
Variation in Susceptibility to Downy Mildew Infection in Spanish Minority Vine Varieties
 , ,
, ,  , , ,
, , ,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
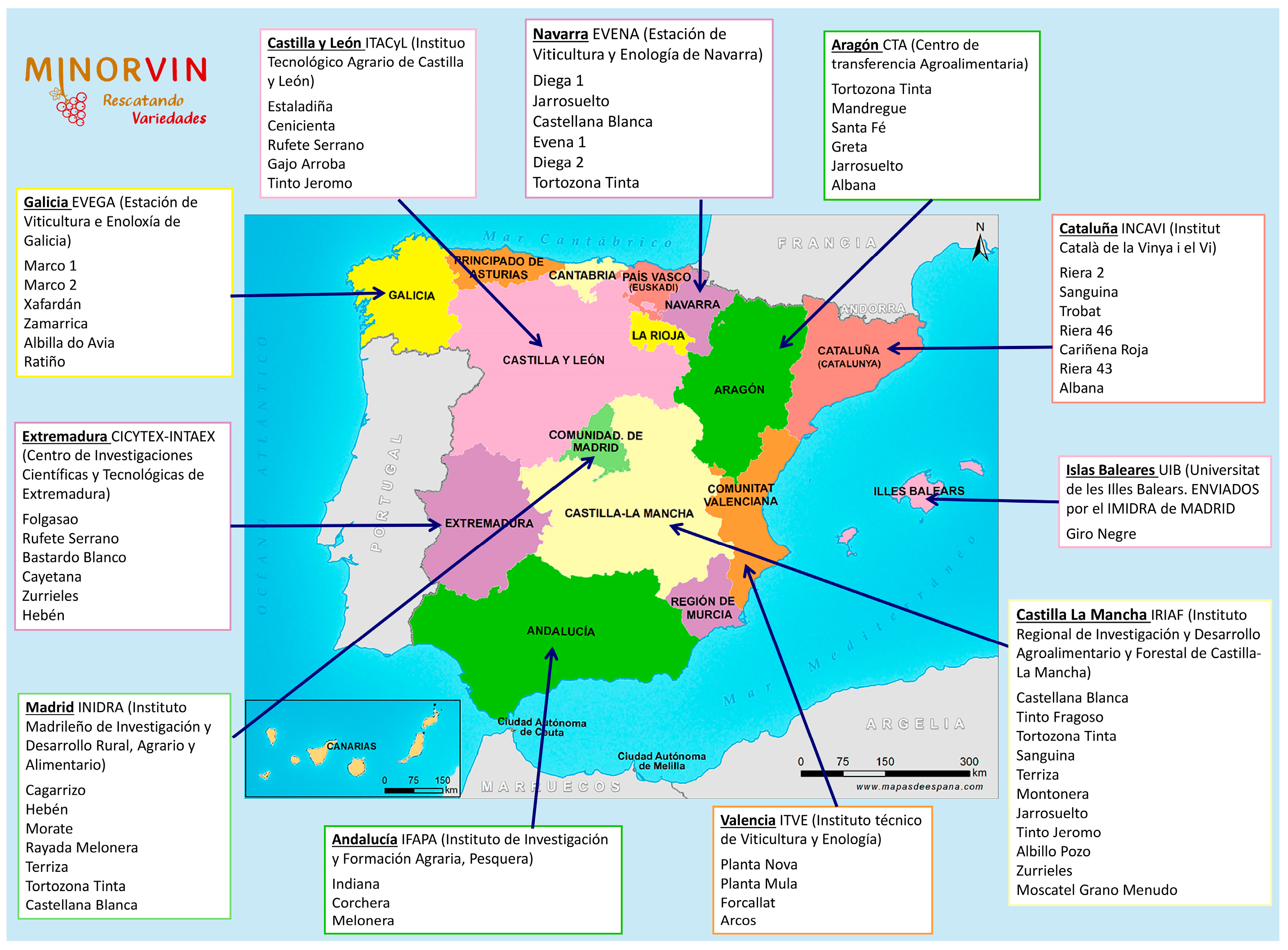
2.1. Plant Materials
2.2. Collection and Preservation of Plasmopara viticola
2.3. Inoculation of Leaf Discs
2.4. Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dick, M.W. Towards and understanding of the evolution of the downy mildew. In Advances in Downy Mildew Research; Spencer-Phillips, P.T.N., Gisi, U., Lebeda, A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 1–57. [Google Scholar]
- Boso, S.; Alonso-Villaverde, V.; Gago, P.; Santiago, J.; Martínez, M.C. Susceptibility of 44 grapevine (Vitis vinifera L.) varieties to downy mildew in the field. Aust. J. Grape Wine Res. 2011, 17, 394–400. [Google Scholar] [CrossRef]
- Boso, S.; Santiago, J.L.; Martínez, M.C. A method to evaluate downy mildew resistance in grapevine. Agron. Sustain. Dev. 2005, 25, 163–165. [Google Scholar] [CrossRef] [Green Version]
- Lacombe, T. Contribution à l’étude de l’histoire Evolutive de la Vigne Cultivée (Vitis vinifera L.) par l’analyse de la Diversité Genetique Neutre et de Gènes d’intérêt; Institut National d’Etudes Supérieures Agronomiques de Montpellier: Montpellier, France, 2012. [Google Scholar]
- Balda, P.; Martínez de Toda, F. Variedades Minoritarias de Vid en La Rioja; Consejería de Agricultura, Ganadería y Medio Ambiente; Gobierno de La Rioja: Logroño, Spain, 2017; 193p. [Google Scholar]
- Ferreira, V.; Pinto-Carnide, O.; Mota, T.; Martín, J.P.; Ortiz, J.M.; Castro, I. Identification of minority grapevine cultivars from Vinhos Verdes Portuguese DOC region. Vitis 2015, 54, 53–58. [Google Scholar]
- Santiago, J.L.; Boso, S.; Martínez, M.C.; Pinto-Carnide, O.; Ortiz, J.M. Ampelographic comparison of grape cultivars (Vitis vinifera L.) grown in northwestern Spain and northern Portugal. Am. J. Enol. Vitic. 2005, 56, 287–290. [Google Scholar] [CrossRef]
