
Movement of Lipid Droplets in the Arabidopsis Pollen Tube Is Dependent on the Actomyosin System


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
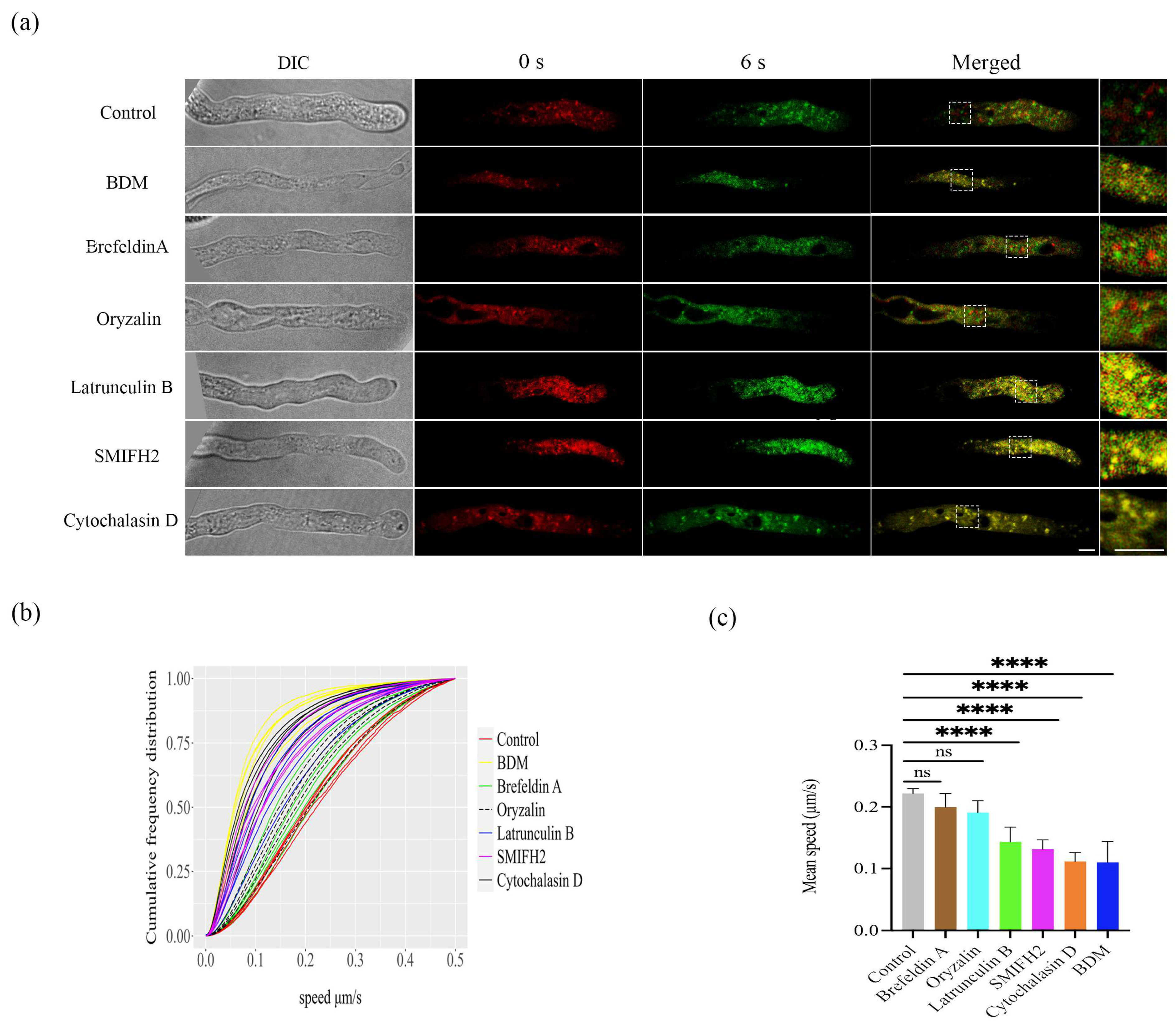
2.1. Tracking LDs Movement in Pollen Tubes in Arabidopsis thaliana
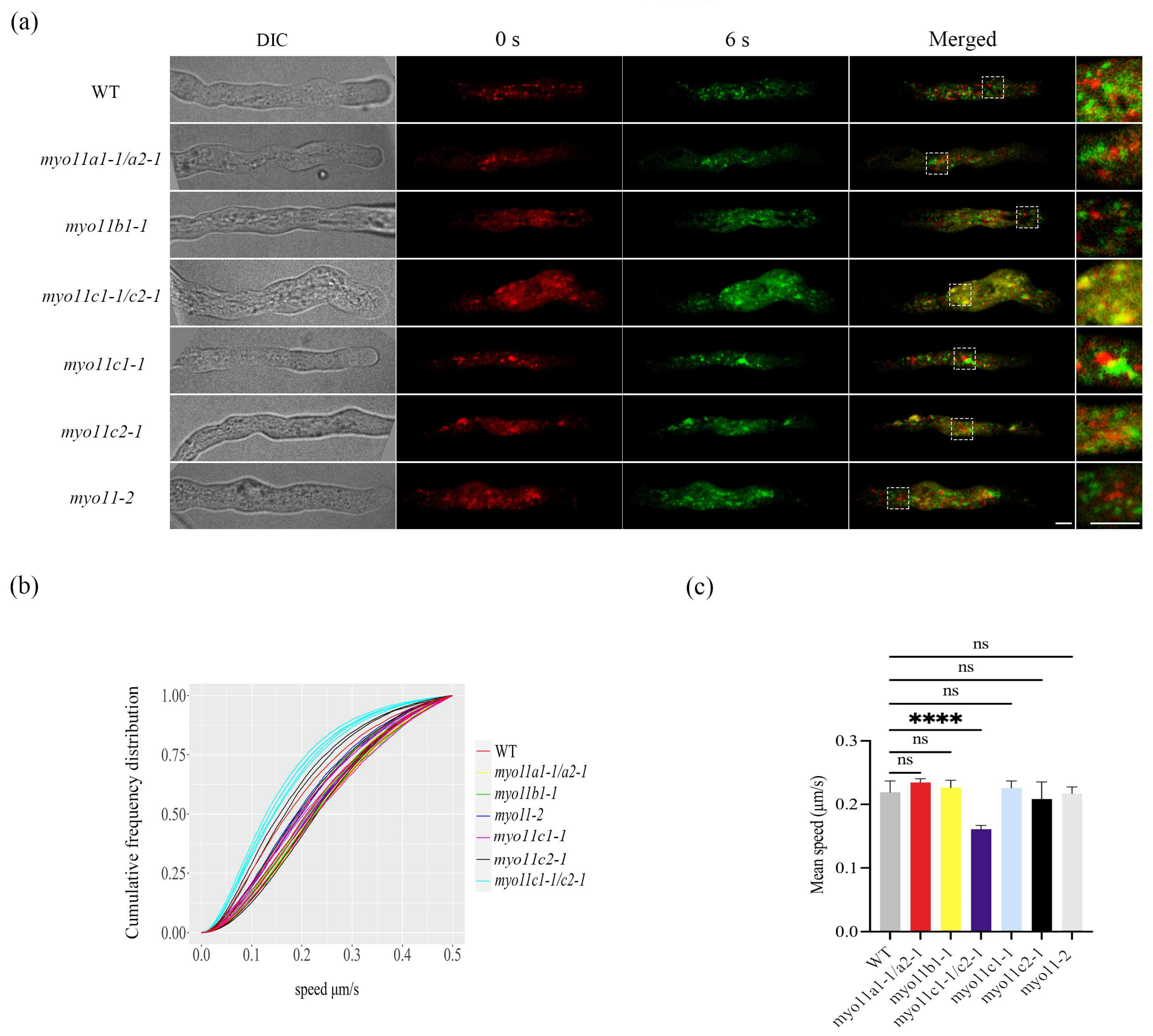
2.2. Mean Speed of the LDs in Pollen Tubes in WT and Myosin Mutants
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Pollen Germination
4.3. Nile Red Staining and Pharmacological Treatments
4.4. Microscope Images
4.5. Tracking LDs’ Movement with TrackMate
4.6. Data Analysis
4.7. Accession Numbers
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheung, A.Y.; Wu, H.M. Structural and signaling networks for the polar cell growth machinery in pollen tubes. Annu. Rev. Plant Biol. 2008, 59, 547–572. [Google Scholar] [CrossRef]
- Cheung, A.Y.; Wu, H.M. Structural and functional compartmentalization in pollen tubes. J. Exp. Bot. 2007, 58, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Ischebeck, T. Lipids in pollen—They are different. Biochim. Biophys. Acta 2016, 1861 Pt B, 1315–1328. [Google Scholar] [CrossRef]
- Rodriguez-Garcia, M.I.; M’Rani-Alaoui, M.; Fernandez, M.C. Behavior of storage lipids during development and germination of olive (Olea europaea L.) pollen. Protoplasma 2003, 221, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Murphy, D.J. The biogenesis and functions of lipid bodies in animals, plants and microorganisms. Prog. Lipid Res. 2001, 40, 325–438. [Google Scholar] [CrossRef] [PubMed]
- Ischebeck, T.; Krawczyk, H.E.; Mullen, R.T.; Dyer, J.M.; Chapman, K.D. Lipid droplets in plants and algae: Distribution, formation, turnover and function. Semin. Cell Dev. Biol. 2020, 108, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Nikiforidis, C.V. Structure and functions of oleosomes (oil bodies). Adv. Colloid Interface Sci. 2019, 274, 102039. [Google Scholar] [CrossRef]
- Wanner, G.; Formanek, H.; Theimer, R.R. The ontogeny of lipid bodies (spherosomes) in plant cells: Ultrastructural evidence. Planta 1981, 151, 109–123. [Google Scholar] [CrossRef]
- Parton, R.G.; Martin, S. Lipid droplets: A unified view of a dynamic organelle. Nat. Rev. Mol. Cell Biol. 2006, 7, 373–378. [Google Scholar]
- Choi, Y.J.; Zaikova, K.; Yeom, S.-J.; Kim, Y.-S.; Lee, D.W. Biogenesis and lipase-mediated mobilization of lipid droplets in plants. Plants 2022, 11, 1243. [Google Scholar] [CrossRef]
