
Morphoanatomy and Histochemistry of Septal Nectaries Related to Female Fertility in Banana Plants of the ‘Cavendish’ Subgroup

 , and
, and
Abstract
:1. Introduction
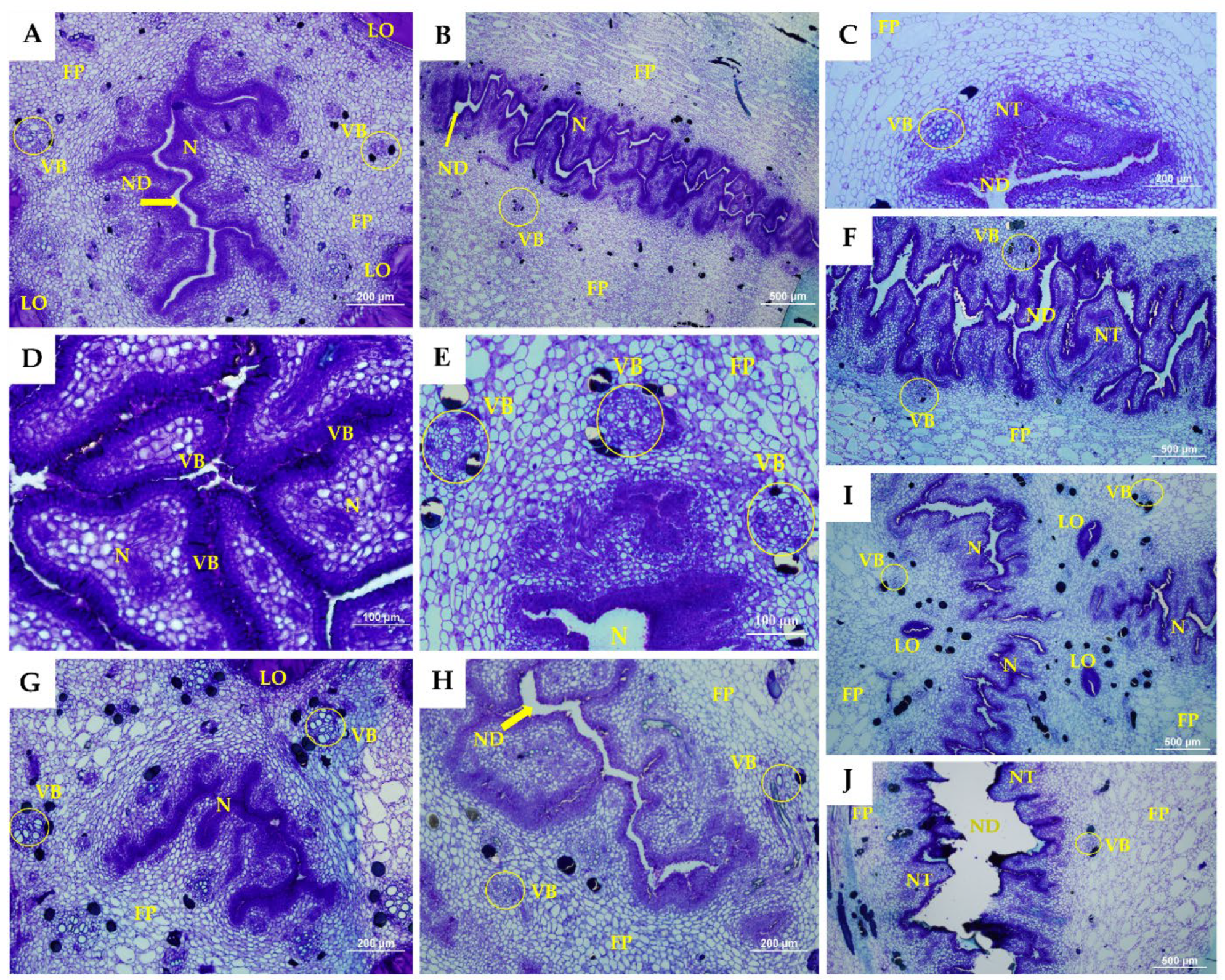
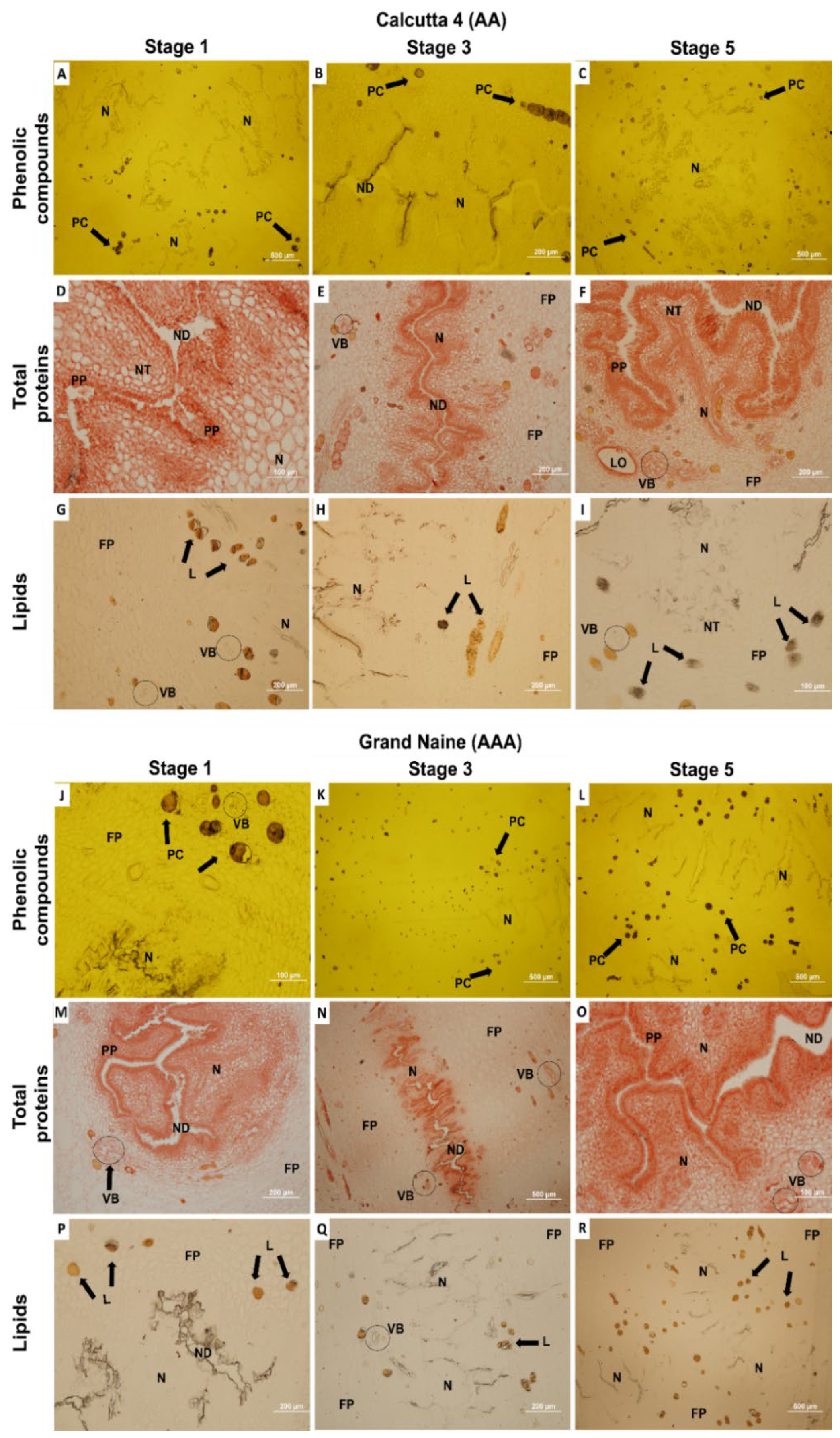
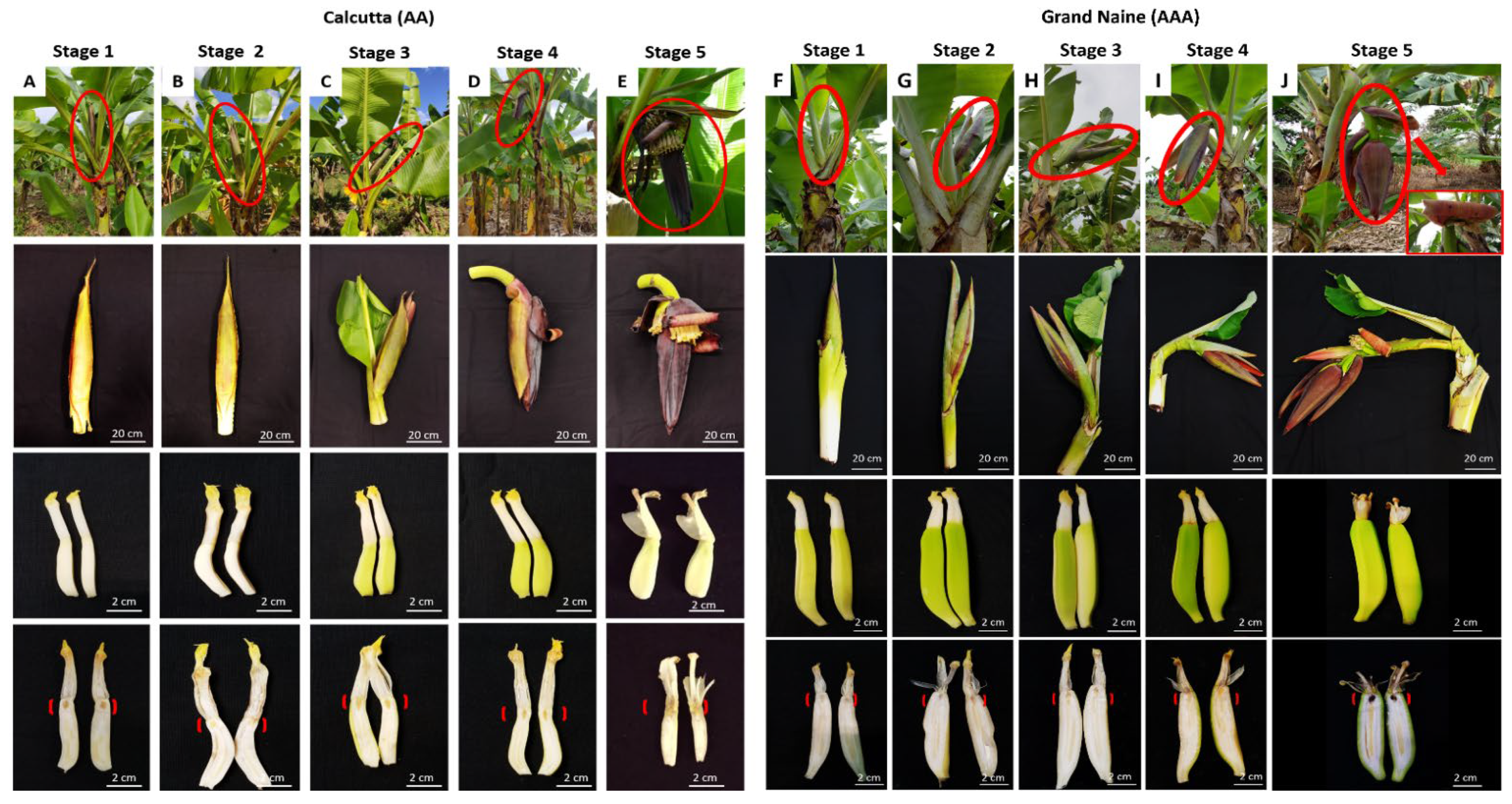
2. Results and Discussion
3. Materials and Methods
3.1. Plant Material
3.2. Flower Development Stages
3.3. Morphoanatomical and Histochemical Characterization
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castañeda, N.E.N.; Alves, G.S.C.; Almeida, R.M.; Amorim, E.P.; Ferreira, C.F.; Togawa, R.C.; Costa, M.M.C.; Grynberg, P.; Santos, J.R.P.; Cares, J.E.; et al. Gene expression analysis in Musa acuminata during compatible interactions with Meloidogyne incognita. Ann. Bot. 2017, 119, 915–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Agriculture Organization of the United Nations Statistics. Available online: http://www.fao.org/faostat/en/ (accessed on 13 January 2022).
