Effects on Plantlet Morphology
2.5. Green Light Effects on Shoot Proliferation and Plantlet Morphology
2.6. Yellow Light Effects on Shoot Proliferation and Plantlet Morphology
2.7. Far Red Light Effects on Shoot Proliferation
3. Effects of Light Intensity
4. Effects of Photoperiod
5. Light and Plant Growth Regulators
5.1. Light Effects on Endogenous Growth Regulators
5.2. Effects of Light on Exogenous Applied Growth Regulators
6. Discussion and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Smith, H. Sensing the light environment: The functions of the phytochrome family. In Photomorphogenesis in Plants; Kendrick, R.E., Kronenberg, G.H.M., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 377–416. [Google Scholar]
- Morini, S.; Muleo, R. Il ruolo della qualità della luce nei processi di sviluppo e differenziazione delle colture in vitro. Italus Hortus Rev. 2012, 19, 37–49. [Google Scholar]
- Kozai, T. Why LED lighting for urban agriculture? In LED Lighting for Urban Agriculture; Kozai, T., Fujiwara, K., Runkle, E., Eds.; Springer: Singapore, 2016; pp. 3–18. [Google Scholar] [CrossRef]
- Whitelam, G.C.; Halliday, K.J. Photomorphogenesis: Phytochrome takes a partner! Curr. Biol. 1999, 9, 225–227. [Google Scholar] [CrossRef] [Green Version]
- Muneer, S.; Kim, E.J.; Park, J.S. Influence of green, red and blue light emitting diodes on multiprotein complex proteins and photosynthetic activity under different light intensities in lettuce leaves (Lactuca sativa L.). Int. J. Mol. Sci. 2014, 15, 4657–4670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higuchi, Y.; Hisamatsu, T. Light acts as a signal for regulation of growth and development. In LED Lighting for Urban Agriculture; Kozai, T., Fujiwara, K., Runkle, E., Eds.; Springer: Singapore, 2016; pp. 57–73. [Google Scholar]
- Pedmale, U.V.; Huang, S.C.; Zander, M.; Cole, B.J.; Hetzel, J.; Ljung, K.; Chory, J. Cryptochromes interact directly with PIFs to control plant growth in limiting blue light. Cell 2016, 164, 233–245. [Google Scholar] [CrossRef] [Green Version]
- Li, C.-X.; Xu, Z.-G.; Dong, R.-Q.; Chang, S.-X.; Wang, L.-Z.; Khalil-Ur-Rehman, M.; Tao, J.-M. An RNA-Seq analysis of grape plantlets grown in vitro reveals different responses to blue, green, red led light, and white fluorescent light. Front. Plant Sci. 2017, 8, 78. [Google Scholar] [CrossRef] [Green Version]
- Batista, D.S.; Felipe, S.H.S.; Silva, T.D.; de Castro, K.M.; Mamedes-Rodrigues, T.C.; Miranda, N.A.; Ríos-Ríos, A.M.; Faria, D.V.; Fortini, E.A.; Chagas, K.; et al. Light quality in plant tissue culture: Does it matter? Vitr. Cell. Dev. Biol. Plant 2018, 54, 195–215. [Google Scholar] [CrossRef]
- Chen, M.; Chory, J.; Fankhauser, C. Light signal transduction in higher plants. Annu. Rev. Genet. 2004, 38, 87–117. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, R.; Batschauer, A. Plant blue-light receptors. Planta 2005, 220, 498–502. [Google Scholar] [CrossRef]
- Chen, M.; Chory, J. Phytochrome signaling mechanisms and the control of plant development. Trends Cell Biol. 2011, 21, 664–671. [Google Scholar] [CrossRef] [Green Version]
- Vierstra, R.D.; Zhang, J. Phytochrome signaling: Solving the Gordian knot with microbial relatives. Trends Plant Sci. 2011, 16, 417–426. [Google Scholar] [CrossRef]
- Galvão, V.C.; Fankhauser, C. Sensing the light environment in plants: Photoreceptors and early signaling steps. Curr. Opin. Neurobiol. 2015, 34, 46–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, I.; Pokorny, R.; Byrdin, M.; Hoang, N.; Ritz, T.; Brettel, K.; Essen, L.-O.; van der Horst, G.T.J.; Batschauer, A.; Ahmad, M. The cryptochromes: Blue light photoreceptors in plants and animals. Annu. Rev. Plant Biol. 2011, 62, 335–364. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, B.; Zhao, C.; Pepper, M.; Lin, C. The action mechanisms of plant cryptochromes. Trends Plant Sci. 2011, 16, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Christie, J.M. Phototropin blue-light receptors. Annu. Rev. Plant Biol. 2007, 58, 21–45. [Google Scholar] [CrossRef] [Green Version]
- Suetsugu, N.; Wada, M. Evolution of three LOV blue light receptor families in green plants and photosynthetic stramenopiles: Phototropin, ZTL/FKF1/LKP2 and aureochrome. Plant Cell Physiol. 2013, 54, 8–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C. Plant blue-light receptors. Trends Plant Sci. 2000, 5, 337–342. [Google Scholar] [CrossRef]
- Rizzini, L.; Favory, J.-J.; Cloix, C.; Faggionato, D.; O’Hara, A.; Kaiserli, E.; Baumeister, R.; Schäfer, E.; Nagy, F.; Jenkins, G.I.; et al. Perception of UV-B by the Arabidopsis UVR8 protein. Science 2011, 332, 103–106. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, G.I. The UV-B Photoreceptor UVR8: From structure to physiology. Plant Cell 2014, 26, 21–37. [Google Scholar] [CrossRef] [Green Version]
- Tilbrook, K.; Arongaus, A.B.; Binkert, M.; Heijde, M.; Yin, R.; Ulm, R. The UVR8 UV-B Photoreceptor: Perception, signaling and response. Arab. Book 2013, 11, e0164. [Google Scholar] [CrossRef] [Green Version]
- Aphalo, P.J.; Ballare, C.L. On the importance of information-acquiring systems in plant-plant interactions. Funct. Ecol. 1995, 9, 5–14. [Google Scholar] [CrossRef]
- Gilbert, I.R.; Jarvis, P.G.; Smith, H. Proximity signal and shade avoidance differences between early and late successional trees. Nature 2001, 411, 792–795. [Google Scholar] [CrossRef] [PubMed]
- Ballaré, C.L. Keeping up with the neighbours: Phytochrome sensing and other signalling mechanisms. Trends Plant Sci. 1999, 4, 97–102. [Google Scholar] [CrossRef]
- Cao, S.; Luo, X.; Xu, D.; Tian, X.; Song, J.; Xia, X.; Chu, C.; He, Z. Genetic architecture underlying light and temperature mediated flowering in Arabidopsis, rice, and temperate cereals. New Phytol. 2021, 230, 1731–1745. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, Y.-L.; Qiao, X.-R.; Wang, J.; Wang, L.-D.; Xu, C.-S.; Zhang, X. Phototropins function in high-intensity blue light-induced hypocotyl phototropism in Arabidopsis by altering cytosolic calcium. Plant Physiol. 2013, 162, 1539–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kami, C.; Allenbach, L.; Zourelidou, M.; Ljung, K.; Schütz, F.; Isono, E.; Watahiki, M.K.; Yamamoto, K.T.; Schwechheimer, C.; Fankhauser, C. Reduced phototropism in Pks mutants may be due to altered auxin-regulated gene expression or reduced lateral auxin transport. Plant J. 2014, 77, 393–403. [Google Scholar] [CrossRef] [Green Version]
- Christie, J.M.; Yang, H.; Richter, G.L.; Sullivan, S.; Thomson, C.E.; Lin, J.; Titapiwatanakun, B.; Ennis, M.; Kaiserli, E.; Lee, O.R.; et al. Phot1 inhibition of ABCB19 primes lateral auxin fluxes in the shoot apex required for phototropism. PLoS Biol. 2011, 9, e1001076. [Google Scholar] [CrossRef]
- Sgamma, T.; Pape, J.; Massiah, A.; Jackson, S. Selection of reference genes for diurnal and developmental time-course real-time PCR expression analyses in lettuce. Plant Methods 2016, 12, 21. [Google Scholar] [CrossRef] [Green Version]
- Cirilli, M.; Delfino, I.; Caboni, E.; Muleo, R. EpiHRMAssay, in tube and in silico combined approach for the scanning and epityping of heterogeneous DNA methylation. Biol. Methods Protoc. 2017, 2, bpw008. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Sgrò, C.M. Climate change and evolutionary adaptation. Nature 2011, 470, 479–485. [Google Scholar] [CrossRef]
- Cavallaro, V.; Tringali, S.; Patanè, C. Large-scale in vitro propagation of giant reed (Arundo donax L.), a promising biomass species. J. Hortic. Sci. Biotechnol. 2011, 86, 452–456. [Google Scholar] [CrossRef]
- Shukla, M.R.; Singh, A.S.; Piunno, K.; Saxena, P.K.; Jones, A.M.P. Application of 3D printing to prototype and develop novel plant tissue culture systems. Plant Methods 2017, 13, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miler, N.; Kulus, D.; Woźny, A.; Rymarz, D.; Hajzer, M.; Wierzbowski, K.; Nelke, R.; Szeffs, L. Application of wide-spectrum light-emitting diodes in micropropagation of popular ornamental plant species: A study on plant quality and cost reduction. Vitr. Cell. Dev. Biol. Plant 2019, 55, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Morini, S.; Muleo, R.; Jain, S.; Ishii, K. Micropropagation of woody trees and fruits. In Effect of Light Quality on Micropropagation of Woody Species; Jain, S.M., Ishii, K., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 3–35. [Google Scholar] [CrossRef]
- Bula, R.J.; Morrow, R.C.; Tibbitts, T.W.; Barta, D.J.; Ignatius, R.W.; Martin, T.S. Light-emitting diodes as a radiation source for plants. HortScience 1991, 26, 203–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seabrook, J.E.A. Light effects on the growth and morphogenesis of potato (Solanum tuberosum) in vitro: A review. Am. J. Potato Res. 2005, 82, 353–367. [Google Scholar] [CrossRef]
- Bantis, F.; Ouzounis, T.; Radoglou, K. Artificial LED lighting enhances growth characteristics and total phenolic content of Ocimum basilicum, but variably affects transplant success. Sci. Hortic. 2016, 198, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Yeh, N.; Chung, J.-P. High-brightness LEDs energy efficient lighting sources and their potential in indoor plant cultivation. Renew. Sustain. Energy Rev. 2009, 13, 2175–2180. [Google Scholar] [CrossRef]
- Dutta Gupta, S.; Jatothu, B. Fundamentals and applications of light-emitting diodes (LEDs) in vitro plant growth and morphogenesis. Plant Biotechnol. Rep. 2013, 7, 211–220. [Google Scholar] [CrossRef]
- Kim, S.-J.; Hahn, E.-J.; Heo, J.-W.; Paek, K.-Y. Effects of LEDs on net photosynthetic rate, growth and leaf stomata of chrysanthemum plantlets in vitro. Sci. Hortic. 2004, 101, 143–151. [Google Scholar] [CrossRef]
- Kasahara, M.; Kagawa, T.; Sato, Y.; Kiyosue, T.; Wada, M. Phototropins mediate blue and red light-induced chloroplast movements in Physcomitrella patens. Plant Physiol. 2004, 135, 1388–1397. [Google Scholar] [CrossRef] [Green Version]
- Dutta Gupta, S.; Agarwal, A. Influence of LED Lighting on in vitro plant regeneration and associated cellular redox balance. In Light Emitting Diodes for Agriculture; Dutta Gupta, S., Ed.; Springer: Singapore, 2017; pp. 273–303. ISBN 978-981-10-5806-6. [Google Scholar]
- Darko, E.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Philos. Trans. R. Soc. B-Biol. Sci. 2014, 369, 1640. [Google Scholar] [CrossRef]
- Gupta, S.D.; Agarwal, A. Artificial Lighting System for Plant Growth and Development: Chronological Advancement, Working Principles, and Comparative Assessment; Gupta, S.D., Ed.; Springer: Singapore, 2017; pp. 1–26. ISBN 978-981-10-5806-6. [Google Scholar]
- Martineau, V.; Lefsrud, M.; Naznin, M.T.; Kopsell, D.A. Comparison of light-emitting diode and high-pressure sodium light treatments for hydroponics growth of Boston lettuce. HortScience 2012, 47, 477–482. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.T.; Bertolucci, S.K.V.; da Cunha, S.H.B.; Lazzarini, L.E.S.; Tavares, M.C.; Pinto, J.E.B.P. Effect of light and natural ventilation systems on the growth parameters and carvacrol content in the in vitro cultures of Plectranthus amboinicus (Lour.) Spreng. Plant Cell Tissue Organ Cult. 2017, 129, 501–510. [Google Scholar] [CrossRef]
- Budiarto, K. Spectral quality affects morphogenesis on Anthurium plantlet during in vitro culture. AJAS 2010, 32, 234–240. [Google Scholar]
- Li, H.; Xu, Z.; Tang, C. Effect of light-emitting diodes on growth and morphogenesis of upland cotton (Gossypium hirsutum L.) Plantlets in vitro. Plant Cell Tissue Organ Cult. 2010, 103, 155–163. [Google Scholar] [CrossRef]