- Veerabagu, M.; Paul, L.K.; Rinne, P.L.; van der Schoot, C. Plant lipid bodies traffic on actin to plasmodesmata motorized by myosin XIs. Int. J. Mol. Sci. 2020, 21, 1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Schoot, C.; Paul, L.K.; Paul, S.B.; Rinne, P.L.H. Plant lipid bodies and cell-cell signaling: A new role for an old organelle? Plant Signal. Behav. 2011, 6, 1732–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhary, V.; Golani, G.; Joshi, A.S.; Cottier, S.; Schneiter, R.; Prinz, W.A.; Kozlov, M.M. Architecture of lipid droplets in endoplasmic reticulum Is determined by phospholipid intrinsic curvature. Curr. Biol. 2018, 28, 915–926.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farese, R.V.; Walther, T.C. Lipid droplets finally get a little RESPECT. Cell 2009, 139, 855–860. [Google Scholar] [CrossRef] [Green Version]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; Debono, A.; Durrett, T.P.; et al. Acyl-lipid metabolism. Arab. Book 2013, 11, e0161. [Google Scholar] [CrossRef] [Green Version]
- Chapman, K.D.; Ohlrogge, J.B. Compartmentation of triacylglycerol accumulation in plants. J. Biol. Chem. 2012, 287, 2288–2294. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Deng, X.; Qu, A.; Zhang, M.; Tao, Y.; Yang, L.; Liu, Y.; Xu, J.; Zhang, S. Regulation of pollen lipid body biogenesis by MAP kinases and downstream WRKY transcription factors in Arabidopsis. PLoS Genet. 2018, 14, e1007880. [Google Scholar] [CrossRef] [Green Version]
- Zienkiewicz, A.; Zienkiewicz, K.; Rejón, J.D.; Rodríguez-García, M.I.; Castro, A.J. New insights into the early steps of oil body mobilization during pollen germination. J. Exp. Bot. 2013, 64, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Müller, A.O.; Ischebeck, T. Characterization of the enzymatic activity and physiological function of the lipid droplet-associated triacylglycerol lipase AtOBL1. New Phytol. 2018, 217, 1062–1076. [Google Scholar] [CrossRef] [Green Version]
- Huang, A.H.C. Plant lipid droplets and their associated proteins: Potential for rapid advances. Plant Physiol. 2018, 176, 1894–1918. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Lim, S.-L.; Zhong, J.Y.; Lim, B.L. Bioenergetics of pollen tube growth in Arabidopsis thaliana revealed by ratiometric genetically encoded biosensors. Nat. Commun. 2022, 13, 7822. [Google Scholar] [CrossRef]
- Mellema, S.; Eichenberger, W.; Rawyler, A.; Suter, M.; Tadege, M.; Kuhlemeier, C. The ethanolic fermentation pathway supports respiration and lipid biosynthesis in tobacco pollen. Plant J. 2002, 30, 329–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, M.L.; Lima-Cabello, E.; Alché, J.D.; Martínez-Rivas, J.M.; Castro, A.J. Lipid composition and associated gene expression patterns during pollen germination and pollen tube growth in Olive (Olea europaea L.). Plant Cell Physiol. 2020, 61, 1348–1364. [Google Scholar] [CrossRef] [PubMed]