- Silva, S.O.; Amorim, E.P.; Santos-Serejo, J.A.; Borges, A.L. Cultivares. In O Agronegócio da Banana; Ferreira, C.F., Silva, S.O., Amorim, E.P., Santos-Serejo, J.A., Eds.; Embrapa: Brasília, Brazil, 2016; pp. 111–136. [Google Scholar]
- Uwimana, B.; Zorrilla-Fontanesi, Y.; van Wesemael, J.; Mduma, H.; Brown, A.; Carpentier, S.; Swennen, R. Effect of seasonal drought on the agronomic performance of four banana genotypes (Musa spp.) in the East African Highlands. Agronomy 2021, 11, 4. [Google Scholar] [CrossRef]
- Ploetz, R.C. Fusarium wilt of banana. Phytopathology 2015, 105, 1512–1521. [Google Scholar] [CrossRef] [Green Version]
- Alakonya, A.E.; Kimunye, J.; Mahuku, G.; Amah, D.; Uwimana, B.; Brown, A.; Swennen, R. Progress in understanding Pseudocercospora banana pathogens and the development of resistant Musa germplasm. Plant Pathol. 2018, 67, 759–770. [Google Scholar] [CrossRef] [Green Version]
- Thangavelu, R.; Edwin Raj, E.; Pushpakanth, P.; Loganathan, M.; Uma, S. Draft genome of Fusarium oxysporum f. sp. cubenses train Tropical Race-4 infecting ‘Cavendish’ (AAA) groupof banana in India. Plant Dis. 2020, 105, 481–483. [Google Scholar] [CrossRef]
- de Jesus Rocha, A.; da Silva Soares, J.M.; dos Santos Nascimento, F.; Santos, A.S.; de Oliveira Amorim, V.B.; Ferreira, C.F.; Haddad, F.; dos Santos-Serejo, J.A.; Amorim, E.P. Improvements in the Resistance of the Banana Species to Fusarium Wilt: A Systematic Review of Methods and Perspectives. J. Fungi 2021, 7, 249. [Google Scholar] [CrossRef]
- Li, C.; Shao, J.; Wang, Y.; Li, W.; Guo, D.; Yan, B.; Xia, Y.; Peng, M. Analysis of banana transcriptome and global gene expression profiles in banana roots in response to infection by race 1 and tropical race 4 of Fusarium oxysporum f. sp. cubense. BMC Genom. 2013, 14, 851. [Google Scholar] [CrossRef] [Green Version]
- Lopes, O.P.; Maia, V.M.; Xavier, A.A.; Costa, M.R.; Rodrigues, M.G.V. Diversidade genética, crescimento e produção de genótipos de bananeira ‘Prata-Anã’ em área com mal do panamá. Rev. Bras. Frutic. 2014, 36, 924–939. [Google Scholar] [CrossRef] [Green Version]
- Dita, M.; Barquero, M.; Heck, D.; Mizubuti, E.S.G.; Staver, C.P. Fusarium wilt of banana: Current knowledge on epidemiology and research needs toward sustainable disease management. Front. Plant. Sci. 2018, 9, 1468. [Google Scholar] [CrossRef] [Green Version]
- Scheerer, L.; Pemsl, D.; Dita, M.; Perez Vicente, L.; Staver, C. A quantified approach to projecting losses caused by Fusarium wilt tropical race 4. Acta Hortic. 2018, 1196, 211–218. [Google Scholar] [CrossRef]
- Soares, J.M.S.; Rocha, A.J.; Nascimento, F.S.; Santos, A.S.; Miller, R.N.G.; Ferreira, C.F.; Haddad, F.; Amorim, V.B.O.; Amorim, E.P. Genetic Improvement for Resistance to Black Sigatoka in Bananas: A Systematic Review. Front. Plant Sci. 2021, 12, 657916. [Google Scholar] [CrossRef] [PubMed]
- Churchill, A.C.L. Mycosphaerella fijiensis, the black leaf streak pathogen of banana: Progress towards understanding pathogen biology and detection, disease development, and the challenges of control. Mol. Plant Pathol. 2011, 12, 307–328. [Google Scholar] [CrossRef]
- Amorim, E.P.; Amorim, V.B.O.; Silva, M.S.; Haddad, F.; Ferreira, C.F.; Santos-Serejo, J.A. Developing Hybrid Banana Varieties with Improved Properties; Germplasm and Genetic Improvement; Burleigh Dodds Science Publishing Limited: Cambridge, UK, 2020; Volume 2, pp. 323–338. [Google Scholar] [CrossRef]