- Li, H.; Tang, C.; Xu, Z. The effects of different light qualities on rapeseed (Brassica napus L.) plantlet growth and morphogenesis in vitro. Sci. Hortic. 2013, 150, 117–124. [Google Scholar] [CrossRef]
- Do Nascimento Vieira, L.; de Freitas Fraga, H.P.; dos Anjos, K.G.; Puttkammer, C.C.; Scherer, R.F.; da Silva, D.A.; Guerra, M.P. Light-emitting diodes (LED) increase the stomata formation and chlorophyll content in Musa acuminata (AAA)‘Nanicão Corupá’in vitro plantlets. Theor. Exp. Plant Physiol. 2015, 27, 91–98. [Google Scholar] [CrossRef]
- Merkle, S.A.; Montello, P.M.; Xia, X.; Upchurch, B.L.; Smith, D.R. Light quality treatments enhance somatic seedling production in three southern pine species. Tree Physiol. 2006, 26, 187–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Yeung, E.C.; Paek, K.Y. Endoreduplication in Phalaenopsis is affected by light quality from light-emitting diodes during somatic embryogenesis. Plant Biotechnol. Rep. 2010, 4, 303–309. [Google Scholar] [CrossRef]
- Kim, Y.W.; Moon, H.K. Enhancement of somatic embryogenesis and plant regeneration in Japanese red pine (Pinus densiflora). Plant Biotechnol. Rep. 2014, 8, 259–266. [Google Scholar] [CrossRef]
- Botero Giraldo, C.; Urrea Trujillo, A.I.; Naranjo Gomez, E.J. Regeneration potential of Psychotria ipecacuanha (Rubiaceae) from thin cell layers. Acta Biol. Colomb. 2015, 20, 181–192. [Google Scholar] [CrossRef]
- Chen, C.-C.; Agrawal, D.C.; Lee, M.-R.; Lee, R.-J.; Kuo, C.-L.; Wu, C.-R.; Tsay, H.-S.; Chang, H.-C. Influence of LED light spectra on in vitro somatic embryogenesis and LC–MS analysis of chlorogenic acid and rutin in Peucedanum japonicum Thunb: A medicinal herb. Bot. Stud. 2016, 57, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mai, N.T.; Binh, P.T.; Khoi, P.H.; Hung, N.K.; Ngoc, P.B.; Ha, C.H.; Binh, H.T.T. Effects of light emitting diodes-LED on regeneration ability of Coffea canephora mediated via somatic embryogenesis. Acad. J. Biol. 2016, 38, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Lotfi, M.; Mars, M.; Werbrouck, S. Optimizing pear micropropagation and rooting with light emitting diodes and trans-cinnamic Acid. Plant Growth Regul. 2019, 88, 173–180. [Google Scholar] [CrossRef]
- Ding, Z.; Galván-Ampudia, C.S.; Demarsy, E.; Łangowski, Ł.; Kleine-Vehn, J.; Fan, Y.; Morita, M.T.; Tasaka, M.; Fankhauser, C.; Offringa, R.; et al. Light-mediated polarization of the PIN3 auxin transporter for the phototropic response in Arabidopsis. Nat. Cell Biol. 2011, 13, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Dewir, Y.; Chakrabarty, D.; Hahn, E.; Paek, K. The effects of paclobutrazol, light emitting diodes (LEDs) and sucrose on flowering of Euphorbia millii plantlets in vitro. Eur. J. Hortic. Sci. 2006, 71, 240. [Google Scholar]
- Moon, H.K.; Park, S.-Y.; Kim, Y.W.; Kim, C.S. Growth of Tsuru-rindo (Tripterospermum japonicum) cultured in vitro under various sources of light-emitting diode (LED) irradiation. J. Plant Biol. 2006, 49, 174–179. [Google Scholar] [CrossRef]
- Shin, K.S.; Murthy, H.N.; Heo, J.W.; Hahn, E.J.; Paek, K.Y. The effect of light quality on the growth and development of in vitro cultured Doritaenopsis plants. Acta Physiol. Plant. 2008, 30, 339–343. [Google Scholar] [CrossRef]
- Ma, X.; Wang, Y.; Liu, M.; Xu, J.; Xu, Z. Effects of green and red lights on the growth and morphogenesis of potato (Solanum tuberosum L.) plantlets in vitro. Sci. Hortic. 2015, 190, 104–109. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Halimah, N.; Ko, C.H.; Jeong, B.R. Blue LED light enhances growth, phytochemical contents, and antioxidant enzyme activities of Rehmannia glutinosa cultured in vitro. Hortic. Environ. Biotechnol. 2015, 56, 105–113. [Google Scholar] [CrossRef]
- Batista, D.S.; de Castro, K.M.; da Silva, A.R.; Teixeira, M.L.; Sales, T.A.; Soares, L.I.; das Graças Cardoso, M.; de Oliveira Santos, M.; Viccini, L.F.; Otoni, W.C. Light quality affects in vitro growth and essential oil profile in Lippia alba (Verbenaceae). Vitr. Cell. Dev. Biol. Plant 2016, 52, 276–282. [Google Scholar] [CrossRef]
- Seibert, M.; Wetherbee, P.J.; Job, D.D. The effects of light intensity and spectral quality on growth and shoot initiation in tobacco callus. Plant Physiol. 1975, 56, 130–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chée, R. In vitro culture of Vitis: The effects of light spectrum, manganese sulfate and potassium iodide on morphogenesis. Plant Cell Tissue Organ Cult. 1986, 7, 121–134. [Google Scholar] [CrossRef]
- Lercari, B.; Tognoni, F.; Anselmo, G.; Chapel, D. Photocontrol of in vitro bud differentiation in Saintpaulia ionantha leaves and lycopersicon esculentum cotyledons. Physiol. Plant. 1986, 67, 340–344. [Google Scholar] [CrossRef]
- Chee, R.; Pool, R. Morphogenic responses to propagule trimming, spectral irradiance, and photoperiod of grapevine shoots recultured in vitro. J. Am. Soc. Hortic. Sci. 1989, 114, 350–354. [Google Scholar]
- Aksenova, N.P.; Konstantinova, T.N.; Sergeeva, L.I.; Macháčková, I.; Golyanovskaya, S.A. Morphogenesis of potato plants in vitro. I. Effect of light quality and hormones. Plant Growth Regul. 1994, 13, 143–146. [Google Scholar] [CrossRef]
- Panizza, M.; Tognoni, F.; Lercari, B. Axillary bud proliferation and ethylene production as controlled by radiation of different spectral composition and exogenous phytohormones. Biol. Plant. 1994, 36, 553–563. [Google Scholar] [CrossRef]
- Castillo, B.; Smith, M.A.L. Direct somatic embryogenesis from Begonia gracilis explants. Plant Cell Rep. 1997, 16, 385–388. [Google Scholar] [CrossRef]
- Da Silva, M.; Debergh, P. The effect of light quality on the morphogenesis of in vitro cultures of Azorina vidalii (Wats.) Feer. Plant Cell Tissue Organ Cult. 1997, 51, 187–193. [Google Scholar] [CrossRef]
- Marks, T.; Simpson, S. Effect of irradiance on shoot development in vitro. Plant Growth Regul. 1999, 28, 133–142. [Google Scholar] [CrossRef]
- Lercari, B.; Moscatelli, S.; Ghirardi, E.; Niceforo, R.; Bertram, L. Photomorphogenic control of shoot regeneration from etiolated and light-grown hypocotyls of tomato. Plant Sci. 1999, 140, 53–62. [Google Scholar] [CrossRef]
- Michalczuk, B.; Michalczuk, L. The effect of light quality on regeneration rate and plantlet development in transgenic petunia ‘Revolution’(Surfinia type). Acta Hortic. 2000, 530, 397–402. [Google Scholar] [CrossRef]
- Lian, M.-L.; Murthy, H.; Paek, K.-Y. Effects of light emitting diodes (LEDs) on the in vitro induction and growth of bulblets of Lilium oriental hybrid ‘Pesaro’. Sci. Hortic. 2002, 94, 365–370. [Google Scholar] [CrossRef]
- Burritt, D.J.; Leung, D.W. Adventitious shoot regeneration from Begonia x erythrophylla petiole sections is developmentally sensitive to light quality. Physiol. Plant. 2003, 118, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Huan, L.V.T.; Tanaka, M. Effects of red and blue light-emitting diodes on callus induction, callus proliferation, and protocorm-like body formation from callus in Cymbidium orchid. Environ. Control Biol. 2004, 42, 57–64. [Google Scholar] [CrossRef]
- Hunter, D.C.; Burritt, D.J. Light quality influences adventitious shoot production from cotyledon explants of Lettuce (Lactuca sativa L.). In Vitro Cell. Dev. Biol. Plant 2004, 40, 215–220. [Google Scholar] [CrossRef]
- Yonghua, Q.; Shanglong, Z.; Asghar, S.; Lingxiao, Z.; Qiaoping, Q.; Kunsong, C.; Changjie, X. Regeneration mechanism of Toyonoka strawberry under different color plastic Films. Plant Sci. 2005, 168, 1425–1431. [Google Scholar] [CrossRef]
- Dewir, Y.H.; Chakrabarty, D.; Kim, S.J.; Hahn, E.J.; Paek, K.Y. Effect of light-emitting diode on growth and shoot proliferation of Euphorbia millii and Spathiphyllum cannifolium. Hortic. Environ. Biotechnol. 2005, 46, 375–379. [Google Scholar]
- Moshe, R.; Dalia, E. On the effect of light on shoot regeneration in petunia. Plant Cell Tissue Organ Cult. 2007, 89, 49–54. [Google Scholar]
- Poudel, P.R.; Kataoka, I.; Mochioka, R. Effect of red and blue-light-emitting diodes on growth and morphogenesis of grapes. Plant Cell Tissue Organ Cult. 2008, 92, 147–153. [Google Scholar] [CrossRef]
- Wongnok, A.; Piluek, C.; Techasilpitak, T.; Tantivivat, S. Effects of light emitting diodes on micropropagation of Phalaenopsis orchids. Acta Hortic. 2007, 788, 149–156. [Google Scholar] [CrossRef]
- Xu, Z.; Cui, J. Effects of different spectral energy distribution on tissue culture of Oncidium in vitro. J. Beijing For. Univ. 2009, 31, 45–50. [Google Scholar]
- Hamada, K.; Shimasaki, K.; Nishimura, Y.; Egawa, H.; Yoshida, K. Effect of red fluorescent films on the proliferation of Cymbidium finlaysonianum Lindl. PLB cultured in vitro. Hortic. Environ. Biotechnol. 2009, 50, 319–323. [Google Scholar]
- Chung, J.-P.; Huang, C.-Y.; Dai, T.-E. Spectral effects on embryogenesis and plantlet growth of Oncidium ‘Gower Ramsey’. Sci. Hortic. 2010, 124, 511–516. [Google Scholar] [CrossRef]
- Mengxi, L.; Zhigang, X.; Yang, Y.; Yijie, F. Effects of different spectral lights on Oncidium PLBs induction, proliferation, and plant regeneration. Plant Cell Tissue Organ Cult. 2011, 106, 1–10. [Google Scholar] [CrossRef]
- Hamada, K.; Shimasaki, K.; Nishimura, Y.; Sasaoka, H.; Nishimura, K. Effects of red, blue and yellow fluorescent films on proliferation and organogenesis in Cymbidium and Phalaenopsis PLB in vitro. Acta Hortic. 2011, 907, 385–388. [Google Scholar] [CrossRef]
- Lin, Y.; Li, J.; Li, B.; He, T.; Chun, Z. Effects of light quality on growth and development of protocorm-like bodies of dendrobium officinale in vitro. Plant Cell Tissue Organ Cult. 2011, 105, 329–335. [Google Scholar] [CrossRef]
- Nahar, S.; Shimasaki, K.; Haque, S. Effect of different light and two polysaccharides on the proliferation of protocorm-like bodies of Cymbidium cultured in vitro. Acta Hortic. 2012, 956, 307–313. [Google Scholar] [CrossRef]
- Werbrouck, S.; Buyle, H.; Geelen, D.; Van Labeke, M.C. Effect of red-, far-red- and blue-light-emitting diodes on in vitro growth of Ficus benjamina. Acta Hortic. 2012, 961, 533–538. [Google Scholar] [CrossRef] [Green Version]
- Kaewjampa, N.; Shimasaki, K. Effects of green LED lighting on organogenesis and superoxide dismutase (SOD) activities in protocorm-like bodies (PLBs) of Cymbidium cultured in vitro. Environ. Control Biol. 2012, 50, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Siegień, I.; Adamczuk, A.; Wróblewska, K. Light affects in vitro organogenesis of Linum usitatissimum L. and its cyanogenic potential. Acta Physiol. Plant. 2013, 35, 781–789. [Google Scholar] [CrossRef] [Green Version]
- Edesi, J.; Kotkas, K.; Pirttilä, A.M.; Häggman, H. Does light spectral quality affect survival and regeneration of potato (Solanum tuberosum L.) shoot tips after cryopreservation? Plant Cell Tissue Organ Cult. 2014, 119, 599–607. [Google Scholar] [CrossRef]
- Lee, N.N.; Choi, Y.E.; Moon, H.K. Effect of LEDs on shoot multiplication and rooting of rare plant Abeliophyllum distichum Nakai. J. Plant Biochem. Biotechnol. 2014, 41, 94–99. [Google Scholar] [CrossRef]
- Habiba, S.U.; Shimasaki, K.; Ahasan, M.M.; Alam, M.M. Effects of different light quality on growth and development of protocorm-like bodies (PLBs) in Dendrobium kingianum cultured in vitro. Bangladesh Res. Public. J. 2014, 10, 223–227. [Google Scholar]
- Kamal, M.M.; Shimasaki, K.; Akter, N. Effect of light emutting diode (LED) lamps and N-Acetylglucosamine (NAG) on organogenesis in protocorm-like bodies (PLBs) of a Cymbidium hybrid cultured in vitro. Plant Tissue Cult. Biotechnol. 2014, 24, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.M.A.; de Oliveira, A.L.B.; Oliveira-Filho, R.A.; Gouveia-Neto, A.S.; Camara, T.J.R.; Willadino, L.G. Effect of blue/red LED light combination on growth and morphogenesis of Saccharum officinarum plantlets in vitro. In Proceedings of the SPIE—The International Society for Optical Engineering, San Francisco, CA, USA, 4 March 2014; Volume 8947. [Google Scholar] [CrossRef]