- Doniwa, Y.; Arimura, S.-i.; Tsutsumi, N. Mitochondria use actin filaments as rails for fast translocation in Arabidopsis and tobacco cells. Plant Biotechnol. 2007, 24, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Jedd, G.; Chua, N.H. Visualization of peroxisomes in living plant cells reveals acto-myosin-dependent cytoplasmic streaming and peroxisome budding. Plant Cell Physiol. 2002, 43, 384–392. [Google Scholar] [CrossRef] [Green Version]
- Ueda, H.; Yokota, E.; Kutsuna, N.; Shimada, T.; Tamura, K.; Shimmen, T.; Hasezawa, S.; Dolja, V.V.; Hara-Nishimura, I. Myosin-dependent endoplasmic reticulum motility and F-actin organization in plant cells. Proc. Natl. Acad. Sci. USA 2010, 107, 6894–6899. [Google Scholar] [CrossRef] [Green Version]
- Prokhnevsky, A.I.; Peremyslov, V.V.; Dolja, V.V. Overlapping functions of the four class XI myosins in Arabidopsis growth, root hair elongation, and organelle motility. Proc. Natl. Acad. Sci. USA 2008, 105, 19744–19749. [Google Scholar] [CrossRef] [Green Version]
- Kollmar, M.; Mühlhausen, S. Myosin repertoire expansion coincides with eukaryotic diversification in the Mesoproterozoic era. BMC Evol. 2017, 17, 211. [Google Scholar] [CrossRef] [Green Version]
- Reddy, A.S.; Day, I.S. Analysis of the myosins encoded in the recently completed Arabidopsis thaliana genome sequence. Genome Biol. 2001, 2, 1–17. [Google Scholar] [CrossRef]
- Haraguchi, T.; Ito, K.; Duan, Z.; Rula, S.; Takahashi, K.; Shibuya, Y.; Hagino, N.; Miyatake, Y.; Nakano, A.; Tominaga, M. Functional Diversity of Class XI Myosins in Arabidopsis thaliana. Plant Cell Physiol. 2018, 59, 2268–2277. [Google Scholar] [CrossRef] [Green Version]
- Madison, S.L.; Buchanan, M.L.; Glass, J.D.; McClain, T.F.; Park, E.; Nebenführ, A. Class XI myosins move specific organelles in pollen tubes and are required for normal fertility and pollen tube growth in Arabidopsis. Plant Physiol. 2015, 169, 1946–1960. [Google Scholar] [CrossRef] [Green Version]
