
Predictive Modeling of Kudzu (Pueraria montana) Habitat in the Great Lakes Basin of the United States

Abstract
:1. Introduction
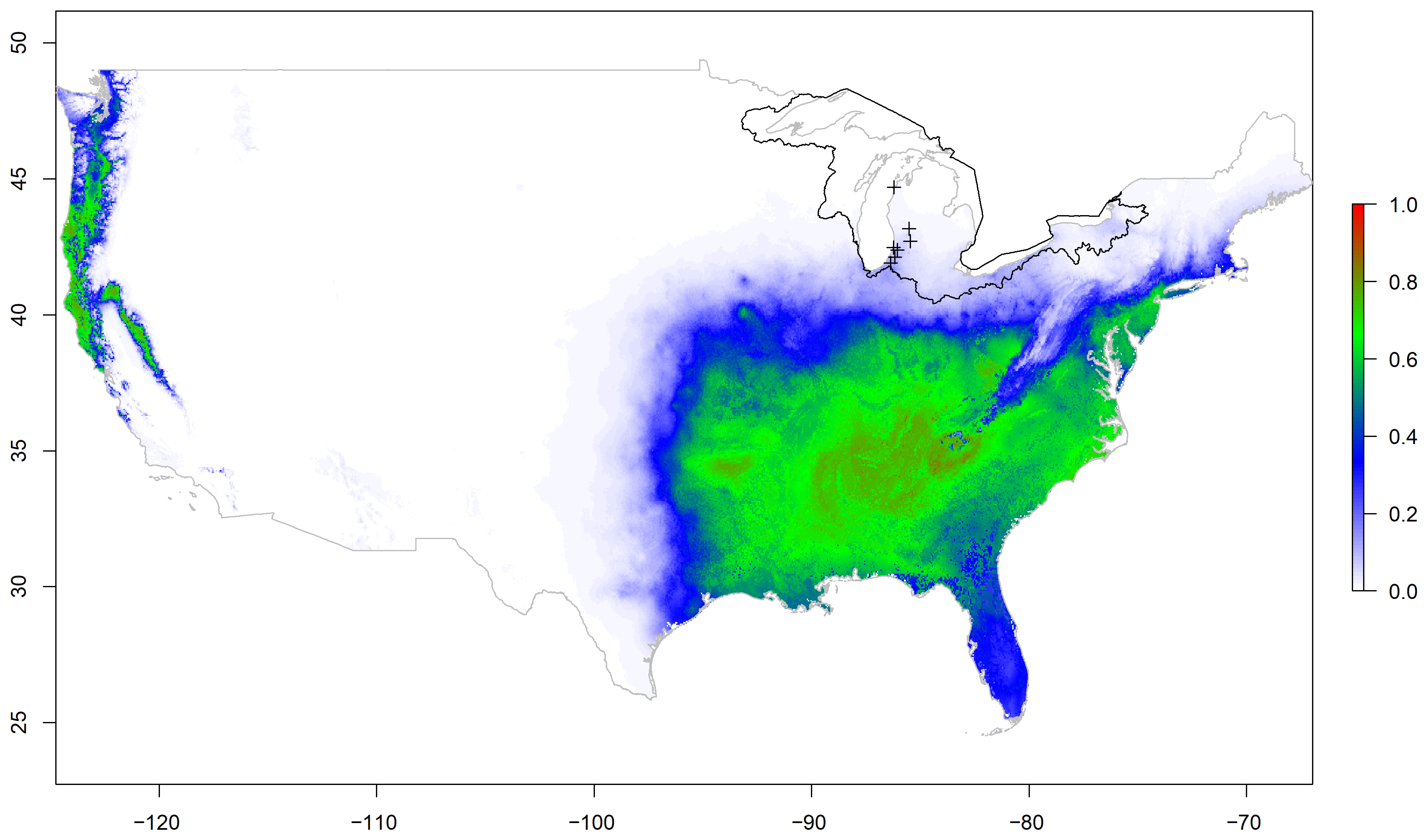
2. Results
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Millennium Ecosystem Assessment. Ecosystems and Human Well-Being: Biodiversity Synthesis; World Resources Institute: Washington, DC, USA, 2005. [Google Scholar]
- Pimentel, D.; Zungia, R.; Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Model 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Lodge, D.M.; Williams, S.; MacIssac, H.J.; Hayes, K.R.; Leung, B.; Reichard, S.; Mack, R.N.; Moyle, P.B.; Smith, M.; Andow, D.A.; et al. Biological invasions: Recommendations for US policy and management. Ecol. Appl. 2006, 16, 2035–2054. [Google Scholar] [CrossRef] [Green Version]
- Lindgren, C.J.; Castro, K.L.; Coiner, H.A.; Nurse, R.E.; Darbyshire, S.J. The biology of invasive alien plants in Canada. 12. Pueraria montana var. lobata (Willd.) Sanjappa & Predeep. Can. J. Plant Sci. 2013, 93, 71–95. [Google Scholar]
- Brown, K.H. Fair Japan: Japanese gardens at American World’s Fairs, 1876–1940. SiteLINES 2008, 4, 13–16. [Google Scholar]
- Guertin, P.J.; Denight, M.L.; Gebhart, D.L.; Nelson, L. Invasive species biology, control, and research. Part 1: Kudzu (Pueraria montana). Engineer Research and Development Center. US Army Corps of Engineers. Available online: https://apps.dtic.mil/sti/pdfs/ADA491410.pdf (accessed on 14 November 2022).