- Gago, P.; Santiago, J.L.; Boso, S.; Alonso-Villaverde, V.; Grando, S.; Martínez, M.C. Biodiversity and Characterization of Twenty-two Vitis vinifera L. Cultivars in the Northwestern Iberian Peninsula. Am. J. Enol. Vitic. 2009, 60, 293–301. [Google Scholar] [CrossRef]
- Martínez-Pinilla, O.; Guadalupe, Z.; Ayestarán, B.; Pérez-Magariño, S.; Ortega-Heras, M. Characterization of volatile compounds and olfactory profile of red minority varietal wines from La Rioja. J. Sci. Food Agric. 2013, 93, 3720–3729. [Google Scholar] [CrossRef]
- Pérez-Magariño, S.; Ortega-Heras, M.; Martínez-Lapuente, L.; Guadalupe, Z.; Ayestarán, B. Multivariate analysis for the differentiation of sparkling wines elaborated from autochthonous Spanish grape varieties: Volatile compounds, amino acids and biogenic amines. Eur. Food Res. Technol. 2013, 236, 827–841. [Google Scholar] [CrossRef]
- Zinelabidine, L.H.; Laiadi, Z.; Benmehaia, R.; Gago, P.; Boso, S.; Santiago, J.L.; Haddioui, A.; Ibáñez, J.; Martínez-Zapater, J.M.; Martínez, M.C. Comparative ampelographic and genetic analyses of grapevine cultivars from Algeria and Morocco. Aust. J. Grape Wine Res. 2014, 20, 324–333. [Google Scholar] [CrossRef]
- Martínez, M.C.; Boso, S.; Gago, P.; Muñoz-Organero, G.; De Andrés, M.T.; Gaforio, L.; Cabello, F.; Santiago, J.L. Value of two Spanish live grapevine collections in the resolution of synonyms, homonyms and naming errors. Aust. J. Grape Wine Res. 2018, 24, 430–438. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Moreno-Simunovic, Y. Terroir and typicity of Carignan from Maule Valley (Chile): The resurgence of a minority variety. OENO One 2019, 53, 75–93. [Google Scholar] [CrossRef] [Green Version]
- Merdinoglu, D.; Schneider, C.; Prado, E.; Wiedemann-Merdinoglu, S.; Mestre, P. Breeding for durable resistance to downy and powdery mildew in grapevine. OENO One 2018, 52, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Maul, E.; Töpfer, R. VIVC—Vitis International Variety Catalogue. Available online: http://www.vivc.de/ (accessed on 31 May 2019).