- Fortescue, J.A.; Turner, D.W. Reproductive biology. In Banana Breeding: Progress and Challenges; Pillay, M., Tenkouano, A., Eds.; CRC Press: Boca Raton, FL, USA, 2011; Volume 1, pp. 145–179. [Google Scholar]
- Aguilar-Morán, J.F. Improvement of ‘Cavendish’ banana cultivars through conventional breeding. Acta Hortic. 2013, 986, 205–208. [Google Scholar] [CrossRef]
- Waniale, A.; Swennen, R.; Mukasa, S.B.; Tugume, A.K.; Kubiriba, J.; Tushemereirwe, W.K.; Batte, M.; Brown, A.; Tumuhimbise, R. Seed Set Patterns in East African Highland Cooking Bananas Show Asymmetric Distribution in Bunches and Fruits. Agronomy 2021, 11, 763. [Google Scholar] [CrossRef]
- Simmonds, N.W. The Evolution of the Bananas; Longmans: London, UK, 1962; p. 170. [Google Scholar]
- Shepherd, K.; Dantas, J.L.L.; Alves, E.J. Banana breeding in Brazil. In Banana and Plantain Breeding Strategies; Persley, G.J., De Langhe, E.A., Eds.; Australian Center for International Agricultural Research: Camberra, Australia, 1986; pp. 78–83. [Google Scholar]
- Fortescue, J.A.; Turner, D.W. The association between low temperatures and anatomical changes in preanthetic ovules of Musa (Musaceae). Sci. Hort. 2005, 104, 433–444. [Google Scholar] [CrossRef]
- Sardos, J.; Rouard, M.; Hueber, Y.; Cenci, A.; Hyma, K.E.; Houwe, I.V.D.; Hribova, E.; Courtois, B.; Roux, N. A Genome-Wide Association Study on the Seedless Phenotype in Banana (Musa spp.) Reveals the Potential of a Selected Panel to Detect Candidate Genes in a Vegetatively Propagated Crop. PLoS ONE 2016, 11, e0154448. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, K. Taxonomia e Caracterização de Cultivares de Banana; Embrapa CNPM: Cruz das Almas, Brazil, 1984; pp. 1–5. [Google Scholar]
- Soares, T.L.; Souza, E.H.; Costa, M.A.P.C.; Silva, S.O.; Santos-Serejo, J.A. In vivo fertilization of banana. Ciênc. Rural 2014, 44, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, A.O. Aspectos Reprodutivos e Fertilidade em Bananeiras Diploides e Triploides. Master’s Thesis, Federal University of Recôncavo da Bahia, Cruz das Almas, Brazil, 2015. [Google Scholar]
- dos Santos Silva, M.; Góes, N.H.; dos Santos-Serejo, J.A.; Ferreira, C.F.; Amorim, E.P. Phenolic compounds and oxidative enzymes involved in female fertility in banana plants of the cavendish subgroup. Plants 2021, 10, 2790. [Google Scholar] [CrossRef]
- Thompson, J.M. Studies in advancing sterility. Part VI. The theory of Scitaminean flowering. Publ. Hartley Bot. Lab. 1993, 11, 1–114. [Google Scholar]
- Juliano, J.B.; Alcala, P.E. Floral morphology of Musa errans (Blanco) Teodora var. botoan Teodoro. Philipp. Agric. 1933, 22, 91–116. [Google Scholar]
- Fahn, A.; Klarman-Kislev, N.; Ziv, D. The abnormal flower and fruit of May-Flowering Dwarf ‘Cavendish’ bananas. Bot. Gaz. 1961, 123, 116–125. [Google Scholar] [CrossRef]
- Fahn, A. Secretory Tissues in Plants; Academic Press: London, UK, 1979; p. 302. [Google Scholar]
- Schmid, R. Functional interpretations of the morphology and anatomy of septal nectaries. Acta Bot. Neerl. 1985, 34, 125–128. [Google Scholar] [CrossRef]
- Ren, Z.; Wang, H. Morphological comparison of floral nectaries in Musaceae, with reference to its pollinators. Biodivers. Sci. 2007, 15, 652–657. [Google Scholar] [CrossRef]
- Kirchoff, B.K. Ovary structure and anatomy in the Heliconiaceae and Musaceae (Zingiberales). Can. J. Bot. 1992, 70, 2490–2508. [Google Scholar] [CrossRef]