- Jeong, B.R.; Sivanesan, I. Adventitious shoot regeneration, in vitro flowering, fruiting, secondary metabolite content and antioxidant activity of Scrophularia takesimensis Nakai. Plant Cell Tissue Organ Cult. 2015, 123, 607–618. [Google Scholar] [CrossRef]
- Dutta Gupta, S.; Sahoo, T. Light emitting diode (LED)-induced alteration of oxidative events during in vitro shoot organogenesis of Curculigo orchioides Gaertn. Acta Physiol. Plant. 2015, 37, 233. [Google Scholar] [CrossRef]
- Hung, C.D.; Hong, C.-H.; Jung, H.-B.; Kim, S.-K.; Van Ket, N.; Nam, M.-W.; Choi, D.-H.; Lee, H.-I. Growth and morphogenesis of encapsulated strawberry shoot tips under mixed LEDs. Sci. Hortic. 2015, 194, 194–200. [Google Scholar] [CrossRef]
- Nhut, D.T.; Huy, N.P.; Tai, N.T.; Nam, N.B.; Luan, V.Q.; Hien, V.T.; Tung, H.T.; Vinh, B.T.; Luan, T.C. Light-emitting diodes and their potential in callus growth, plantlet development and saponin accumulation during somatic embryogenesis of Panax vietnamensis Ha et Grushv. Biotechnol. Biotechnol. Equip. 2015, 29, 299–308. [Google Scholar] [CrossRef]
- Bello-Bello, J.J.; Martínez-Estrada, E.; Caamal-Velázquez, J.H.; Morales-Ramos, V. Effect of LED light quality on in vitro shoot proliferation and growth of Vanilla (Vanilla planifolia Andrews). Afr. J. Biotechnol. 2016, 15, 272–277. [Google Scholar]
- Gök, K.M.; Şan, B.; Bayhan, A.K. Micropropagation of Gerbera (Gerbera jamesonii bolus) under different color of light-emitting diodes. Süleyman Demirel Üniversitesi Fen Bilimleri Enstitüsü Dergis 2016, 20, 468. [Google Scholar] [CrossRef] [Green Version]
- Nahar, S.J.; Haque, S.M.; Kazuhiko, S. Application of chondroitin sulfate on organogenesis of two Cymbidium spp. under different sources of lights. Not. Sci. Biol. 2016, 8, 156–160. [Google Scholar] [CrossRef] [Green Version]
- Karataş, M.; Aasim, M.; Dazkirli, M. Influence of light-emitting diodes and benzylaminopurin on adventitious shoot regeneration of water hyssop (Bacopa monnieri (L.) Pennell) in vitro. Arch. Biol. Sci. 2016, 68, 501–508. [Google Scholar] [CrossRef]
- Hung, C.D.; Hong, C.-H.; Kim, S.-K.; Lee, K.-H.; Park, J.-Y.; Dung, C.D.; Nam, M.-W.; Choi, D.-H.; Lee, H.-I. In vitro proliferation and ex vitro rooting of microshoots of commercially important rabbiteye blueberry (Vaccinium ashei reade) Using spectral lights. Sci. Hortic. 2016, 211, 248–254. [Google Scholar] [CrossRef]
- Martínez-Estrada, E.; Caamal-Velázquez, J.H.; Morales-Ramos, V.; Bello-Bello, J. Light emitting diodes improve in vitro shoot multiplication and growth of Anthurium andreanum Lind. Propag. Ornam. Plants 2016, 16, 3–8. [Google Scholar]
- Silva, M.M.; de Oliveira, A.L.B.; Oliveira-Filho, R.A.; Camara, T.; Willadino, L.; Gouveia-Neto, A. The effect of spectral light quality on in vitro culture of sugarcane. Acta Sci. Biol. Sci. 2016, 38, 157–161. [Google Scholar] [CrossRef] [Green Version]
- Szewczyk-Taranek, B.; Pawłowska, B.; Prokopiuk, B.; Żupnik, M. Effectiveness of LED and fluorescent light on in vitro shoot proliferation of Staphylea pinnata. Acta Hortic. 2015, 1155, 375–380. [Google Scholar]
- Ramírez-Mosqueda, M.A.; Iglesias-Andreu, L.G.; Bautista-Aguilar, J.R. The effect of light quality on growth and development of in vitro plantlet of Stevia rebaudiana Bertoni. Sugar Tech. 2017, 19, 331–336. [Google Scholar] [CrossRef]
- Ramírez-Mosqueda, M.; Iglesias-Andreu, L.; Luna-Sánchez, I. Light quality affects growth and development of in vitro plantlet of Vanilla planifolia Jacks. S. Afr. J. Bot. 2017, 109, 288–293. [Google Scholar] [CrossRef]
- Billore, V.; Jain, M.; Suprasanna, P. Monochromic radiation through light-emitting diode (LED) positively augments in vitro shoot regeneration in Orchid (Dendrobium sonia). Can. J. Biotech. 2017, 1, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, L.T.; de Araújo Silva, M.M.; Ulisses, C.; Camara, T.R.; Willadino, L. Using LED lighting in somatic embryogenesis and micropropagation of an elite sugarcane variety and its effect on redox metabolism during acclimatization. Plant Cell Tissue Organ Cult. 2017, 128, 211–221. [Google Scholar] [CrossRef]
- Pawłowska, B.; Żupnik, M.; Szewczyk-Taranek, B.; Cioć, M. Impact of LED light sources on morphogenesis and levels of photosynthetic pigments in Gerbera jamesonii grown in vitro. Hortic. Environ. Biotechnol. 2018, 59, 115–123. [Google Scholar] [CrossRef]
- Lazzarini, L.E.S.; Bertolucci, S.K.V.; Pacheco, F.V.; dos Santos, J.; Silva, S.T.; de Carvalho, A.A.; Pinto, J.E.B.P. Quality and intensity of light affect Lippia gracilis Schauer plant growth and volatile compounds in vitro. Plant Cell Tissue Organ Cult. 2018, 135, 367–379. [Google Scholar] [CrossRef]
- Cioć, M.; Szewczyk, A.; Żupnik, M.; Kalisz, A.; Pawłowska, B. LED lighting affects plant growth, morphogenesis and phytochemical contents of Myrtus communis L. in vitro. Plant Cell Tissue Organ Cult. 2018, 132, 433–447. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Song, C.; Sun, L.; Li, L.; Xu, Z.; Tang, C. Effects of light-emitting diodes on tissue culture plantlets and seedlings of Rice (Oryza sativa L.). J. Integr. Agric. 2020, 19, 1743–1754. [Google Scholar] [CrossRef]
- Kadkade, P.; Jopson, H. Influence of light quality on organogenesis from the embryo-derived callus of Douglas fir (Pseudotsuga menziesii). Plant Sci. Lett. 1978, 13, 67–73. [Google Scholar] [CrossRef]
- Norton, C.R.; Herrington, T.; Phillips, D.; Norton, M. Light quality and light pipe in the micropropagation of woody ornamental plants grown in vitro. Acta Hort. 1988, 227, 413–416. [Google Scholar] [CrossRef]
- Herrington, E.; McPherson, J.C. Light quality growth promotion of Spiraea nipponica: The influence of a low photon fluence rate and transfer time to a higher fluence rate. Plant Cell Tissue Organ Cult. 1993, 32, 161–167. [Google Scholar] [CrossRef]
- Muleo, R.; Thomas, B. Effects of light quality on shoot proliferation of Prunus cerasifera in vitro are the result of differential effects on bud induction and apical dominance. J. Hortic. Sci. 1997, 72, 483–499. [Google Scholar] [CrossRef]
- D’Onofrio, C.; Morini, S.; Bellocchi, G. Effect of light quality on somatic embryogenesis of quince leaves. Plant Cell Tissue Organ Cult. 1998, 53, 91–98. [Google Scholar] [CrossRef]
- Piagnani, C.; Iacona, C.; Intrieri, M.C.; Muleo, R. A New somaclone of Prunus avium shows diverse growth pattern under different spectral quality of radiation. Biologia Plant. 2002, 45, 11–17. [Google Scholar] [CrossRef]
- Muleo, R.; Morini, S. Light quality regulates shoot cluster growth and development of MM106 apple genotype in vitro culture. Sci. Hortic. 2006, 108, 364–370. [Google Scholar] [CrossRef]
- Wang, H.; Liu, H.; Wang, W.-J.; Zu, Y. Effects of thidiazuron, basal medium and light quality on adventitious shoot regeneration from in vitro cultured stem of Populus alba × P. berolinensis. J. For. Res. 2008, 19, 257–259. [Google Scholar] [CrossRef]
- Wilken, D.; Jiménez Gonzalez, E.; Gerth, A.; Gómez-Kosky, R.; Schumann, A.; Claus, D. Effect of immersion systems, lighting, and TIS designs on biomass increase in micropropagating banana (Musa Spp. Cv.’Grande Naine’AAA). Vitr. Cell. Dev. Biol. Plant. 2014, 50, 582–589. [Google Scholar] [CrossRef]
- Kwon, A.; Cui, H.-Y.; Lee, H.; Shin, H.; Kang, K.-S.; Park, S.-Y. Light quality affects shoot regeneration, cell division, and wood formation in elite clones of Populus euramericana. Acta Physiol. Plant. 2015, 37, 65. [Google Scholar] [CrossRef]
- Geng, F.; Moran, R.; Day, M.; Halteman, W.; Zhang, D. In vitro shoot proliferation of apple rootstocks ‘B. 9’, ‘G. 30’, and ‘G. 41’grown under red and blue light. HortScience 2015, 50, 430–433. [Google Scholar] [CrossRef]
- Al-Mayahi, A.M.W. Effect of red and blue light emitting diodes “CRB-LED” on in vitro organogenesis of date palm (Phoenix dactylifera L.) cv. Alshakr. World J. Microbiol. Biotechnol. 2016, 32, 160. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Zeng, Y.; Fu, Y.; Wu, J.; Liang, Q. Light quality affects the proliferation of in vitro cultured plantlets of Camellia oleifera Huajin. PeerJ 2020, 8, e10016. [Google Scholar] [CrossRef] [PubMed]
- Muleo, R.; Morini, S. Physiological dissection of blue and red light regulation of apical dominance and branching in M9 apple rootstock growing in vitro. J. Plant Physiol. 2008, 165, 1838–1846. [Google Scholar] [CrossRef] [PubMed]
- Gabryszewska, E.; Rudnicki, R. The influence of light quality on the shoot proliferation and rooting of Gerbera jamesonii in vitro. Acta Agrobot. 1995, 48, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Kadkade, P.; Seibert, M. Phytochrome-regulated organogenesis in lettuce tissue culture. Nature 1977, 270, 49–50. [Google Scholar] [CrossRef]
- Cybularz-Urban, T.; Hanus-Fajerska, E.; Swiderski, A. Effect of light wavelength on in vitro organogenesis of a Cattleya hybrid. Acta Biol. Crac. Ser. Bot. 2007, 49, 113–118. [Google Scholar]
- Morelli, G.; Ruberti, I. Shade avoidance responses. Driving auxin along lateral routes. Plant Physiol. 2000, 122, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Finlayson, S.A.; Krishnareddy, S.R.; Kebrom, T.H.; Casal, J.J. Phytochrome regulation of branching in Arabidopsis. Plant Physiol. 2010, 152, 1914–1927. [Google Scholar] [CrossRef] [Green Version]
- Baraldi, R.; Rossi, F.; Lercari, B. In vitro shoot development of Prunus GF 655–2: Interaction between light and benzyladenine. Physiol. Plant. 1988, 74, 440–443. [Google Scholar] [CrossRef]
- Muleo, R.; Morini, S.; Casano, S. Photoregulation of growth and branching of plum shoots: Physiological action of two photosystems. Vitr. Cell. Dev. Biol. Plant 2001, 37, 609–617. [Google Scholar] [CrossRef]
- Schuerger, A. Anatomical features of pepper plants (Capsicum annuum L.) grown under red light-emitting diodes supplemented with blue or far-red light. Ann. Bot. 1997, 79, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Appelgren, M. Effects of light quality on stem elongation of Pelargonium in vitro. Sci. Hortic. 1991, 45, 345–351. [Google Scholar] [CrossRef]
- Heo, J.W.; Shin, K.S.; Kim, S.K.; Paek, K.Y. Light quality affects in vitro growth of grape Teleki 5BB. J. Plant Biol. 2006, 49, 276–280. [Google Scholar] [CrossRef]
- Hahn, E.-J.; Kozai, T.; Paek, K.-Y. Blue and red light-emitting diodes with or without sucrose and ventilation affect in vitro growth of Rehmannia glutinosa plantlets. J. Plant Biol. 2000, 43, 247–250. [Google Scholar] [CrossRef]
- Hung, C.D.; Hong, C.-H.; Kim, S.-K.; Lee, K.-H.; Park, J.-Y.; Nam, M.-W.; Choi, D.-H.; Lee, H.-I. LED Light for in vitro and ex vitro efficient growth of economically important highbush blueberry (Vaccinium corymbosum L.). Acta Physiol. Plant. 2016, 38, 152. [Google Scholar] [CrossRef]
- Tanaka, M.; Takamura, T.; Watanabe, H.; Endo, M.; Yanagi, T.; Okamoto, K. In vitro growth of cymbidium plantlets cultured under superbright red and blue light-emitting diodes (LEDs). J. Hortic. Sci. Biotechnol. 1998, 73, 39–44. [Google Scholar] [CrossRef]
- Nhut, D.T.; Takamura, T.; Watanabe, H.; Okamoto, K.; Tanaka, M. Responses of strawberry plantlets cultured in vitro under superbright red and blue light-emitting diodes (LEDs). Plant Cell Tissue Organ Cult. 2003, 73, 43–52. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, M.J. Development of zygotic embryos and seedlings is affected by radiationspectral compositions from light emitting diode (LED) system in Chestnut (Castanea crenata S. et Z.). J. Korean For. Soc. 2010, 99, 750–754. [Google Scholar]