- Moreira, F.M.; Madini, A.; Marino, R.; Zulini, L.; Stefanini, M.; Velasco, R.; Kozma, P.; Grando, M.S. Genetic linkage maps of two interspecific grape crosses (Vitis spp.) used to localize quantitative trait loci for downy mildew resistance. Tree Genet. Genomes 2011, 7, 153–167. [Google Scholar] [CrossRef]
- Bove, F.; Savary, S.; Willocquet, L.; Rossi, V. Designing a modelling structure for the grapevine downy mildew pathosystem. Eur. J. Plant Pathol. 2020, 157, 251–268. [Google Scholar] [CrossRef]
- Foria, S.; Copetti, D.; Eisenmann, B.; Magris, G.; Vidotto, M.; Scalabrin, S.; Testolin, R.; Cipriani, G.; Wiedemann-Merdinoglu, S.; Bogs, J.; et al. Gene duplication and transposition of mobile elements drive evolution of the Rpv3 resistance locus in grapevine. Plant J. 2020, 101, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Sargolzaei, M.; Maddalena, G.; Bitsadze, N.; Maghradze, D.; Bianco, P.A.; Failla, O.; Toffolatti, S.L.; De Lorenzis, G. Rpv29, Rpv30 and Rpv31: Three Novel Genomic Loci Associated with Resistance to Plasmopara viticola in Vitis vinifera. Front. Plant Sci. 2020, 11, 562432. [Google Scholar] [CrossRef]
- Ruiz-García, L.; Gago, P.; Martínez-Mora, C.; Santiago, J.L.; Fernádez-López, D.J.; Martínez, M.D.C.; Boso, S. Evaluation and Pre-selection of New Grapevine Genotypes Resistant to Downy and Powdery Mildew, Obtained by Cross-Breeding Programs in Spain. Front. Plant Sci. 2021, 12, 674510. [Google Scholar] [CrossRef] [PubMed]
- Rumbolz, J.; Wirtz, S.; Kassemeyer, H.-H.; Guggenheim, R.; Schäfer, E.; Büche, C. Sporulation of Plasmopara viticola: Differentiation and Light Regulation. Plant Biol. 2002, 4, 413–422. [Google Scholar] [CrossRef]
- Staudt, G.; Kassemeyer, H.H. Evaluation of downy mildew resistance in various accessions of wild Vitis species. Vitis 1995, 34, 225–228. [Google Scholar]
- SAS Institute, Inc. SAS Online Doc, Version 8; SAS Institute, Inc.: Cary, NC, USA, 2000. [Google Scholar]
- Gutiérrez-Gamboa, G.; Liu, S.-Y.; Pszczólkowski, P. Resurgence of minority and autochthonous grapevine varieties in South America: A review of their oenological potential. Sci. Food Agric. 2020, 100, 465–482. [Google Scholar] [CrossRef]
- Galbiati, C. Structure di infezione di Plasmopora vitícola in diversi portainnesti di plante di vite. Riv. Patol. Veg. 1980, 16, 5–8. Available online: http://www.jstor.org/stable/42556771 (accessed on 25 May 2023).