- Fahn, A.; Benouaiche, P. Ultrastructure, development and secretion in the nectary of banana flowers. Ann. Bot. 1979, 44, 85–93. [Google Scholar] [CrossRef]
- Roshchina, V.V.; Roshchina, V.D. The Excretory Function of Higher Plants; Springer: Berlin, Germany, 1993; p. 314. [Google Scholar]
- Smith, A.G.; Eberle, C.A.; Moss, N.G.; Anderson, N.O.; Clasen, B.M.; Hegeman, A.D. The transmitting tissue of Nicotiana tabacum is not essential to pollen tube growth, and its ablation can reverse prezygotic interspecific barriers. Plant Reprod. 2013, 26, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Souza, M.M.; Pereira, T.N.S.; Martins, E.R. Microsporogênese associadas ao tamanho do botão floral e da antera e viabilidade polínica em maracujá-amarelo (Passiflora edulis Sims f. flavicarpa Degener). Cienc. Agrotecnologia 2002, 26, 1209–1217. [Google Scholar]
- Lersten, N.R. Flowering Plant Embryology: With Emphasis on Economic Species; Blackwell Publishing: Ames, IW, USA, 2004. [Google Scholar]
- White, P.R. Studies on the Banana. An investigation of the floral morphology and cytology of certain types of the genus Musa L. Z. Zellforsch. Mikrosk. Anat. 1928, 7, 673–733. [Google Scholar] [CrossRef]
- Castro, P.R.C.; Kluge, R.A. Ecofisiologia de Fruteiras Tropicais: Abacaxizeiro, Maracujazeiro, Mangueira, Bananeira e Cacaueiro; Nobel: São Paulo, Brazil, 1998. [Google Scholar]
- Rocha, M.M. Extratos Florais de Bananeira e sua Influência na Germinação In Vitro de Grãos de Pólen. Master’s Thesis, Federal University of Recôncavo da Bahia, Cruz das Almas, Brazil, 2014. [Google Scholar]
- Veitch, N.C. Horseradish peroxidase: A modern view of a classic enzyme. Phytochemistry 2004, 65, 249–259. [Google Scholar] [CrossRef]
- Shetty, K. Role of proline-linked pentose phosphate pathway in biosyn thesis of plant phenolics for functional food and environmental applications: A review. Process Biochem. 2004, 39, 789–803. [Google Scholar] [CrossRef]
- Chunlong, C.; Song, L.; Rongsu, L.; Fengping, W.; Junqing, L. Concentration of phenolic compounds of Populus euphratica and soil water contents in Ejinaoasis, Inner Mongolia, China. Acta Ecol. Sin. 2008, 28, 69–75. [Google Scholar] [CrossRef]
- Locato, V.; Pinto, M.C.; Paradiso, A.; Gara, L. Reactive oxygen species and ascorbate-glutathione interplay in signaling and stress responses. In Reactive Oxygen Species and Antioxidants in Higher Plants; CRC Press: Boca Raton, FL, USA, 2010; pp. 45–64. [Google Scholar]
- Ellenman, C.J.; Dickinson, H.G. Pollen-stigma interactions in Brassica. IV. Structural reorganization in the pollen grains during hydration. J. Cell Sci. 1998, 80, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Wolters-Arts, M.; Lush, W.M.; Mariani, C. Lipids are required for directional pollen-tube growth. Nature 1998, 392, 818–821. [Google Scholar] [CrossRef] [PubMed]
- Backiyarani, S.; Sasikala, R.; Sharmiladevi, S.; Uma, S. Decoding the molecular mechanism of parthenocarpy in Musa spp. through protein–protein interaction network. Sci. Rep. 2021, 11, 14592. [Google Scholar] [CrossRef] [PubMed]
- Amah, D.; Turner, D.W.; Gibbs, D.J.; Wanilae, A.; Gram, G.; Swennen, R. Overcoming the fertility crisis in bananas (Musa spp.). In Achieving Sustainable Cultivation of Bananas; Germplasm and Genetic Improvement, Kema, G.H.J., Drenth, A., Eds.; Burleigh Dodds Science Publishing Limited: Cambridge, UK, 2021; Volume 2, pp. 257–306. [Google Scholar] [CrossRef]