- Peremyslov, V.V.; Prokhnevsky, A.I.; Avisar, D.; Dolja, V.V. Two class XI myosins function in organelle trafficking and root hair development in Arabidopsis. Plant Physiol. 2008, 146, 1109–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Gestel, K.; Köhler, R.H.; Verbelen, J.P. Plant mitochondria move on F-actin, but their positioning in the cortical cytoplasm depends on both F-actin and microtubules. J. Exp. Bot. 2002, 53, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.G.; Langhans, M.; Saint-Jore-Dupas, C.; Hawes, C. BFA effects are tissue and not just plant specific. Trends Plant Sci. 2008, 13, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Aloisi, I.; Cai, G.; Faleri, C.; Navazio, L.; Serafini-Fracassini, D.; Del Duca, S. Spermine regulates pollen tube growth by modulating Ca2+-dependent actin organization and cell wall structure. Front. Plant Sci. 2017, 8, 1701. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.; Zhang, R.; Zhang, M.; Diao, M.; Xue, Y.; Huang, S. Organizational innovation of apical actin filaments drives rapid pollen tube growth and turning. Mol. Plant 2017, 10, 930–947. [Google Scholar] [CrossRef] [Green Version]
- Cárdenas, L.; Lovy-Wheeler, A.; Kunkel, J.G.; Hepler, P.K. Pollen tube growth oscillations and intracellular calcium levels are reversibly modulated by actin polymerization. Plant Physiol. 2008, 146, 1611–1621. [Google Scholar] [CrossRef]
- Yu, L.; Fan, J.; Zhou, C.; Xu, C. Sterols are required for the coordinated assembly of lipid droplets in developing seeds. Nat. Commun. 2021, 12, 5598. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowska, M.; Popłońska, K.; Wojtczak, A.; Stȩpiński, D.; Polit, J.T. Lipid body biogenesis and the role of microtubules in lipid synthesis in Ornithogalum umbellatum lipotubuloids. Cell Biol. Int. 2012, 36, 455–462. [Google Scholar] [CrossRef]
- Groebner, J.L.; Girón-Bravo, M.T.; Rothberg, M.L.; Adhikari, R.; Tuma, D.J.; Tuma, P.L. Alcohol-induced microtubule acetylation leads to the accumulation of large, immobile lipid droplets. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G373–G386. [Google Scholar] [CrossRef]
- Romagnoli, S.; Cai, G.; Faleri, C.; Yokota, E.; Shimmen, T.; Cresti, M. Microtubule-and actin filament-dependent motors are distributed on pollen tube mitochondria and contribute differently to their movement. Plant Cell Physiol. 2007, 48, 345–361. [Google Scholar] [CrossRef]
- Basu, R.; Chang, F. Shaping the actin cytoskeleton using microtubule tips. Curr. Opin. Cell Biol. 2006, 19, 88–94. [Google Scholar] [CrossRef]
- Dogterom, M.; Koenderink, G.H. Actin-microtubule crosstalk in cell biology. Nat. Rev. Mol. Cell Biol. 2019, 20, 38–54. [Google Scholar] [CrossRef] [PubMed]