- Bankovskaya, M.G.; Molchanova, Y.V. Evaluation of disease resistance of grape leaves under laboratory conditions. Vinograd Vino Ross. 1999, 5, 35–36. [Google Scholar]
- Galet, P. Précis de Viticulture, 7th ed.; Pierre Galet: Montpellier, France, 2000. [Google Scholar]
- Dai, G.H.; Andary, C.; Mondolot-Cosson, L.; Boubals, D. Histochemical studies on the interaction between three species of grapevine, Vitis vinifera, V. rupestris and V. rotundifolia and the Downy mildew fungus, Plasmopara viticola. Physiol. Mol. Plant Pathol. 1995, 46, 177–188. [Google Scholar] [CrossRef]
- Kortekamp, A.; Wind, R.; Zyprian, E. Investigation of the interaction of Plasmopara viticola with susceptible and resistant grapevine varieties. J. Plant Dis. Prot. 1998, 105, 475–488. [Google Scholar]
- Unger, S.; Büche, C.; Boso, S.; Kassemeyer, H.-H. The course of colonization of two different Vitis genotypes by Plasmopara viticola indicates compatible and incompatible host-pathogen interactions. Phytopathology 2007, 97, 780–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, M.M.; Castillo, C.; Menendez, C.M. Variability of Spanish minority varieties to Erysiphe necator infection. In Proceedings of the 11th International Symposium on Grapevine Physiology and Biotechnology, Stellenbosch, South Africa, 31 October–5 November 2021. [Google Scholar]
- Gaforio, L.; García-Muñoz, S.; Cabello, F.; Muñoz-Organero, G. Evaluation of susceptibility to powdery mildew (Erysiphe necator) in Vitis vinifera varieties. Vitis 2011, 50, 123–126. [Google Scholar]
- Delmas, C.E.L.; Fabre, F.; Jolivet, J.; Mazet, I.D.; Richart Cervera, S.; Delière, L.; Delmotte, F. Adaptation of a plant pathogen to partial host resistance: Selection for greater aggressiveness in grapevine downy mildew. Evol. Appl. 2016, 9, 709–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankel, E.N.; Waterhouse, A.L.; Teissedre, P.L. Principal phenolic phytochemicals in selected California wines and their antioxidant activity in inhibiting oxidation of human low-density lipoproteins. J. Agric. Food Chem. 1995, 43, 890–894. [Google Scholar] [CrossRef]
- Goldberg, D.M.; Yan, J.; Ng, E.; Diamandis, E.P.; Karumanchiri, A.; Soleas, G.; Waterhouse, A.L.A. Global survey of transresveratrol concentration in commercial wines. Am. J. Enol. Vitic 1995, 46, 159–165. [Google Scholar] [CrossRef]
- Pezet, R.; Gindro, K.; Viret, O.; Spring, J.-L. Glycosylation and oxidative dimerization of resveratrol are respectively associated to sensitivity and resistance of grapevine cultivars to downy mildew. Physiol. Mol. Plant Pathol. 2004, 65, 297–303. [Google Scholar] [CrossRef]