- De Jesus, O.N.; de Oliveira e Silva, S.; Amorim, E.P.; Ferreira, C.F.; de Campos, J.M.S.; de Gaspari Silva, G.; Figueira, A. Genetic diversity and population structure of Musa accessions in ex situ conservation. BMC Plant Biol. 2013, 13, 41. [Google Scholar] [CrossRef]
- Silva, T.S.M.; Filho, M.A.C.; Coelho, E.F. Boletim Meteorológico da Estação Convencional de Cruz das Almas, BA. Variabilidade e tendências climáticas; Documento 216; Embrapa Mandioca e Fruticultura: Brasilia, Brazil, 2016; p. 79. [Google Scholar]
- Menezes, A.J.E.A.; Galvão, E.U.P. Bananeira: Recomendações de Cultivo; Comunicado Técnico; Embrapa: Brasilia, Brazil, 2004; Volume 113, pp. 1–4. [Google Scholar]
- Johansen, D.A. Plant Microtechnique; MacGraw-Hill: New York, NY, USA, 1940; p. 523. [Google Scholar]
- Gerrits, P.O. The Application of Glycol Methacrylate in Histotechnology: Some Fundamental Principles; Faculteit der Geneeskunde, Rijksuniversiteit: Groningen, The Netherlands, 1991; p. 160. [Google Scholar]
- O’brien, T.P.; Feder, N.; Mccully, M.E. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 1964, 59, 368–373. [Google Scholar] [CrossRef]
- Gerlach, D. Botanische Mikrotechnik; Georg Thieme Verlag: Stuttgart, Germany, 1969; p. 298. [Google Scholar]
- Pearse, A.G.E. Histochemistry; Theoretical Groningen and Applied, 3rd ed.; Little, Brown and Company: Boston, MA, USA, 1968; p. 1518. [Google Scholar]
- Cortelazzo, A.L.; Vidal, B.C. Soybean seed proteins: Detection in situ and mobilization during germination. Rev. Bras. Bot. 1991, 14, 27–33. [Google Scholar]
- Marques, J.P.R.; Nuevo, L.G. Double-Staining Method to Detect Pectin in Plant-Fungus Interaction. J. Vis. Exp. 2022, 180. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stages | Length (mm) | Diameter (mm) | ||
|---|---|---|---|---|
| Calcutta 4 | Grand Naine | Calcutta 4 | Grand Naine | |
| S1 | 3.85cB | 5.49bA | 0.93cA | 1.87cA |
| S2 | 4.44cB | 5.21cA | 1.68bA | 2.87cA |
| S3 | 5.27bA | 5.37cA | 1.70bA | 2.20cA |
| S4 | 6.86aA | 6.29bA | 2.07aA | 2.68bA |
| S5 | 7.35aA | 7.31aA | 2.17aB | 3.60aA |
| CV (%) | 20.49 | 33.61 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
dos Santos Silva, M.; Santana, A.N.; dos Santos-Serejo, J.A.; Ferreira, C.F.; Amorim, E.P. Morphoanatomy and Histochemistry of Septal Nectaries Related to Female Fertility in Banana Plants of the ‘Cavendish’ Subgroup. Plants 2022, 11, 1177. https://doi.org/10.3390/plants11091177
dos Santos Silva M, Santana AN, dos Santos-Serejo JA, Ferreira CF, Amorim EP. Morphoanatomy and Histochemistry of Septal Nectaries Related to Female Fertility in Banana Plants of the ‘Cavendish’ Subgroup. Plants. 2022; 11(9):1177. https://doi.org/10.3390/plants11091177
Chicago/Turabian Styledos Santos Silva, Manassés, Adriele Nascimento Santana, Janay Almeida dos Santos-Serejo, Claudia Fortes Ferreira, and Edson Perito Amorim. 2022. "Morphoanatomy and Histochemistry of Septal Nectaries Related to Female Fertility in Banana Plants of the ‘Cavendish’ Subgroup" Plants 11, no. 9: 1177. https://doi.org/10.3390/plants11091177
APA Styledos Santos Silva, M., Santana, A. N., dos Santos-Serejo, J. A., Ferreira, C. F., & Amorim, E. P. (2022). Morphoanatomy and Histochemistry of Septal Nectaries Related to Female Fertility in Banana Plants of the ‘Cavendish’ Subgroup. Plants, 11(9), 1177. https://doi.org/10.3390/plants11091177