- Chen, Y.-C.; Chang, C.; Lin, H.-L. Topolins and red light improve the micropropagation efficiency of passion fruit (Passiflora edulis Sims) ‘Tainung No. 1’. Hortscience 2020, 55, 1337–1344. [Google Scholar] [CrossRef]
- Wu, H.-C.; Lin, C.-C. Red light-emitting diode light irradiation improves root and leaf formation in difficult to propagate Protea cynaroides L. plantlets in vitro. HortScience 2012, 47, 1490–1494. [Google Scholar] [CrossRef] [Green Version]
- Tibbitts, T.; Morgan, D.; Warrington, I. Growth of lettuce, spinach, mustard, and wheat plants under four combinations of high-pressure sodium, metal halide, and tungsten halogen lamps at equal PPFD. J. Am. Soc. Hort. Sci. 1983, 108, 622–630. [Google Scholar]
- Sæbø, A.; Krekling, T.; Appelgren, M. Light quality affects photosynthesis and leaf anatomy of birch plantlets in vitro. Plant Cell Tissue Organ Cult. 1995, 41, 177–185. [Google Scholar] [CrossRef]
- Maluta, F.A.; Bordignon, S.R.; Rossi, M.L.; Ambrosano, G.M.B.; Rodrigues, P.H.V. In vitro culture of sugarcane exposed to different light sources. Pesqui. Agropecu. Bras. 2013, 48, 1303–1307. [Google Scholar] [CrossRef] [Green Version]
- Miao, Y.; Chen, Q.; Qu, M.; Gao, L.; Hou, L. Blue light alleviates ‘red light syndrome’ by regulating chloroplast ultrastructure, photosynthetic traits and nutrient accumulation in cucumber plants. Sci. Hortic. 2019, 257, 108680. [Google Scholar] [CrossRef]
- Kowallik, W. Blue light effects on respiration. Annu. Rev. Plant Physiol. 1982, 33, 51–72. [Google Scholar] [CrossRef]
- Liu, M.; Xu, Z.; Guo, S.; Tang, C.; Liu, X.; Jao, X. Evaluation of leaf morphology, structure and biochemical substance of balloon flower (Platycodon grandiflorum (Jacq.) A. DC.) plantlets in vitro under different light spectra. Sci. Hortic. 2014, 174, 112–118. [Google Scholar] [CrossRef]
- Weis, J.; Jaffe, M. Photoenhancement by blue light of organogenesis in tobacco Pith Cultures. Physiol. Plant. 1969, 22, 171–176. [Google Scholar] [CrossRef]
- Bach, A.; Świderski, A. The effect of light quality on organogenesis of Hyacinthus orientalis L. in vitro. Acta Biol. Crac. Ser. Bot. 2000, 42, 115–120. [Google Scholar]
- Cybularz-Urban, T.; Hanus-Fajerska, E.; Bach, A. Callus induction and organogenesis in vitro of Cattleya from protocorm-like bodies (PLBs) under different light conditions. Acta Sci. Pol.-Hortorum Cultus 2015, 14, 19–28. [Google Scholar]
- Latkowska, M.; Kvaalen, H.; Appelgren, M. Genotype dependent blue and red light inhibition of the proliferation of the embryogenic tissue of Norway spruce. In Vitro Cell. Dev. Biol. Plant 2000, 36, 57–60. [Google Scholar] [CrossRef]
- Muleo, R.; Diodati, A.; Sgamma, T.M.; Morini, S. Ruolo del fotoperiodo e della qualità della luce nella micropropagazione delle specie arboree da frutto. Italus Hortus 2009, 16, 221–225. [Google Scholar]
- Karpinski, S.; Szechynska-Habda, M. Evidence for light wavelength-specific systemic photoelectrical signalling and cellular light memory in Arabidopsis thaliana. Plant Cell 2010, 22, 1–18. [Google Scholar]
- Manivannan, A.; Soundararajan, P.; Park, Y.G.; Wei, H.; Kim, S.H.; Jeong, B.R. Blue and red light-emitting diodes improve the growth and physiology of in vitro grown carnations ‘green beauty’ and ‘purple beauty’. Hortic. Environ. Biotechnol. 2017, 58, 12–20. [Google Scholar] [CrossRef]
- Toscano, S.; Cavallaro, V.; Ferrante, A.; Romano, D.; Patané, C. Effects of different light spectra on final biomass production and nutritional quality of two microgreens. Plants 2021, 10, 1584. [Google Scholar] [CrossRef]
- Sgamma, T.; Forgione, I.; Luziatelli, F.; Iacona, C.; Mancinelli, R.; Thomas, B.; Ruzzi, M.; Muleo, R. Monochromic radiations provided by light emitted diode (LED) modulate infection and defense response to fire blight in pear trees. Plants 2021, 10, 1886. [Google Scholar] [CrossRef]
- Cirvilleri, G.; Spina, S.; Iacona, C.; Catara, A.; Muleo, R. Study of rhizosphere and phyllosphere bacterial community and resistance to bacterial canker in genetically engineered phytochrome a cherry plants. J. Plant Physiol. 2008, 165, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Manivannan, A.; Soundararajan, P.; Park, Y.G.; Jeong, B.R. Physiological and proteomic insights into red and blue light-mediated enhancement of in vitro growth in Scrophularia kakudensis a potential medicinal plant. Front. Plant Sci. 2021, 11, 607007. [Google Scholar] [CrossRef] [PubMed]
- Iacona, C.; Muleo, R. Light quality affects in vitro adventitious rooting and ex vitro performance of cherry rootstock colt. Sci. Hortic. 2010, 125, 630–636. [Google Scholar] [CrossRef]
- Jao, R.-C.; Lai, C.-C.; Fang, W.; Chang, S.-F. Effects of red light on the growth of Zantedeschia plantlets in vitro and tuber formation using light-emitting diodes. HortScience 2005, 40, 436–438. [Google Scholar] [CrossRef] [Green Version]
- Alvarenga, I.C.A.; Pacheco, F.V.; Silva, S.T.; Bertolucci, S.K.V.; Pinto, J.E.B.P. In vitro culture of Achillea millefolium L.: Quality and intensity of light on growth and production of volatiles. Plant Cell Tissue Organ Cult. 2015, 122, 299–308. [Google Scholar] [CrossRef]
- Nhut, D.; Takamura, T.; Watanabe, H.; Tanaka, M. Efficiency of a novel culture system by using light-emitting diode (led) on in vitro and subsequent growth of micropropagated banana plantlets. Acta Hortic. 2003, 616, 121–127. [Google Scholar] [CrossRef]
- Kurilčik, A.; Miklušytė-Čanova, R.; Dapkūnienė, S.; Žilinskaitė, S.; Kurilčik, G.; Tamulaitis, G.; Duchovskis, P.; Žukauskas, A. In vitro culture of Chrysanthemum plantlets using light-emitting diodes. Open Life Sci. 2008, 3, 161–167. [Google Scholar] [CrossRef]
- Macedo, A.F.; Leal-Costa, M.V.; Tavares, E.S.; Lage, C.L.S.; Esquibel, M.A. The effect of light quality on leaf production and development of in vitro cultured plants of Alternanthera brasiliana Kuntze. Environ. Exp. Bot. 2011, 70, 43–50. [Google Scholar] [CrossRef]
- Richter, G.; Wessel, K. Red light inhibits blue light-induced chloroplast development in cultured plant cells at the mRNA level. Plant Mol. Biol. 1985, 5, 175–182. [Google Scholar] [CrossRef]
- Bello-Bello, J.J.; Pérez-Sato, J.A.; Cruz-Cruz, C.A.; Martínez-Estrada, E. Light-Emitting Diodes: Progress in Plant Micropropagation. In Chlorophyll; Jacob-Lopes, E., Zepka, L.Q., Queiroz, M.I., Eds.; InTech: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Larcher, W. Plant Ecophysiology (Ecofisiologia Vegetal); RIMA Artes e Textos (Por): São Carlos, Brazil, 2006; pp. 525–550. [Google Scholar]
- Sivakumar, G.; Heo, J.; Kozai, T.; Paek, K. Effect of continuous or intermittent radiation on sweet potato plantlets in vitro. J. Hortic. Sci. Biotechnol. 2006, 81, 546–548. [Google Scholar] [CrossRef]
- Nhut, D.T.; Hong, L.; Watanabe, H.; Goi, M.; Tanaka, M. Growth of banana plantlets cultured in vitro under red and blue light-emitting diode (LED) irradiation source. Acta Hortic. 2002, 575, 117–124. [Google Scholar] [CrossRef]
- Nhut, D.; Takamura, T.; Watanabe, H.; Murakami, A.; Murakami, K.; Tanaka, M. Sugar-free micropropagation of Eucalyptus citriodora using light-emitting diodes (LEDs) and film-rockwool culture system. Environ. Control Biol. 2002, 40, 147–155. [Google Scholar]
- Lee, S.-H.; Tewari, R.K.; Hahn, E.-J.; Paek, K.-Y. Photon flux density and light quality induce changes in growth, stomatal development, photosynthesis and transpiration of Withania somnifera (L.) Dunal. Plantlets. Plant Cell Tissue Organ Cult. 2007, 90, 141–151. [Google Scholar] [CrossRef]
- Jao, R.-C.; Fang, W. Effects of frequency and duty ratio on the growth of potato plantlets in vitro using light-emitting diodes. HortScience 2004, 39, 375–379. [Google Scholar] [CrossRef] [Green Version]
- Baque, M.A.; Shin, Y.-K.; Elshmari, T.; Lee, E.-J.; Paek, K.-Y. Effect of light quality, sucrose and coconut water concentration on the microporpagation of Calanthe hybrids (‘Bukduseong’ × ‘Hyesung’ and ‘Chunkwang’ × ‘Hyesung’). Aust. J. Crop Sci. 2011, 5, 1247–1254. [Google Scholar]
- Cavallaro, V.; Avola, G.; Fascella, G.; Pellegrino, A.; La Rosa, S.; Di Silvestro, I.; Ierna, A. Effetti della qualità della luce indotta da lampade a LED sull’accrescimento e la moltiplicazione in vitro di specie ornamentali diverse per l’ambiente Mediterraneo. Acta Italus Hortus 2018, 21, 83–99. [Google Scholar]
- Andrade, H.; Braga, A.; Bertolucci, S.; Hsie, B.; Silva, S.; Pinto, J. Effect of plant growth regulators, light intensity and LED on growth and volatile compound of Hyptis suaveolens (L.) Poit in vitro plantlets. Acta Hortic. 2015, 1155, 277–284. [Google Scholar] [CrossRef]
- Brodersen, C.R.; Vogelmann, T.C. Do changes in light direction affect absorption profiles in leaves? Funct. Plant Biol. 2010, 37, 403–412. [Google Scholar] [CrossRef]
- Smith, H.L.; McAusland, L.; Murchie, E.H. Don’t ignore the green light: Exploring diverse roles in plant processes. J. Exp. Bot. 2017, 68, 2099–2110. [Google Scholar] [CrossRef]
- Golovatskaya, I.; Karnachuk, R.A. Role of green light in physiological activity of plants. Russ. J. Plant Physiol. 2015, 62, 727–740. [Google Scholar] [CrossRef]
- Folta, K.M.; Maruhnich, S.A. Green light: A signal to slow down or stop. J. Exp. Bot. 2007, 58, 3099–3111. [Google Scholar] [CrossRef]
- Liu, B.; Yang, Z.; Gomez, A.; Liu, B.; Lin, C.; Oka, Y. Signaling mechanisms of plant cryptochromes in Arabidopsis thaliana. J. Plant Res. 2016, 129, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Tanada, T. The photoreceptors in the high irradiance response of plants. Physiol. Plant. 1997, 101, 451–454. [Google Scholar] [CrossRef]
- Wang, Y.; Folta, K.M. Contributions of green light to plant growth and development. Am. J. Bot. 2013, 100, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Nishio, J.N.; Vogelmann, T.C. Green light drives CO2 fixation deep within leaves. Plant Cell Physiol. 1998, 39, 1020–1026. [Google Scholar] [CrossRef]
- Kim, H.-H.; Goins, G.D.; Wheeler, R.M.; Sager, J.C. Green-light supplementation for enhanced lettuce growth under red-and blue-light-emitting diodes. HortScience 2004, 39, 1617–1622. [Google Scholar] [CrossRef] [Green Version]
- Folta, K.M. Green light stimulates early stem elongation, antagonizing light-mediated growth inhibition. Plant Physiol. 2004, 135, 1407–1416. [Google Scholar] [CrossRef] [Green Version]
- Johkan, M.; Shoji, K.; Goto, F.; Hahida, S.-N.; Yoshihara, T. Effect of green light wavelength and intensity on photomorphogenesis and photosynthesis in Lactuca sativa. Environ. Exp. Bot. 2012, 75, 128–133. [Google Scholar] [CrossRef]
- Frechilla, S.; Talbott, L.D.; Bogomolni, R.A.; Zeiger, E. Reversal of blue light-stimulated stomatal opening by green light. Plant Cell Physiol. 2000, 41, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Loreti, F.; Muleo, R.; Morini, S. Effect of light quality on growth of in vitro cultured organs and tissues. Proc. Inter. Plant Prop. Soc. 1990, 40, 615–623. [Google Scholar]
- Zhang, Z.; Li, S.; Li, W. Effects of different light qualities on the multiplication of the callus from Vitis vinifera L. and resveratrol content. Plant Physiol. Commun. 2008, 44, 106. [Google Scholar]
- Dougher, T.A.; Bugbee, B. Evidence for yellow light suppression of lettuce growth. Photochem. Photobiol. 2001, 73, 208–212. [Google Scholar] [CrossRef]
- Casal, J.J. Photoreceptor signaling networks in plant responses to shade. Annu. Rev. Plant Biol. 2013, 64, 403–427. [Google Scholar] [CrossRef] [PubMed]