- Langcake, P.; Pryce, R.J. The production of resveratrol by Vitis vinifera and other members of the Vitaceae as a response to infection or injury. Physiol. Plant Pathol. 1976, 9, 77–86. [Google Scholar] [CrossRef]
- Montero, C.; Cristescu, S.M.; Jimenez, J.B.; Orea, J.M.; Lintel Hekkert, S.T.; Harren, F.J.M.; Gonzalez, U.A. Trans-resveratrol and grape resistance. A dynamic study by high-resolution laser-based techniques. Plant Physiol. 2003, 131, 129–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boso Alonso, S.; Alonso-Villaverde Iglesias, V.; Santiago Blanco, J.L.; Gago Montaña, P.; Dürrenberger, M.; Düggelin, M.; Martínez Rodríguez, M.C. Macro-and microscopic leaf characteristics of six grapevine genotypes (Vitis spp.) with different susceptibilities to grapevine downy mildew. Vitis 2010, 49, 43–50. [Google Scholar]
- Paolocci, M.; Muganu, M.; Alonso-Villaverde Iglesias, V.; Gindro, K. Leaf morphological characteristics and stilbene production differently affect downy mildew resistance of Vitis vinifera varieties grown in Italy. Vitis 2014, 53, 155–161. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Incidence | Severity (%) | Density (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Variety | Origin | Mean | SD | CV | Mean | SD | CV | Mean | SD | CV |
| Greta | DGA | 100.0 a | 0.0 | 0.0 | 37.5 efghijkl | 17.7 | 47.1 | 56.2 bcdefg | 23.9 | 42.5 |
| Moscatel G. Menudo | IVICAM | 100.0 a | 0.0 | 0.0 | 37.5 efghijkl | 14.4 | 38.5 | 50.0 bcdefgh | 28.9 | 57.7 |
| Ratiño | EVEGA | 100.0 a | 0.0 | 0.0 | 41.7 defghijk | 14.4 | 34.6 | 50.0 bcdefgh | 0.0 | 0.0 |
| Riera 2 | INCAVI | 100.0 a | 0.0 | 0.0 | 56.2 abcdef | 12.5 | 22.2 | 62.5 abcde | 25.0 | 40.0 |
| Rufete Serrano | CICYTEX | 100.0 a | 0.0 | 0.0 | 52.5 bcdefg | 16.6 | 31.6 | 62.5 abcde | 14.4 | 23.1 |
| Diega 1 | EVENA + UPNA | 98.4 ab | 3.1 | 3.2 | 53.1 abcdefg | 15.7 | 29.6 | 65.6 abcd | 12.0 | 18.2 |
| Giro Negre | UIB | 97.5 ab | 5.0 | 5.1 | 40.6 defghijk | 18.7 | 46.1 | 75.0 ab | 0.0 | 0.0 |
| Hebén | CICYTEX | 97.5 ab | 17.8 | 19.9 | 75.0 a | 23.9 | 34.8 | 75.0 ab | 12.5 | 18.2 |
| Santa Fe | DGA | 97.5 ab | 5.0 | 5.1 | 43.7 cdefghijk | 23.9 | 54.7 | 62.5 abcde | 25.0 | 40.0 |
| Tinto Jeromo | ITACYL | 97.0 ab | 3.6 | 3.7 | 56.5 abcdef | 16.1 | 28.7 | 56.2 bcdefg | 23.9 | 42.5 |
| Zurieles | CICYTEX | 97.0 ab | 6.0 | 6.2 | 50.0 bcdefghi | 0.00 | 0.0 | 68.7 abcd | 12.5 | 18.2 |
| Evena 1 | EVENA + UPNA | 96.2 ab | 7.5 | 7.8 | 40.0 efghijk | 12.2 | 30.6 | 50.0 bcdefgh | 20.4 | 40.8 |
| Castellana Blanca | IVICAM | 95.7 ab | 8.6 | 8.9 | 29.0 ijklmn | 8.9 | 30.7 | 43.7 dfgh | 12.5 | 28.5 |