- Sampathkumar, A.; Lindeboom, J.J.; Debolt, S.; Gutierrez, R.; Ehrhardt, D.W.; Ketelaar, T.; Persson, S. Live cell imaging reveals structural associations between the actin and microtubule cytoskeleton in Arabidopsis. Plant Cell 2011, 23, 2302–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funaki, K.; Nagata, A.; Akimoto, Y.; Shimada, K.; Ito, K.; Yamamoto, K. The motility of Chara corallina myosin was inhibited reversibly by 2,3-butanedione monoxime (BDM). Plant Cell Physiol. 2004, 45, 1342–1345. [Google Scholar] [CrossRef] [Green Version]
- Sparkes, I.A. Motoring around the plant cell: Insights from plant myosins. Biochem. Soc. Trans. 2010, 38, 833–838. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Zhang, Y.; Kang, E.; Xu, Q.; Wang, M.; Rui, Y.; Liu, B.; Yuan, M.; Fu, Y. MAP18 regulates the direction of pollen tube growth in Arabidopsis by modulating F-actin organization. Plant Cell 2013, 25, 851–867. [Google Scholar] [CrossRef] [Green Version]
- Peremyslov, V.V.; Klocko, A.L.; Fowler, J.E.; Dolja, V.V. Arabidopsis myosin XI-K localizes to the motile endomembrane vesicles associated with F-actin. Front. Plant Sci. 2012, 3, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, X.; Wang, Q.; Wang, H.; Huang, S. Visualization of actin organization and quantification in fixed Arabidopsis pollen grains and tubes. Bio-Protocol 2020, 10, e3509. [Google Scholar] [CrossRef]
- Tinevez, J.-Y.; Perry, N.; Schindelin, J.; Hoopes, G.M.; Reynolds, G.D.; Laplantine, E.; Bednarek, S.Y.; Shorte, S.L.; Eliceiri, K.W. TrackMate: An open and extensible platform for single-particle tracking. Methods 2017, 115, 80–90. [Google Scholar] [CrossRef]
- Chustecki, J.M.; Gibbs, D.J.; Bassel, G.W.; Johnston, I.G. Network analysis of Arabidopsis mitochondrial dynamics reveals a resolved tradeoff between physical distribution and social connectivity. Cell Syst. 2021, 12, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Morgan, C.J. Use of proper statistical techniques for research studies with small samples. Am. J. Physiol.-Lung Cell Mol. Physiol. 2017, 313, L873–L877. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Liu, J.; Wong, C.-K.; Lim, B.L. Movement of Lipid Droplets in the Arabidopsis Pollen Tube Is Dependent on the Actomyosin System. Plants 2023, 12, 2489. https://doi.org/10.3390/plants12132489
Yang L, Liu J, Wong C-K, Lim BL. Movement of Lipid Droplets in the Arabidopsis Pollen Tube Is Dependent on the Actomyosin System. Plants. 2023; 12(13):2489. https://doi.org/10.3390/plants12132489
Chicago/Turabian StyleYang, Lang, Jinhong Liu, Ching-Kiu Wong, and Boon Leong Lim. 2023. "Movement of Lipid Droplets in the Arabidopsis Pollen Tube Is Dependent on the Actomyosin System" Plants 12, no. 13: 2489. https://doi.org/10.3390/plants12132489