- Fraser, D.P.; Hayes, S.; Franklin, K.A. Photoreceptor crosstalk in shade avoidance. Curr. Opin. Plant Biol. 2016, 33, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Zhen, S.; van Iersel, M.W. Far-red light is needed for efficient photochemistry and photosynthesis. J. Plant Physiol. 2017, 209, 115–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, D. Effects of far-red light on the hormonal control of side shoot growth in the tomato. Ann. Bot. 1976, 40, 1033–1042. [Google Scholar] [CrossRef]
- Noe, N.; Eccher, T.; Del Signore, E.; Montoldi, A. Growth and proliferation in vitro of Vaccinium corymbosum under different irradiance and radiation spectral composition. Biol. Plant. 1998, 41, 161–167. [Google Scholar] [CrossRef]
- Pham, V.N.; Kathare, P.K.; Huq, E. Phytochromes and phytochrome interacting factors. Plant Physiol. 2018, 176, 1025–1038. [Google Scholar] [CrossRef] [Green Version]
- Debergh, P.; Aitken-Christie, J.; Cohen, D.; Grout, B.; Von Arnold, S.; Zimmerman, R.; Ziv, M. Reconsideration of the term ‘vitrification’ as used in micropropagation. Plant Cell Tissue Organ Cult. 1992, 30, 135–140. [Google Scholar] [CrossRef]
- Gago, J.; Martínez-Núñez, L.; Landín, M.; Flexas, J.; Gallego, P.P. Modeling the effects of light and sucrose on in vitro propagated plants: A multiscale system analysis using artificial intelligence technology. PLoS ONE 2014, 9, e85989. [Google Scholar] [CrossRef]
- Semorádová, Š.; Synková, H.; Pospíšilová, J. Responses of tobacco plantlets to change of irradiance during transfer from in vitro to ex vitro conditions. Photosynthetica 2002, 40, 605–614. [Google Scholar] [CrossRef]
- Kadleček, P.; Rank, B.; Tichá, I. Photosynthesis and photoprotection in Nicotiana tabacum L. in vitro grown plantlets. J. Plant Physiol. 2003, 160, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Patel, R. Factors influencing in vitro growth and shoot multiplication of pomegranate. Bioscan 2014, 9, 1031–1035. [Google Scholar]
- Taiz, L.; Zeiger, E. Fisiologia Vegetal; Universitat Jaume I: Castellón, Spain, 2006; Volume 10. [Google Scholar]
- Singh, A.S.; Jones, A.M.P.; Shukla, M.R.; Saxena, P.K. High Light intensity stress as the limiting factor in micropropagation of sugar maple (Acer saccharum Marsh.). Plant Cell Tissue Organ Cult. 2017, 129, 209–221. [Google Scholar] [CrossRef]
- Noè, N.; Eccher, T. Influence of irradiance on in vitro growth and proliferation of Vaccinium corymbosum (highbush blueberry) and subsequent rooting in vivo. Physiol. Plant. 1994, 91, 273–275. [Google Scholar] [CrossRef]
- Nhut, D.; Takamura, T.; Watanabe, H.; Tanaka, M. Artificial light source using light-emitting diodes (LEDs) in the efficient micropropagation of Spathiphyllum plantlets. Acta Hortic. 2005, 692, 137–142. [Google Scholar] [CrossRef]
- Soontornchainaksaeng, P.; Chaicharoen, S.; Sirijuntarut, M.; Kruatrachue, M. In vitro studies on the effect of light intensity on plant growth of Phaius tankervilliae (Banks Ex L’Herit.) Bl. and Vanda coerulea Griff. Sci. Asia 2001, 27, 233–237. [Google Scholar] [CrossRef]
- Wang, Q. The effect of light, darkness and temperature on micropropagation of the pear rootstock BP10030. J. Hort. Sci. 1992, 67, 869–876. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, D.; Ma, Z.; Li, X.; Xiao, Y. Growth and photosynthetic capability of Momordica grosvenori plantlets grown photoautotrophically in response to light intensity. HortScience 2009, 44, 757–763. [Google Scholar] [CrossRef]
- Infante, R.; Magnanini, E.; Righetti, B. The role of light and CO2 in optimising the conditions for shoot proliferation of Actinidia deliciosa in vitro. Physiol. Plant. 1989, 77, 191–195. [Google Scholar] [CrossRef]
- Bressan, P.; Kim, Y.; Hyndman, S.; Hasegawa, P.; Bressan, R. Factors affecting in vitro propagation of rose. J. Am. Soc. Hort. Sci. 1982, 107, 979–990. [Google Scholar]
- Jo, U.; Murthy, H.; Hahn, E.; Paek, K. Micropropagation of Alocasia amazonica using semisolid and liquid cultures. In Vitro Cell. Dev. Biol. Plant 2008, 44, 26–32. [Google Scholar] [CrossRef]
- Biswal, A.K.; Pattanayak, G.K.; Pandey, S.S.; Leelavathi, S.; Reddy, V.S.; Govindjee; Tripathy, B.C. Light intensity-dependent modulation of chlorophyll b biosynthesis and photosynthesis by overexpression of chlorophyllide a oxygenase in Tobacco. Plant Physiol. 2012, 159, 433–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Guan, Q.; Wei, Y.; Zhang, Z. Effects of Sucrose Concentration and light intensity on growth and photosynthesis of ginger plantlets in vitro. Chin. J. Appl. Environ. Biol. 2008, 14, 356–361. [Google Scholar]
- Fiola, J.A.; Hassan, M.A.; Swartz, H.J.; Bors, R.H.; McNicols, R. Effect of thidiazuron, light fluence rates and kanamycin on in vitro shoot organogenesis from excised Rubus cotyledons and leaves. Plant Cell Tissue Organ Cult. 1990, 20, 223–228. [Google Scholar]
- Krause, G.H. Photoinhibition of photosynthesis. An evaluation of damaging and protective mechanisms. Physiol. Plant. 1988, 74, 566–574. [Google Scholar] [CrossRef]
- Sáez, P.L.; Bravo, L.A.; Sánchez-Olate, M.; Bravo, P.B.; Ríos, D.G. Effect of Photon Flux Density and exogenous sucrose on the photosynthetic performance during in vitro culture of Castanea sativa. Am. J. Plant Sci. 2016, 7, 2087. [Google Scholar] [CrossRef] [Green Version]
- Kozai, T. Effects of CO2 enrichment and sucrose concentration under high photosynthesis photon fluxes on growth of tissue-cultured Cymbidium plantlets during the preparation stage. In Proceedings of the Congress ‘Plant Micropropagation in Horticultural Industries’, Arlon, Belgium, 10–14 August 1987; pp. 135–141. [Google Scholar]
- Kozai, T.; Oki, H.; Fujiwara, K. Photosynthetic characteristics of Cymbidium plantlet in vitro. Plant Cell Tissue Organ Cult. 1990, 22, 205–211. [Google Scholar] [CrossRef]
- Desjardins, Y.; Laforge, F.; Lussier, C.; Gosselin, A. Effect of CO2 enrichment and high photosynthetic photon flux on the development of autotrophy and growth of tissue-cultured strawberry, raspberry and asparagus plants. Acta Hortic. 1988, 230, 45–53. [Google Scholar] [CrossRef]
- Kozai, T.; Koyama, Y.; Watanabe, I. Multiplication of potato plantlets in vitro with sugar free medium under high photosynthetic photon flux. Acta Hortic. 1988, 230, 121–128. [Google Scholar] [CrossRef]
- McClung, C.R. Plant circadian rhythms. Plant Cell 2006, 18, 792–803. [Google Scholar] [CrossRef] [Green Version]
- Bell-Pedersen, D.; Cassone, V.M.; Earnest, D.J.; Golden, S.S.; Hardin, P.E.; Thomas, T.L.; Zoran, M.J. Circadian rhythms from multiple oscillators: Lessons from diverse organisms. Nat. Rev. Genet. 2005, 6, 544–556. [Google Scholar] [CrossRef] [PubMed]
- Devlin, P.F.; Kay, S.A. Cryptochromes are required for phytochrome signaling to the circadian clock but not for rhythmicity. Plant Cell 2000, 12, 2499–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodd, A.N.; Salathia, N.; Hall, A.; Kévei, E.; Tóth, R.; Nagy, F.; Hibberd, J.M.; Millar, A.J.; Webb, A.A. Plant circadian clocks increase photosynthesis, growth, survival, and competitive advantage. Science 2005, 309, 630–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loudon, A.S. Circadian biology: A 2.5 Billion year old clock. Curr. Biol. 2012, 22, R570–R571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmer, S.L.; Hogenesch, J.B.; Straume, M.; Chang, H.-S.; Han, B.; Zhu, T.; Wang, X.; Kreps, J.A.; Kay, S.A. Orchestrated transcription of key pathways in Arabidopsis by the circadian clock. Science 2000, 290, 2110–2113. [Google Scholar] [CrossRef] [PubMed]
- Barneche, F.; Malapeira, J.; Mas, P. The impact of chromatin dynamics on plant light responses and circadian clock function. J. Exp. Bot. 2014, 65, 2895–2913. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.Y.; Harmer, S.L. Wheels within wheels: The plant circadian system. Trends Plant Sci. 2014, 19, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Mermet, J.; Yeung, J.; Hurni, C.; Mauvoisin, D.; Gustafson, K.; Jouffe, C.; Nicolas, D.; Emmenegger, Y.; Gobet, C.; Franken, P. Clock-dependent chromatin topology modulates circadian transcription and behavior. Genes Dev. 2018, 32, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Abbott, A.; Belcher, A. Analysis of gases in culture flasks. In Report Long Ashton Research Station, Dawson & Goodall Ltd.; The Mendip Press: Bristol, UK, 1980; p. 79. [Google Scholar]
- Fujiwara, K. Measurements of carbon dioxide gas concentration in closed vessels containing tissue cultured plantlets and estimates of net photosynthetic rates of the plantlets. J. Agric. Methods 1987, 43, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Morini, S.; Muleo, R.; Sciutti, R.; Fortuna, P. Effect of different light-dark cycles on growth of fruit tree shoots cultured in vitro. Adv. Hort. Sci. 1990, 3, 1000–1004. [Google Scholar]
- Zimmerman, T.W.; Scorza, R. Benzyladenine and shortened light/dark cycles improve in vitro shoot proliferation of peach. HortScience 1994, 29, 698. [Google Scholar] [CrossRef]
- Morini, S.; Muleo, R.; Sciutti, R.; Fortuna, P. Relationship between evolution of CO2 and growth of plum shoot tips cultured in vitro under different light/dark regimes. Physiol. Plant. 1993, 87, 286–290. [Google Scholar] [CrossRef]
- Iarema, L.; da Cruz, A.C.F.; Saldanha, C.W.; Dias, L.L.C.; Vieira, R.F.; de Oliveira, E.J.; Otoni, W.C. Photoautotrophic propagation of Brazilian ginseng [Pfaffia glomerata (Spreng.) Pedersen]. Plant Cell Tissue Organ Cult. 2012, 110, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Badr, A.; Angers, P.; Desjardins, Y. Metabolic profiling of photoautotrophic and photomixotrophic potato plantlets (Solanum tuberosum) provides new insights into acclimatization. Plant Cell Tissue Organ Cult. 2011, 107, 13–24. [Google Scholar] [CrossRef]
- Kubota, C.; Kozai, T. Growth and net photosynthetic rate of Solanun tuberosum in vitro under forced and natural ventilation. HortScience 1992, 27, 1312–1314. [Google Scholar] [CrossRef] [Green Version]
- Schmildt, O.; Netto, A.T.; Schmildt, E.R.; Carvalho, V.S.; Otoni, W.C.; Campostrini, E. Photosynthetic capacity, growth and water relations in ‘golden’papaya cultivated in vitro with modifications in light quality, sucrose concentration and ventilation. Theor. Exp. Plant Physiol. 2015, 27, 7–18. [Google Scholar] [CrossRef]
- During, H. CO2 assimilation and photorespiration of grapevine leaves: Responses to light and drought. Vitis 1988, 27, 199–208. [Google Scholar]
- Düring, H.; Harst, M. Stomatal behaviour, photosynthesis and photorespiration of in vitro—Grown grapevines: Effects of light and CO2. Vitis 1996, 35, 163–168. [Google Scholar]
- Fujiwara, K.; Kozai, T.; Watanabe, I. Development of a photoautotrophic tissue culture system for shoots and/or plantlets at rooting and acclimatization stages. Acta Hortic. 1988, 230, 153–158. [Google Scholar] [CrossRef]
- Costa, A.C.; Rosa, M.; Megguer, C.A.; Silva, F.G.; Pereira, F.D.; Otoni, W.C. A Reliable methodology for assessing the in vitro photosynthetic competence of two Brazilian savanna species: Hyptis marrubioides and Hancornia speciosa. Plant Cell, Tissue Organ Cult. 2014, 117, 443–454. [Google Scholar] [CrossRef]
- Tamas, I.A. Hormonal regulation of apical dominance. In Plant Hormones; Springer: Berlin/Heidelberg, Germany, 1995; pp. 572–597. [Google Scholar]
- Chatfield, S.P.; Stirnberg, P.; Forde, B.G.; Leyser, O. The hormonal regulation of axillary bud growth in Arabidopsis. Plant J. 2000, 24, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Crawford, S.; Shinohara, N.; Sieberer, T.; Williamson, L.; George, G.; Hepworth, J.; Müller, D.; Domagalska, M.A.; Leyser, O. Strigolactones enhance competition between shoot branches by Dampening auxin transport. Development 2010, 137, 2905–2913. [Google Scholar] [CrossRef] [Green Version]