- Pieters, A.J. Kudzu, a Forage Crop for the Southeast; Leaflet No. 91; U.S. Department of Agriculture: Washington, DC, USA, 1932.
- Winberry, J.J.; Jones, D.M. Rise and decline of the “miracle vine”: Kudzu in the southern landscape. Southeast Geogr. 1973, 13, 61–70. [Google Scholar] [CrossRef]
- Callen, S.T.; Miller, A.J. Signatures of niche conservatism and niche shift in the North American kudzu (Pueraria montana) invasion. Divers. Distrib. 2015, 21, 853–863. [Google Scholar] [CrossRef]
- EDDMapS. Early Detection & Distribution Mapping System. The University of Georgia—Center for Invasive Species and Ecosystem Health. Available online: http://www.eddmaps.org/ (accessed on 14 November 2022).
- Boyette, C.D.; Walker, H.L.; Abbas, H.K. Biological control of kudzu (Pueraria lobata) with an isolate of Myrothecium verrucaria. Biocontrol Sci. Technol. 2002, 12, 75–82. [Google Scholar] [CrossRef]
- Mitich, L.W. Intriguing World of Weeds. Kudzu [Pueraria lobata (Willd.) Ohwi]. Weed Technol. 2000, 14, 231–235. [Google Scholar] [CrossRef]
- Britton, K.O.; Orr, D.; Sun, J. Kudzu. In Biological Control of Invasive Plants in Eastern United States; Van Driesche, R., Lyon, S., Blossey, B., Hoddle, M., Reardon, R., Eds.; USDA Forest Service: Morgantown, WV, USA, 2002; pp. 325–330. [Google Scholar]
- Forseth, I.N., Jr.; Innis, A.F. Kudzu (Pueraria montana): History, physiology, and ecology combine to make a major ecosystem threat. Crit. Rev. Plant Sci. 2004, 23, 401–413. [Google Scholar] [CrossRef]
- Munger, G.T. Pueraria montana var. lobata. Fire Effects Information System. USDA Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory. Available online: https://www.fs.usda.gov/database/feis/plants/vine/puemonl/all.html (accessed on 14 November 2022).
- Abramovitz, J.N. Pueraria lobata Willd. (OHWI), Kudzu: Limitations to Sexual Reproduction. Master’s Thesis, University of Maryland, College Park, MD, USA, 1983. [Google Scholar]
- Bentley, K.E.; Mauricio, R. High degree of clonal reproduction and lack of large-scale geographic patterning mark the introduced range of the invasive vine, kudzu (Pueraria montana var. lobata), in North America. Am. J. Bot. 2016, 103, 1499–1507. [Google Scholar]
- Coiner, H.A.; Hayhoe, K.; Ziska, L.H.; Van Dorn, J.; Sage, R.F. Tolerance of subzero winter cold in kudzu (Pueraria montana var. lobata). Oecologia 2018, 187, 839–849. [Google Scholar] [CrossRef] [PubMed]
- USDA. Plant Hardiness Zone Map. Agricultural Research Service, U.S. Department of Agriculture. Available online: https://planthardiness.ars.usda.gov/ (accessed on 18 November 2022).
- NOAA. Climate Data Online: Benzie County, MI (FIPS:26019). National Centers for Environmental Information. Available online: https://www.ncdc.noaa.gov/cdo-web/datasets/GHCND/locations/FIPS:26019/detail (accessed on 14 November 2022).