- Luisi, A.; Lorenzi, R.; Sorce, C. Strigolactone may interact with gibberellin to control apical dominance in pea (Pisum sativum). Plant Growth Regul. 2011, 65, 415–419. [Google Scholar] [CrossRef]
- Klee, H.J.; Lanahan, M.B. Transgenic plants in hormone biology. In Plant Hormones; Springer: Berlin/Heidelberg, Germany, 1995; pp. 340–353. [Google Scholar]
- Wareing, P.; Thompson, A.G. Rapid effect of red light on hormone levels. In Light and Plant Development; Smith, H., Ed.; Butterworths Publ. Co.: London, UK, 1976; pp. 284–295. [Google Scholar]
- Köhler, K.; Dörfler, M.; Göring, H. The influence of light on the cytokinin content of Amaranthus seedlings. Biol. Plant. 1980, 22, 128–134. [Google Scholar] [CrossRef]
- Lercari, B.; Micheli, P. Photoperiodic regulation of cytokin levels in leaf blades of Allium cepa L. Plant Cell Physiol. 1981, 22, 501–505. [Google Scholar]
- Stirk, W.; Bálint, P.; Tarkowská, D.; Novák, O.; Maróti, G.; Ljung, K.; Turečková, V.; Strnad, M.; Ördög, V.; Van Staden, J. Effect of light on growth and endogenous hormones in Chlorella minutissima (Trebouxiophyceae). Plant Physiol. Biochem. 2014, 79, 66–76. [Google Scholar] [CrossRef]
- Roman, H.; Girault, T.; Barbier, F.; Péron, T.; Brouard, N.; Pěnčík, A.; Novák, O.; Vian, A.; Sakr, S.; Lothier, J. Cytokinins are initial targets of light in the control of bud outgrowth. Plant Physiol. 2016, 172, 489–509. [Google Scholar] [CrossRef] [Green Version]
- Thimann, K. Hormone Action in the Whole Life of Plants; University of Massachusetts Press: Amherst, MA, USA, 1977. [Google Scholar]
- Koltai, H.; Kapulnik, Y. Strigolactones as mediators of plant growth responses to environmental conditions. Plant Signal. Behav. 2011, 6, 37–41. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.M.; Cochran, D.S.; Lamerson, P.M.; Evans, M.L.; Cohen, J.D. Red light-regulated growth: I. changes in the abundance of indoleacetic acid and a 22-Kilodalton Auxin-binding protein in the maize mesocotyl. Plant Physiol. 1991, 97, 352–358. [Google Scholar] [CrossRef] [Green Version]
- Mandava, N.B. Plant growth-promoting brassinosteroids. Annu. Rev. Plant Physiol. 1988, 39, 23–52. [Google Scholar] [CrossRef]
- Villalobos, V.M.; Leung, D.W.; Thorpe, T.A. Light-cytokinin interaction in shoot formation in cultured cotyledon explants of radiata pine. Physiol. Plant 1984, 61, 497–504. [Google Scholar] [CrossRef]
- Economou, A.; Read, P. Effect of red and far-red light on Azalea microcutting production in vitro and rooting in vivo. In Proceedings of the 6th International Congress Plant Tissue and Cell Culture, Minneapolis, MN, USA, 3–8 August 1986; Volume 431. [Google Scholar]
- Economou, A.S.; Read, P.E. Light treatments to improve efficiency of in vitro propagation systems. Hort. Sci. 1987, 22, 751–754. [Google Scholar]
- Teichmann, T.; Muhr, M. Shaping plant architecture. Front. Plant Sci. 2015, 6, 233. [Google Scholar] [CrossRef] [Green Version]
- Trouwborst, G.; Hogewoning, S.W.; van Kooten, O.; Harbinson, J.; van Ieperen, W. Plasticity of photosynthesis after the ‘red light syndrome’in cucumber. Environ. Exp. Bot. 2016, 121, 75–82. [Google Scholar] [CrossRef]
- Craver, J.K.; Miller, C.T.; Williams, K.A.; Bello, N.M. Ultraviolet radiation affects intumescence development in ornamental sweetpotato (Ipomoea batatas). HortScience 2014, 49, 1277–1283. [Google Scholar] [CrossRef]
- Nguyen, T.K.L.; Cho, K.M.; Lee, H.Y.; Cho, D.Y.; Lee, G.O.; Jang, S.N.; Lee, Y.; Kim, D.; Son, K.-H. Effects of white LED lighting with specific shorter blue and/or green wavelength on the growth and quality of two lettuce cultivars in a vertical farming system. Agronomy 2021, 11, 2111. [Google Scholar] [CrossRef]
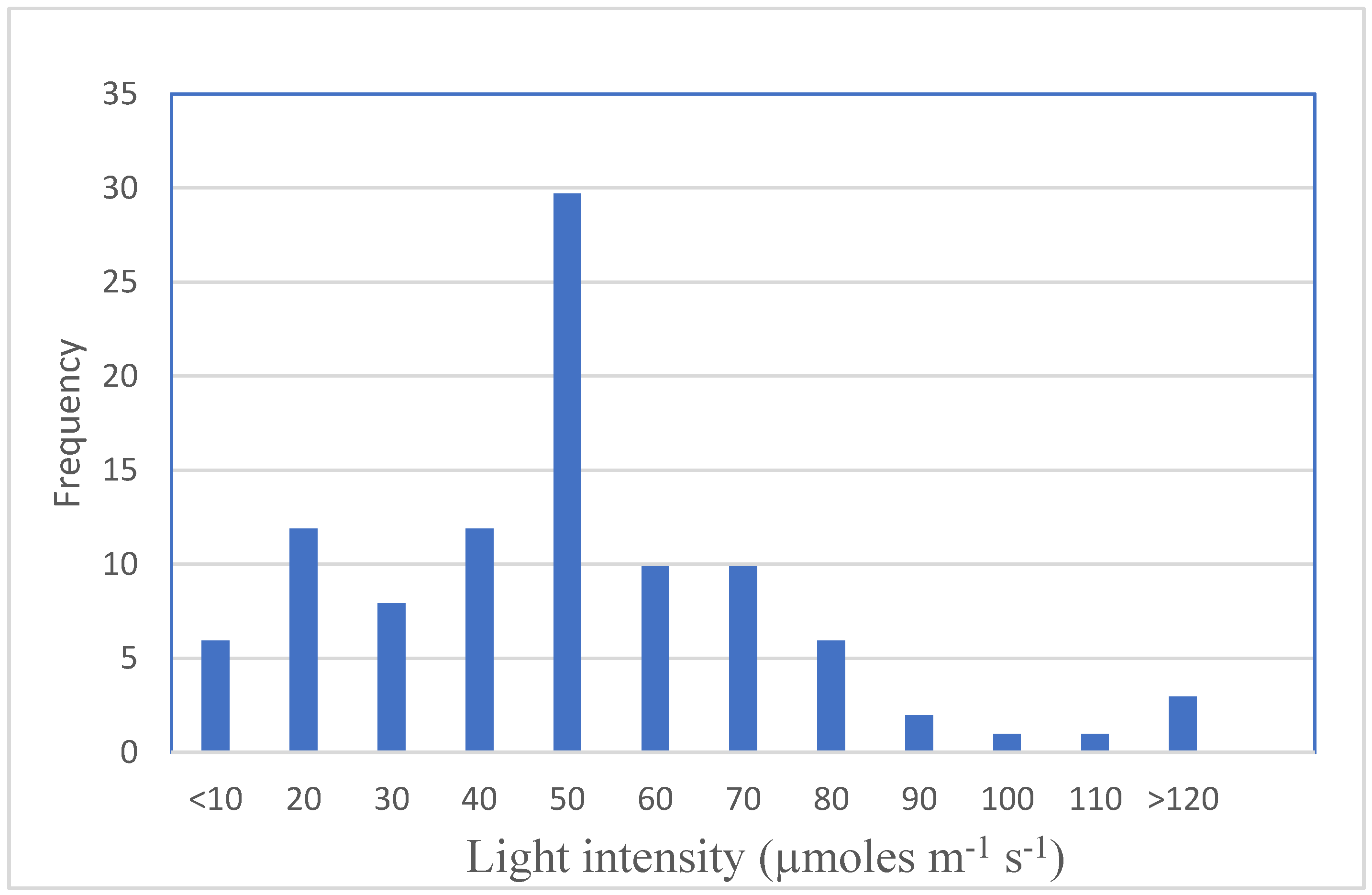

{kind=link}
| Studied Species/Explant Type | Light Intensity and Photoperiod | Light Spectra | Growth Regulators in Medium | Results on In Vitro Proliferation | Morphogenetic Response | Authors and Year |
|---|---|---|---|---|---|---|
| Nicotiana tabacum L. var. Wisconsin 38)/Callus | mW cm−2: 0, 0.0028; 0.024; 0.13; 0.37; 0.60; 0.80 photoperiod 16 h | 8 narrow band lights: 371, 419.5, 467, 504, 550, 590, 660, 750 nm, 4 commercial broad band-Fl lamps | For shoot differentiation: 2 mg L−1 K, 2 mg L−1 IAA, 80 mg L−1 adenine sulfate dihydrate | Near UV at low intensity (0.024 mw/cm2) and BL at higher intensities, callus growth and shoot initiation. | Higher carotenoids, porphyrins, associated with the high irradiance response. | [67] No LEDs |
| Vitis vinifera L. hybrid ‘Remaily Seedless’/Node shoots (axillary bud proliferation) | µW cm−2: 1500 for RL 1600 for BL light | RL BL No LED | BAP at 5 µM | BL = more shoots in the medium containing the lower concentration of manganese sulphate. | BL = larger shoots and more vigorous plantlets. | [68] No LEDs |
| Saintpaulia ionantha Wendl cv. Sona/leaves and Lycopersicon esculentum Mill./Cotyledons cv. UC 105 | Continuous light and daily light pulses | RL ad WL = highest bud regeneration in L. esculentum, BL in S. ionantha | [69] No LEDs | |||
| Vitis vinifera L. hybrid ‘Remaily Seedless’/Leaf axillary buds | 10-h and 16-h photoperiods | WL of various spectral irradiances, BL and RL light. | Apex removal from the explant was evaluated. | BL = best for shoot production. Under W, shoot production was greater with ratios of BL:RL of 0.6 to 0.9. | [70] No LEDs | |
| Solanum tuberosum L., cv. Miranda/Three- to four-node shoots (15 mm) | 160 µmol m−2 s−1 18 h (LD) or 10 h (SD). photoperiod | RL, BL | With or without 1 mg L−1 IAA or 1 mg L−1 K. | BL and K = better tuber production. RL and IAA application = high root/shoot ratio. Darkening strongly promoted tuber formation | Under BL, K increased total fresh weight and root (>stolons)/shoot ratio). | [71] No LEDs |
| Lavandin (Lavandula officinalis Chaix ×Lavandula latifolia Villars cv. Grosso)/Node explants | µmol m−2 s−1: Fl high fluence (HF) = 66 Fl low fluence (LF) = 7 RL (HF) = 7 RL (LF) = 1 FrL (HF) = 8 FrL (LF) = 2 BL (HF) = 13 BL (LF) = 1.5 UVL (HF) = 62 UVL (LF) = 5 | D control WL RL Fr L FrD (25 min Frh + 30 d D) FrRD (25 min Frh + 10 min R high + 30 d D) BL UV (UV A and B) | BA (l µM), putrescine (Put, 1 and 10 µM) | Low fluence RL = higher shoot number in presence or absence of BA. At low fluence rates also WL and BL enhanced shoot number on BA-free medium. 10 µM putrescine + Ba improved proliferation. | Rl and D positively affected shoot length. | [72] No LEDs |
| Begonia gracilis Kunth/Direct somatic embryogenesis from petiole explants. | 45 µmol m−2 s−l | RL and D | 0.5 mg L−1 kinetin | Somatic embryo production was higher under RL that in the dark. | [73] two cycles | |
| Azorina vidalii (Wats.) Feer (Dwarf shrub) | 50 µmol m−2 s−1: 16 h photoperiod | High and low ratios of BL + RL (2.3; 0.9) or RL + FRL (1.1; 0.6). Control: Fl | in vitro shoots no growth regulators | High ratio of RL/FRL light or BL/RL = the highest number of axillary shoots as compared to control. | Low ratio RL/FRL = maximum plant length and leaf area | [74] three months |
| Rhododendron spp./Axillary buds Disanthus cercidifolius Maxim./Shoot. Crataegus oxyacantha L./Axillary bud | µmol m−2 s−1: 11, 25, 55, 106 and 161 for Disanthus and Crataegus; 16, 26, 60 and 120 for Rhododendron | RL, GL and BL | Rhododendron 2.5 µM 2iP. Disanthus cercidifolius 3 µM BAP Crataegus oxyacantha 2.5 µM BAP and 0.5 µM IBA. | RL promoted axillary branching. All cultures grew well at low levels of irradiance | RL promoted shoot extension. | [75] No LEDS |
| Solanum lycopersicum cv. UC 105 an aurea (au) mutant and its isogenic wild type/Organogenesis from hypocotyls | µmol m−2 s−1: Fl = 50 0, 2.5 and 5 the other light treatments. 16 h photoperiod | D and Fl for aseptic seed germination RL, FRL, BL for regeneration. | Hormone free medium | All genotypes germinated under Fl. The wild type even under dark. Under RL, FRL and BL, hypocotyls showed a position-dependent regeneration. | [76] two cycles No LEDs | |
| Petunia x atkinsiana ‘Surfinia White’ cv.‘Revolution’/ Leaf explants | 19–21 µmol m−2 s−1 | WL, RL, BL, GL | 0.1 mg L−1 NAA, 1 mg L−1 BAP | Organogenesis was carried out in darkness. WL, GL and RL = the highest number of adventitious shoots. | Blue = the longest shoots and the biggest leaf area. | [77] |
| Lilium oriental hybrid ‘Pesaro’/In vitro-raised bulbs | 70 mmol m−2 s−1 12 h photoperiod | D, Fl, RL, BL, RL + BL (1:1). | 1.0 mg L−1 BA + 0.3 mg L−1 NAA | Fl, BL, and BL + Rl enhanced, plant regeneration as compared to D. | Bulblets under R + B were bigger in size, in fresh and dry weight. | [78] |
| Begonia erythrophylla J. Neuman/Petiole explants. | μmol m−2 s−1: WL, RL, and BL, and RL + BL = 35 Fr = 5 Continuous light | D, WL, R, B, RL + BL (1:1), FR | 0.54 mM NAA, 4.44 mM BA | RL or WL, as pre-treatments, promoted competence. RL or WL during culture, enhanced shoot number. | White light produced best developed and expanded shoots. | [79] No LEDS |
| Cymbidium Twilight Moon cv.‘Day Light’/ PLB segments. | 45 μmol m−2 s−1 16-h photoperiod | RL, RL + BL (3:1), RL + BL (50:50), RL + BL (1:3), BL. Control = Fl (PGF) | For callus induction from PLBs: 0.1 mg L−1 NAA and 0.01 mg L−1 TDZ For callus proliferation: 0.1 mg L−1 NAA and 0.01 mg L−1 TDZ. For PLBs production from callus: no growth regulators. | RL determined more callus induction; RL + BL (3:1) and PGF more callus proliferation RL + BL (1:3) more PLBs formation | [80] | |
| Lactuca sativa L./Cotyledon explants | 35 μmol m−2 s−1 | D, WL, RL, BL, BL + RL | 0.44mM BA, 0.54mM NAA | Light improved organogenesis as compared to D. RL and WL light promoted shoot production. | [81] No LEDS | |
| Fragaria × ananassa Duch. cv. Toyonoka/Leaf discs | 2000 lux | GL, RL, BL and YL Fl as control | 1.5 mg L−1 TDZ and 0.4 mg L−1 IBA. | Red and Green films determined the highest percentage of shoot regeneration and the max number of shoots per explant | RL and GL = a lower chlorophyll a/b ratio and higher antioxidant enzymes activity. | [82] No LEDs |
| Euphorbia milii Des Moul./Inflorescences Spathiphyllum cannifolium (Dryand. ex Sims) Schott/In vitro shoots | μmol m−2 s−1: 50 for Euphorbia: 35 for Spatifillum 16 h photoperiod | LEDS: RL, BL, RL + BL (1:1); BL + FrL (1:1); RL + FrL (1:1) Fl = Control | For E. miliii 1 mg L−1 BA, and 0.3 mg L−1 IBA. For S. cannifolium 3 mg L−1 BA, and 1 mg L−1 IBA. | S. cannifolium = best shoot proliferation under RL, RL + FRL. | For E.milii. BL = higher fresh and dry weight, and leaf number. For Spatifillum. BL= the highest chlorophyll and carotenoid contents. In both species, RL= higher plantlet length and higher fresh and dry weights. | [83] |
| Two species of Petunia: Petunia × atkinsiana (Sweet) D. Don and P. axillaris (Lam.)/Leaf tissue | 50 µmol m−2 s−1 16-h photoperiod | Fl, D | 5.7 μM IAA and 2.25 μM Zeatin. | Petunia × atkinsiana did not regenerate in darkness. Both species regenerate under light. | [84] | |
| Vitis vinifera L. cvs: Hybrid Franc, Ryuukyuuganebu (a wild grape native to Japan) and Kadainou R-1/Nodal segments | 50 µmol m−2 s−1 16-h photoperiod | RL and BL PGF light was used as control | PGR-free medium | No differences or slight differences on proliferation due to light treatments | RL = longest shoots. BL = higher chlorophyll content, leaf and stomata number per explant. | [85] |
| Phalaenopsis hybrid cv. Cassandra Rose/PLBs from in vitro germinated seeds and flower-stalk nodes. | RL, RL + BL (9:1, 8:2), RL + WL (1:1) Fl | RL + BL (8:2) = the highest PLBs development. RL + BL (9:1) = the highest shoots number. Shoot tips had higher PLBs induction under RL and BL. | RL and BL =the highest PLBs fresh weight. LED lights = more fresh weight, Height and leaf length. | [86] | ||
| Oncidium Sweets Sugar/Shoot apex | Fl (control), RL, BL | RL promoted PLB induction from shoot apex with the highest proliferation rate; BL the highest differentiation. | RL determined the highest content of carbohydrates. BL the highest protein content and enzyme activity. | [87] | ||
| Cymbidium finlaysonianum Lindl./PLBs | 16 h photoperiod | RL, Fl. | RL increased PLBs proliferation and number | [88] No LEDs | ||
| OncidiumGower Ramsey/Embryogenic calli | 50 µmol m−2 s−1 | D, Fl, BL, RL or RL + BL + Fr (RBFr) | 0.1 mg L−1 NAA and 0.4 mg L−1 BA | PLB formation and plantlet conversion was higher under (RBFr) LEDs and Fl. | RBFr enhanced leaf number and expansion, root, chlor. contents, fresh and dry weight. | [89] |
| Oncidium Gower Ramsey/Shoot tips | 11 µmol m−2 s−1 | Fl(control)RL, BL, YL and GL. | For PLBs induction, 1.0 mg L−1 BA, For PLB proliferation: 1.0 mg L−1 BA, 0.5 mg L−1 NAA. | RL enhanced PLB induction and multiplication, but low differentiation BL promoted PLbs differentiation into shoots | RL = the highest PLBs fresh weight and starch content. BL = higher chlorophyll, carotenoids and soluble protein content. | [90] |
| Cymbidium finlaysonianum Lindl., Cymbidium Waltz cv.‘Idol’, and Phalaenopsis cv:‘1327’/protocorm-like bodies (PLBs) | RL, BL and YL fluorescent films | RL and YL increased the number of PLBs of C. Waltz., RL, BL and YL increased the formation of shoots. RL and BL increased PLBs number in Phalaenopsis. | RL, BL and YL increased the fresh weight of PLBs in C.finlaysonianum. | [91] No LEDS | ||
| Dendrobium officinale Kimura & Migo/PLBs | 70 µmol m−2 s−1 16 h photoperiod | D, Fl, RL, BL; RL + BL (1:1); RL + BL (2:1); and RL + BL (1:2). | 0.5 g L−1 NAA, 0.2 g L−1, 6-BA | BL, RL + BL (1:1) and RL + BL (1:2) = higher percentage of PLBs producing shoots and the number of shoots produced per PLB | BL and different RL + BL ratios enhanced chlorophyll and carotenoids. BL, Fl, and RL + BL (1:2) produced higher dry matter. | [92] three cycles |
| Cymbidium insigne Rolfe/PLBs | WL, RL, BL and GL | Chondroitin sulfate The medium was added with Chitosan H or hyaluronic acid (HA9) | GL and 0.1 (mg L−1) and Chitosan H determined the highest PLBs and shoot formation. | Fresh weight of PLBs was higher at HA9 (1 mg L−1) treatment with GL. | [93] | |
| Ficus benjamina L. cv Exotica | BL, RL and FR. Fl as control | 0.5 mg L−1 IAA and 2 mg L−1 BA. | BL increased shoot number, and callus growth. | RL determined an increase in shoot length. | [94] | |
| Cymbidium Waltz cv ‘Idol’/5 mm protocorm-like bodies (PLBs) | 50 μmol m−2 s−1 16 h photoperiod | Fl, RL, BL, GL, Fl + GL, RL + GL, BL + GL. The last three treatment were subjected to 1d green exposure every 7d. | No growth regulators | RL + GL and BL promoted the highest PLB formation. Fl + GL and increased shoot formation from PLBs. | Fl gave the highest fresh weight. B + G the highest SOD activity. | [95] |
| Brassica napus L. cv Westar/Cotyledons from germinated seeds. | 60 μmol m−2 s−1 12 h photoperiod | Fl, BL, BL + RL (B:R = 3:1, 1:1, 1:3) RL. | For induction: 2,4-D in the dark; for shoots differentiation: 0.8 mg L−1 BA, 0.5 mg L−1 NAA; for shoots proliferation 1.0 mg L−1 BA. | The proliferation rate was greater under BL and BL:RL = 3:1 than under Fl | BL:RL (3:1) = higher fresh dry mass, chlorophyll a, soluble sugar, stem diameter, leaf stomata surface, than under Fl. Starch was higher in plantlets cultured under R light as compared to Fl. | [51] |
| Linum usitatissimum L., cv. ‘Szafir/Hypocotyls | 50 µmol m−2 s−1 | Light (Fl) or D conditions | 0.05 mg L−1 2,4-D and 1 mg L−1 BA | Shoot multiplication was about twice higher in light-grown cultures than those in darkness. | Fresh and dry mass and cyanogenic potential of light-grown cultures was about twice higher than those in the dark | [96] two cycles |
| Solanum tuberosum L. cvs Agrie Dzeltenie, Maret, Bintje, Désirée and Anti/Shoot tips from in vitro plantets | 40 µmol m−2 s−1 | Fl, warm WL light BL, RL, RL + BL (9:1 RB) and RL + BL + FR (70:10:20 RBF) | 0.5 mg L−1 zeatin riboside, 0.2 mg L−1, GA3 and 0.5 mg L−l IAA. | RL + BL (9:1) doubled the regeneration percentage of all cultivars after cryoconservation | [97] | |
| Abeliophyllum distichum Nakai,/Apical and axillary buds | 40 µmol m−2 s−1 | BL, RL + BL (1:1 RB), RL, Fl | BA 1.0 mg L−1, IBA 0.5 mg L−1 | BL and RL + BL promoted shoot proliferation. | RL increased shoot length. | [98] |
| Dendrobium kingianum Bidwill ex Lindl./PLBs | 50 μmol m−2 s−1 16 h photoperiod | RL, BL, RL + BL (1:1), GL and WL, Fl = control | MS medium supplemented with 412.5 mg/L ammonium nitrate, 950 mg/L potassium nitrate | BL and RL determined the highest PLBs number. RL and WL increased the percentage of shoot formation. | BL increased chlorophyll percentage, RL determined the highest fresh weight. | [99] |
| Cymbidium Waltz cv ‘Idol’ | 16 h photoperiod | GL, RL, BL | N- acetylglucosamine (NAG) 0, 0.01, 0.1, 1, and 10 mg L−1 | GL and RL + NAG determined the highest PLB formation rate RL or GL + NAG determined high shoot formation (80%) | Fresh weight of PLBs was highest at 0.01 mg L−1 NAG under green LED | [100] |
| Saccharum officinarum L., variety RB92579/in vitro grown plantlets | µmol m−2 s−1:
| (1) BL + RL (70:30) (2) BL + RL (50:50) (3) BL + RL (40:60) (4) BL + RL (30:70) (5) WL | 1.3 µM BAP. | BL + RL (70:30) gave the highest multiplication followed by 50:50. WL the lowest one. | BL + RL (70:30) and (50:50) = the highest total fresh weight. WL = the highest total chlorophyll content | [101] |
| Scrophularia takesimensis Nakai/Leaf, petiole, and stem explants | 45 µmol m−2 s−1 16 h photoperiod | Fl, RL, BL | 2.0 mg L−1 BA and 1.0 mg L−1 IAA | Fl = the highest number of shoots per leaf, petiole and stem explants | RL gave better shoot growth followed by Fl and BL. | [102] |
| Curculigo orchioides Gaertn./Leaf explants | 60 µmol m−2 s−1 | BL, RL, RL + BL (1:1). Fl as control. | 4 mg L−1 BA | BL determined the highest percentage of shoot organogenesis and shoot buds per explant. | [103] | |
| Fragaria x ananassa Duch. cv. ‘Camarosa’/Encapsulated shoot tips | 50 μmol m−2 s−1 16 h photoperiod | Fl (control) RL + BL (9:1 R9B1); RL + BL (7:3 R7B3); RL + BL (1:1 R5B5); RL + BL (3:7 R3B7); | Hormone free medium for plantlets development, and 4.9 µM IBA or 6.7 µM BA plus 2.3 µM K for shoots proliferation | RL + BL (1:9) were most effective for in vitro sprouting of encapsulated strawberry shoot tips. | R7B3 promoted shoot length, chlorophyll content, fresh and dry biomass accumulation. | [104] |
| Panax vietnamensis Ha et Grushv/Callus | 20–25 µmol m−2 s−1 16 h photoperiod | D, Fl, BL, GL, YL, RL, WL, and RL + BL: 90:10, 80:20, 70:30, 60:40, 50:50, 40:60, 30:70, 20:80, 10:90. | For embryogenic callus differentiation: 1 mg L−1 BA, 0.5 mg L−1 NAA. For plantlets differentiation: 0.5 mg L−1 BA, 0.5 mg L−1 NAA | YL most effective for callus production. RL + BL (6:4) was the most effective for differentiating the highest number of plants per explant from embryogenic callus. | YL gave the highest values of callus fresh and dry weight, followed by RL + BL (60:40). This last light gave the highest values of plantlet height, fresh and dry weight. | [105] |
| Vanilla planifolia Andrews./Axillary buds axillary bud cuttings | 25 µmol m−2 s−1 16 h photoperiod | BL, RL, RL + BL (1:1), WL, Fl | 9.55 µM BA | Fl, WL and RL + BL gave best results on shoot proliferation | Fl, WL and BL + RL determined higher shoot growth, plant height, leaves number, fresh weight, dry weight and chlorophyll content | [106] |
| Gerbera jamesonii Bolus ex Hooker f. cv Rosalin/In vitro propagated shoots | 140 ± 10 μmol m−2 s−1 | RL, BL, and their various mixtures. Fl was used as control | 1 mg L−1 BAP and 0.1 mg L−1 NAA | Fl lamps, BL, WL and RL + BL (70:30) = the highest number of shoots/explant and 70% R + 30%. | The same treatments also yielded the highest values in terms of shoot length, plant fresh and dry weight. | [107] |
| Cymbidium dayanum Rchb.f. and Cymbidium finlaysonianum Lindl./PLBs | 50 μmol m−2 s−1 16 h photoperiod | RL, BL, GL Fl. | (0, 0.1, 1 and 10 mg L−1), chondroitin sulfate | GL and BL + different concentrations of chondroitin sulfate promoted PLBs and shoots formation in the two species | [108] | |
| Bacopa monnieri L. (Water hyssop)/Full, upper and lower, leaf cuttings. | WL, RL + BL (4:1, 3:1, 2:1,1:1) | 0.25, 0.50 and 1.0 mg L−1 BA | WL was most effective in enhancing shoot regeneration. | Shoot length was increased by RL:BL (1:1) + 0.25 BA | [109] | |
| Vaccinium ashei Reade cv Titan | 50 µmol m−2 s−1 16 h photoperiod | Fl, RL, RL + BL (80:20) (R8B2), RL + BL (50:50 (R5B5), BL. | 1 mg L−1 zeatin riboside. Ventilated and non-ventilated vessels | No differences in shoot number between the different light treatments. | R8B2 and ventilated vessels were the most suitable for plant growth. | [110] |
| Anthurium andreanum Lind./Nodal segments | 25 μmol m−2 s−1 16 h light photoperiod | Fl, WL, RL, BL, BL + RL. | No growth regulators during the light treatments | BL + RL gave the highest number of adventitious shoots. | WL LEDs and BL LEDs, showed the greatest plantlet length and number of leaves. BL gave the greatest growth and chlorophyll content. | [111] |
| Saccharum officinarum L. variety RB867515) |
| BL:RL= (1) 70:30, (2) 50:50, (3) 30:70, (4) WL, (5) Fl | 1.3 μM BAP. | BL:RL = 50:50 promoted proliferation | BL:RL = 50:50 promoted the highest stem length, fresh mass production, leaf number. | [112] |
| Staphylea pinnata L./in vitro regenerated shoots | 35 μmol m−2 s−1 16 h photoperiod | Fl, RL + BL (50:50:1), RL + BL + FR (49:49:2) RL + BL + WL (40:40:20) | 5 µM BA, 0.5 µM NAA | Treatment with RB and RBFR resulted in increased multiplication rate as compared to Fl. | RB and RBFR increased leaf chlorophyll content and carotenoids. RBW light increased the number of newly developed leaves. | [113] |
| Stevia rebaudiana Bertoni/Nodal segments measuring 0.5–1 cm in length | 40–50 μmol m−2 s−1. 16 h light photoperiod | Fl (Control), BL, RL, RL + BL (1:1), WL | 1 mg L−1 BA. | RL = higher proliferation rate | Under BL + RL, maximum shoot elongation and leaf number | [114] |
| Vanilla planifolia Andrews/Nodal segments measuring 0.5–1 cm in length | 40 μmol m−2 s−1 16 h light photoperiod | Fl (control) BL, RL, RL + BL (1:1), WL | 2.1 mg L−1 BA | No differences in shoot multiplication. | BL enhanced leaf number and area. RL + BL enhanced shoot lengtht and chlorophyll content Fl determined higher fresh and dry weight and carotenoids. | [115] |
| Dendrobium sonia,/Mature PLBs | μmol m−2 s−1: W 17.7 B 22.5 Y 24.6 R 15.6 16 h photoperiod | WL (control), BL, YL, and RL. | 11.1 μM BAP and 11.42 μM IAA | YL induced early PLB formation, shoot differentiation and initiation, higher number of shoots per explant. | Under YL, higher leaf area and fresh weight, longer shoots under the other lights. | [116] |
| Nicotiana tabacum L. and Artemisia annua/In vitro-grown plantlets | 35 µmoles cm−1 s−1 | WL, RL + BL (1:1), RL + BL (3:1) RL + BL (1:3) | no growth regulators | In Nicotiana more shoots under 1:1 RL + BL In Artemisia under RL + BL (3:1) | In both species, RL + BL (3:1) determined taller shoots, and higher fresh weight. | [34] |
| Saccharum officinarum var. RB98710 (Sugarcane)/shoot segments | 50 µmol m−2 s−1 for FL, 80 µmol m−2 s−1 for LED 16-h photoperiod | Fl, WL, RL + BL (82:18). | For callus induction in the dark two substrates: C1 = 9 μM 2,4-D and 1.1 μM BA; C2 = 13.6 μM 2,4-D + 2.2 μM BAP. For shoot regeneration: hormone free medium. | LED were ineffective on somatic embryo regeneration but successful on shoot multiplication from somatic embryo. | Root length, number of leaves, shoot fresh and dry biomass did not differ between treatments. | [117] six subcultures |