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Gastón, A.; García-Viñas, J.I. Modelling species distributions with penalized logistic regressions: A comparison with maximum entropy models. Ecol. Model 2011, 222, 2037–2041. [Google Scholar] [CrossRef]
- Baldwin, R.A. Use of maximum entropy modeling in wildlife research. Entropy 2009, 11, 854–866. [Google Scholar] [CrossRef]
- Bradley, B.A.; Wilcove, D.S.; Oppenheimer, M. Climate change increases risk of plant invasion in the Eastern United States. Biol. Invasions 2010, 12, 1855–1872. [Google Scholar] [CrossRef]
- Vázquez, D.P. Exploring the relationship between niche breadth and invasion success. In Conceptual Ecology and Invasion Biology; Cadotte, M.W., McMahon, S.M., Fukami, T., Eds.; Springer: Dordrecht, NL, USA, 2006; pp. 307–322. [Google Scholar]
- Freeman, E.A.; Moisen, G.G. A comparison of the performance of threshold criteria for binary classification in terms of predicted prevalence and kappa. Ecol. Model 2008, 217, 48–58. [Google Scholar] [CrossRef]
- Pörtner, H.-O.; Roberts, D.C.; Tignor, M.; Poloczanska, E.S.; Mintenbeck, K.; Alegría, A.; Craig, M.; Langsdorf, S.; Löschke, S. Climate Change 2022: Impacts, Adaptation and Vulnerability. In Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022. [Google Scholar]
- Widrlechner, M.P.; Daly, C.; Keller, M.; Kaplan, K. Horticultural applications of a newly revised USDA Plant Hardiness Zone Map. HortTechnology 2012, 22, 6–19. [Google Scholar] [CrossRef] [Green Version]
- Widrlechner, M.P. Environmental analogs in the stress-tolerant landscape plants. J. Arboric. 1994, 20, 114–119. [Google Scholar] [CrossRef]
- Geerts, S.; Mashele, B.V.; Visser, V.; Wilson, J.R.U. Lack of human-assisted dispersal means Pueraria montana var. lobata (kudzu vine) could still be eradicated from South Africa. Biol. Invasions 2016, 18, 3119–3126. [Google Scholar]
- Ehrlén, J.; Eriksson, O. Dispersal limitation and patch occupancy in forest herbs. Ecology 2000, 81, 1667–1674. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Dewitz, J. National Land Cover Database (NLCD) 2019 Products. US Geological Survey. Available online: https://doi.org/10.5066/P9KZCM54 (accessed on 15 November 2022).
- Ruefenacht, B.; Finco, M.V.; Nelson, M.D.; Czaplewski, R.; Helmer, E.H.; Blackard, J.A.; Holden, G.R.; Lister, A.J.; Salajanu, D.; Weyermann, D.; et al. Conterminous U.S. and Alaska forest type mapping using Forest Inventory Analysis data. Photogramm. Eng. Remote Sens. 2008, 74, 1379–1388. [Google Scholar] [CrossRef]
- Horton, J.D. The State Geologic Map Compilation (SGMC) Geodatabase of the Conterminous United States (ver. 1.1, August 2017), U.S. Geological Survey. Available online: https://www.sciencebase.gov/catalog/item/5888bf4fe4b05ccb964bab9d (accessed on 15 November 2022).
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. Dismo: Species Distribution Modeling. R Package Version 1.3-9. Available online: https://cran.r-project.org/packages=dismo (accessed on 11 November 2022).
- Merow, C.; Smith, M.J.; Silander, S.A., Jr. A practical guide to MaxEnt for modeling species distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 4th ed.; W.H. Freeman and Company: New York, NY, USA, 2012. [Google Scholar]
- Midwest Invasive Species Information Network. Michigan State University. Available online: https://www.misin.msu.edu/ (accessed on 11 November 2022).
- Pembury Smith, M.Q.R.; Ruxton, G.D. Effective use of the McNemar test. Behav. Ecol. Sociobiol. 2020, 74, 133. [Google Scholar] [CrossRef]
- Chicco, D.; Jurman, G. The advantages of the Matthews correlation coefficient (MCC) over F1 score and accuracy in binary classification evaluation. BMC Genom. 2020, 21, 6. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | National | Regional | Basin |
|---|---|---|---|
| Forest | 1.3 | 4.6 | 0.0 |
| Geology | 0.2 | 3.6 | 1.0 |
| NLCD | 0.0 | 2.2 | 7.4 |
| Precipitation | 81.3 | 0.4 | 1.2 |
| Temperature | 17.2 | 89.3 | 90.5 |
| Area (km2) | 7.80 × 106 | 1.12 × 106 | 4.48 × 105 |
| Kudzu Locations | 4263 | 164 | 10 |
| Background | 10,000 | 1436 | 574 |
| AUC | 0.795 | 0.906 | 0.900 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovach-Hammons, A.M.; Marshall, J.M. Predictive Modeling of Kudzu (Pueraria montana) Habitat in the Great Lakes Basin of the United States. Plants 2023, 12, 216. https://doi.org/10.3390/plants12010216
Kovach-Hammons AM, Marshall JM. Predictive Modeling of Kudzu (Pueraria montana) Habitat in the Great Lakes Basin of the United States. Plants. 2023; 12(1):216. https://doi.org/10.3390/plants12010216
Chicago/Turabian StyleKovach-Hammons, Ashley M., and Jordan M. Marshall. 2023. "Predictive Modeling of Kudzu (Pueraria montana) Habitat in the Great Lakes Basin of the United States" Plants 12, no. 1: 216. https://doi.org/10.3390/plants12010216
APA StyleKovach-Hammons, A. M., & Marshall, J. M. (2023). Predictive Modeling of Kudzu (Pueraria montana) Habitat in the Great Lakes Basin of the United States. Plants, 12(1), 216. https://doi.org/10.3390/plants12010216