| Gerbera jamesonii Bolus ex. Hook f. cv. Dura/in vitro propagated shoots | 40 μmol m−2 s−1 16-h photoperiod | BL, RL + BL1 (50:50), RL + BL2 (70:30), RL + BL + WL (40:40:20), RL + BL + FR (49: 49:2), RL, Fl (Control) | 5 μM BA (1,1 mg L−1) and 0.5 μM NAA (0.1 mg L−1) | RB1 and RB2 determined a higher shoot multiplication rate as compared to the control | RL = the greatest shoot elongation; BL = the highest leaf dry weight; RB2 = higher concentrations of total chlorophyll and carotenoids; RB1 = high leaf number. | [118] |
| Lippia gracilis Schauer./Apical and nodal segments | 42 μmol m−2 s−1 16 h photoperiod | WL, RL, BL, RL + BL (2.5:1 and 1:2.5) | no growth regulators | No influence of the light intensity nor of quality on shoot number both on nodal and apical segments. | RL and WL = best results on leaf and dry weights. B = higher photosynthetic pigment production in plantlets from apical explants, WL of those from nodal explants. | [119] |
| Myrtus communis L./Axillary shoots | 35 µmol m−2 s−1 16 h photoperiod | BL; RL:BL (70:30); RL; Fl = control. | 0.5 μM L−1 NAA and different concentrations of BA: 1, 2.5 and 5 µM. | RL and 5 µM BA resulted in the highest multiplication rate. | At 5 µM BA, RL determined the higher dry weight; BL = a greater leaves number, BL and RL:BL increased the FW compared to Fl. | [120] |
| Chrysanthemum × morifolium Ramat., Ficus benjamina L., Gerbera jamesonii Bolus f., Heuchera hybrida, and Lamprocapnos spectabilis (L.) Fukuhara. | 62–65 µM m−2 s−1 16 h photoperiod | Fl (control), NS1 lamps (BL + GL + RL + FRL- 21:38:35: 6) G2 lamps (BL + GL + RL+ FRL- 8:2:65:25), AP673L (BL + GL + RL + FRL- 12:19:61:8), AP67 (BL + GL + RL + FRL-14:16: 53: 17) | No PGRs for C. grandiflorum; 4.0 mg L−1 BA and 30 mg L−1 adenine sulfate for F. benjamina; 3.0 mg L−1 K. for G. jamesonii; 0.1 mg L−1 BA and 0.1 mg L−1 IAA for H. hybrida; 0.25 mg L−1 BA and 0.25 mg L−1 IAA for L. spectabilis | Except for F. benjamina, RL and G2 lamp gave highest or similar propagation ratios as compared to Fl. NS1 lamps was also efficient for G. jamesonii, H. hybrida and L. spectabilis | The highest chlorophyll content was recorded under Fl and AP673L in all species, in NS1 in two species. | [35] |
| Oryza sativa L. cultivar Nipponbare. | 50 μmol m−2 s−1. 12 h photoperiod | Fl, BL BL:RL = 3:1 BL:RL = 1:1; B:R = 1:3; RL; | For callus induction:2.0 mg L−1 2,4-D. For callus differentiation: 1.0 mg L−1 2,4-D. For shoot differentiation 0.5 mg L−1 K, 2 mg L−1 BA, 0.25 mg L−1 NAA | BL = decreased time for callus proliferation, differentiation and regeneration, and highest frequency of plantlet differentiation, and regeneration. | BL:RL = 1:1 highest seedling growth, chlorophyll, and carotenoid contents and photosynthetic rates. | [121] |
| Studied Species/Explant Type | Light Intensity and Photoperiod | Light Spectra | Growth Regulators in Medium | Results on In Vitro Proliferation | Morphogenetic Response | Authors and Year |
|---|---|---|---|---|---|---|
| Pseudotsuga menziesii Mirb. Douglas fir embryo | 0.01–0.71 W/cm2 16 h photoperiod | 8 different narrow bandwidth Fl having maxima each at one of the following wavelengths 371, 420, 467, 504, 550, 590, 660, and 740 nm. | Embryo from seeds; For callus induction: 800 pg L−1 IAA, 1 mg L−1 IBA, 1 mg L−1 BA, 1 mg L−1 AS-isopentyladenine After four weeks, 0.5 mM BA and 0.25 mM zeatin were added. No growth regulators for growing buds. | Callus and adventitious bud formation on the embryo-derived callus was maximum at (0.42 mW/cm−2) under RL (660 nm). | [122] | |
| Woody ornamental plants. Organogenesis (axillary bud proliferation) | Fl (control), high pressure sodium lamps (HPS), BL and RL | Light pipe modified growth chambers | HPS increased shoot number as compared to FL. RL increased shoot number over control. | [123] | ||
| Spirea nipponica Maxim/Shoot explants from 8 to 10 week-old stock cultures | WL: low fluence 15.0–23.0; high fluence 47.0–62.0 µmol m−2 s−1; RL+FR: low fluence) 8.7–15.9 µmol m−2 s−l 16 h photoperiod | WL, RL + Fr | BA 0.25, 0.4, or 0.5 mg L−1. | RL + FR = improved proliferation especially by 0.5 Ba addition. RL + FR followed by high fluence WL improved proliferation at lower BA levels. | RL + Fr favourably influenced shoot length and growth | [124] No LEDS |
| ‘Mr.S 2/5’ clone of Prunus domestica Ehrh./Cuttings; | WL = 38.0 BL = 9.1 RL = 19.6 FR = 7.2 µmol m−2 s−1 | WL BL RL FR | Ba 0.6 mg L−1 | In intact cuttings, WL gave the highest shoot proliferation In decapitated seedlings, all lights gave 100% bud outgrowth. | BL and WL = a higher number of nodes; RL = longer internodes. Shoots produced in RL were longer in decapitated seedlings. | [125] all experiments were repeated twice |
| Cydonia oblonga Mill/Leaves from the second to the fourth node of the apical portion of in vitro shoots | BL, WL and RL = 20 ± 1; FR = 1.2 R + B 10 + 10 B + Fr= 20 + 1.2 Fr + RL = 0.5 + 1.6 (µmol m−2 s−1) | D, BL, WL, FRL, RL, RL+Bl, BL+FRL RL+FRL After All light treatments, further 20 days of WL light exposure. | 4.7 µM K and 0.5 µM NAA | Somatic embryogenesis was highest under RL treatment. | [126] No LEDS | |
| Prunus avium L. cv ‘Hedelfinger’and one of its somatoclones/Leaves | ~9 µmol 16 h photoperiod | WL, RL, BL, FR, D | 2 mg dm3 TDZ+ 2,4-D or IAA | WL and BL = the highest node number. BL and FR = the highest shoot outgrowth from buds. | RL = highest shoot length under. WL and BL and WL= high chlorophyll. | [127] no LEDS |
| Malus domestica [Suckow] Borkh. genotype MM106/Shoot tips from in vitro cultures | ~40 μmol m−2 s−1 16 h photoperiod | WL, RL, BL, GL, YL, UV-AL, D | 8.86 (2 mg L−1) µM BA, 0.53 (0.06 mg L−1) µM Ga3, 0.3 µM (0.1 mg L−1) IBA | GL and WL gave the higher total number of shoots at the end of the fourth culturing cycle. | Leader stem height was greater under D, RL and YL. | [128] No LEDs Four cycles |
| Populus alba × P. berolinensis/Stems from in vitro shoots | 40 µmol m−2 s−1 16 h photoperiod | GL, RL, BL and YL. Fl (control) | 0.02 mg·L−1 NAA, and 0.1 mg·L−1 TDZ. | Fl and YL exhibited better effects on shoot regeneration | [129] no LEDs | |
| Musa spp. cv.’Grande naine’ AAA)/Meristematic shoot tips | 40 μmol m−2 s−1 16 h photoperiod | WL, Fl | 16.8 μM BAP, 3.8 μM IAA, 1 mg L−1 on a temporary immersion system (TIS) | WL under TIS enhanced shoot proliferation. | [130] | |
| Populus x euramericana selected clones, ‘I-476’ and ‘Dorskamp’/Petioles (5-mm long) from in vitro plants | 60 μm m−2 s−1 16 h photoperiod | Fl, BL, RL, RL +BL (1:1 and 7:3), and RL + BL + GL (7:2:1) | 0.44 µM BA | Highest shoot regeneration on RL + BL (1:1) for ‘I-476’, on BL +RL (7:3) for ‘Dorskamp’ as compared to Fl. | High RL (100% or 7:3) = higher shoot length and leaf area BL or RL +BL (7:3) = higher stem diameter | [131] |
| Malus domestica [Suckow] Borkh rootstock cvs. Budagovsky 9 (B.9), Geneva 30 (G.30), and Geneva 41 (G.41). I exp = single-node segments | BL = 5.7 RL = 6.6 WL = 25 μmol·m−2·s−1 | WL, RL, BL for both experiments | 1.0 mg·L−1 BA, 0.1 mg·L−1 IBA, and 0.5 mg·L−1 GA3. II exp: cv. G.30 with and without gibberellic acid (GA3). | RL increased the number of shoots in B.9 and G.30 as compared to WL. | RL increased the length, and the number of elongated shoots of B.9 and G.30. GA3 promoted shoot growth of G.30 under RL and BL. | [132] No LEDS |
| Phoenix dactylifera L. cv. ‘Alshakr’ (Date palm)/shoot buds | 20–25 μmol m−2 s−1 14 h photoperiod | FL (control), RL +BL (18:2) (CRB-LED) | 1 mg L−1 (NAA), 0.5 mg L−1 (BA) and 0.5 mg L−1 kinetin (K) | CRB enhanced the percentage of buds producing shoots and average shoots formation compared to FL | CRB-LED enhanced total soluble carbohydrates, starch, free amino acids, and peroxidase activity | [133] |
| Camellia oleifera C. Abel/Axillary buds | 50 m−2 s−1 16 h photoperiod | RL, BL, RL + BL, (4:1) RL + BL (1:4), WL was used as control | 3.0 mg L−1 BA + 0.02 mg L−1 IBA | RL + BL (4:1) = the highest proliferation coefficient. | RL + BL (4:1) = good chlorophyll content, the thickest leaves, high stomatal density. | [134] |
| Species | Tested Intensities | Best Yielding Intensity (μmol m−2 s−1) | Main Parameters Affected and Notes | Authors |
|---|---|---|---|---|
| Disanthus cercidifolius, Rhododendron spp., and Crataegus oxyacantha | 11, 25, 55, 106 and 161 µmol m−2 s−1 | 11–27 | Better growth and leaf chlorophyll content | [75] |
| Acer saccharum Marshall | 4, 16 and 40 µmol m−2 s−1 | 4 and 16 | Low intensity overcomes recalcitrance. | [214] |
| Achillea millefolium L. | 13; 27; 35; 47 and 69 µmol m−2 s−1 | 27 µmol m−2 s−1 | Higher dry mass of shoots and roots, shoot length | [172] |
| Withania somnifera (L.) | 15, 30, 60, and 90 µmol m−2 s−1 | 30 μmol m−2 s−1 | Greater growth and development. | [182] |
| Chrysanthemum morifolium Ramat. ‘Ellen’ | 25, 40, 55, 70, 55 µmol m−2 s−1 | 40 µmol m−2 s−1 | Better plantlet growth | [174] |
| Vaccinium corymbosum) | 55 to 240 µmol m−2 s−1 for 7 to 60 days | Higher irradiances (≥55 = 210 µmol m−2 s−1) improved proliferation only with short time applications (7 days). | [215] | |
| Spathiphyllum cannifolium Culture Pack”, on rockwool system, with CO2 enrichment | 45, 60, 75 μmol m−2 s−1 80% RL + 20% BL LED | 60 μmol m−2 s−1 | Best growth | [216] |
| Fragaria × ananassa Duchesne | 45, 60, 75 µmol m−2 s−1 | 60 μmol m−2 s−1 | Better shoot growth | [149] |
| Plectranthus amboinicus (Lour.) Sprengof | 26, 51, 69, 94 and 130 μmol m−2 s−1 | 69 µmol m−2 s−1 and to a lesser extend 94 | Higher shoot number, leaf area, total dry weight and carvacrol content | [48] |
| Phaius tankervilliae (Banks ex L’Herit) and Vanda coerulea Giff | 28, 37, 56, 74 and 93 μmol m−2 s−1 | 74 µmol m−2 s−1 | Better plantlet growth | [217] |
| Pyrus spp. rootstock BP10030 | from 10 to 80 μmol m−2 s−1 16 and 24 h photoperiod | from 10 to 80 μmol m−2 s−1 16 h photoperiod = greatest shoot number | 10 μmol m−2 s−1 better for initial explant growth. Increasing irradiance to max higher growth 24 h = the highest shoot fresh and dry weight. | [218] |
| Lippia gracilis Schauer | 26, 51, 69, 94, or 130 μmol m−2 s−1 | 94 µmol m−2 s−1 | higher number of segments, leaf, shoot, root, and total weight plantlet−1 | [119] |
| Momordica grosvenorii Swingle | 25, 50, 100, or 200 μmol·m−2·s−1, and an increased CO2 concentration | increasing intensities up to 100 µmol m−2 s−1 | Better plantlet growth | [219] |
| Actinidia deliciosa (A. Chev.) C.F. Liang & A.R. | 30 to 250 μmol m−2 s−1 and an increased CO2 concentration | 120 μmol m−2 s−1 | better plantlet growth and proliferation | [220] |
| Rosa hybrida | 0, 4, 17, 66, and 148 μE m−2 s−1 | 17 μE m−2 s−1 148 μE m−2 s−1 | At the highest intensity best proliferation. At 17 μE m−2 s−1 lower propagation but better leaves | [221] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavallaro, V.; Pellegrino, A.; Muleo, R.; Forgione, I. Light and Plant Growth Regulators on In Vitro Proliferation. Plants 2022, 11, 844. https://doi.org/10.3390/plants11070844
Cavallaro V, Pellegrino A, Muleo R, Forgione I. Light and Plant Growth Regulators on In Vitro Proliferation. Plants. 2022; 11(7):844. https://doi.org/10.3390/plants11070844
Chicago/Turabian StyleCavallaro, Valeria, Alessandra Pellegrino, Rosario Muleo, and Ivano Forgione. 2022. "Light and Plant Growth Regulators on In Vitro Proliferation" Plants 11, no. 7: 844. https://doi.org/10.3390/plants11070844
APA StyleCavallaro, V., Pellegrino, A., Muleo, R., & Forgione, I. (2022). Light and Plant Growth Regulators on In Vitro Proliferation. Plants, 11(7), 844. https://doi.org/10.3390/plants11070844