| Indiana | IFAPA | 95.2 ab | 3.5 | 3.6 | 59.4 abcde | 12.0 | 20.2 | 62.5 abcde | 14.4 | 23.1 |
| Albana | INCAVI | 95.0 abc | 9.9 | 10.4 | 51.5 bcdefgh | 22.1 | 42.6 | 62.5 abcde | 14.4 | 23.1 |
| Albillo del Pozo | IVICAM | 95.0 abcd | 5.77 | 6.08 | 56.2 abcdef | 23.9 | 42.5 | 56.2 bcdefg | 12.5 | 22.2 |
| Tortozona Tinta | IMIDRA | 95.0 abcd | 10.0 | 10.5 | 43.7 cdefghijk | 12.5 | 28.6 | 56.2 bcdefg | 12.5 | 22.2 |
| Jarrosuelto | DGA | 93.7 abcde | 12.5 | 13.3 | 50.0 bcdefghi | 20.4 | 40.8 | 75.0 ab | 0.0 | 0.0 |
| Tortozona Tinta | DGA | 93.7 abcde | 12.5 | 13.3 | 50.0 bcdefghi | 10.2 | 20.4 | 62.5 abcde | 14.4 | 23.1 |
| Zurieles | IVICAM | 93.7 abcde | 12.5 | 13.3 | 59.4 abcde | 23.7 | 39.8 | 75.0 ab | 20.4 | 27.2 |
| Forcallat | ITVE | 93.6 abcde | 11.0 | 11.7 | 53.3 abcdefg | 20.2 | 37.89 | 70.8 abc | 7.2 | 10.2 |
| Montonera del Casar | IVICAM | 93.2 abcde | 9.4 | 10.1 | 40.6 defghijk | 23.7 | 58.2 | 62.5 abcde | 14.4 | 23.1 |
| Cenicienta | ITACYL | 92.7 abcde | 9.1 | 9.9 | 27.5 ijklmn | 26.0 | 94.5 | 50.0 bcdefgh | 20.4 | 40.8 |
| Corchera | IFAPA | 92.5 abcde | 15.0 | 16.2 | 42.5 defghijk | 12.0 | 28.0 | 62.5 abcde | 17.7 | 28.3 |
| Gajo Arroba | ITACYL | 92.5 abcde | 11.9 | 12.9 | 50.0 bcdefghi | 20.4 | 40.8 | 46.9 cdefgh | 15.7 | 33.5 |
| Tortozona Tinta | EVENA + UPNA | 92.5 abcde | 11.9 | 12.9 | 50.0 ijklmn | 20.4 | 40.8 | 68.7 bcdefg | 37.5 | 54.5 |
| Santa Fe/Cadrete | EVENA + UPNA | 92.5 abcde | 15.0 | 16.2 | 27.5 bcdefghi | 5.0 | 18.1 | 56.2 abcd | 23.9 | 42.5 |
| Tinto Fragoso | IVICAM | 92.2 abcde | 11.8 | 12.8 | 26.2 jklmno | 18.4 | 70.2 | 37.5 efghi | 25.0 | 66.7 |
| Albariño Tinto | EVEGA | 91.5 abcde | 13.9 | 15.2 | 10.0 no | 10.0 | 100.0 | 37.5 efghi | 25.0 | 66.7 |
| Trobat Negre | INCAVI | 90.8 abcde | 12.7 | 14.0 | 68.7 ab | 23.9 | 34.8 | 75.0 ab | 0.0 | 0.0 |
| Cagarrizo | CICYTEX | 90.7 abcde | 11.1 | 12.3 | 26.2 jklmno | 18.4 | 70.2 | 50.0 bcdefgh | 0.0 | 0.0 |
| Albilla do Avia | EVEGA | 89.8 abcde | 16.7 | 18.6 | 32.5 ghijklm | 21.8 | 67.1 | 50.0 bcdefgh | 28.9 | 57.7 |
| Rayada Melonera | IFAPA | 88.8 abcde | 22.5 | 25.3 | 65.6 abc | 31.2 | 47.6 | 84.4 a | 18.7 | 22.2 |
| Albarín Tinto | EVEGA | 87.7 abcdef | 14.8 | 16.9 | 46.9 bcdefghij | 15.7 | 33.5 | 53.1 bcdefg | 21.3 | 40.2 |
| Castellana Blanca | EVENA + UPNA | 87.5 abcdef | 14.4 | 16.5 | 37.5 efghijkl | 14.4 | 38.5 | 62.5 abcde | 14.4 | 23.1 |
| Jarrosuelto | EVENA + UPNA | 87.5 abcdef | 14.4 | 16.5 | 40.6 defghijk | 23.9 | 54.7 | 75.0 ab | 12.5 | 18.2 |
| Rufete Serrano | ITACYL | 86.7 abcdef | 18.9 | 21.7 | 37.5 efghijkl | 14.4 | 38.5 | 62.5 abcde | 14.4 | 23.1 |
| Estaladiña | ITACYL | 86.5 abcdef | 27.5 | 31.9 | 37.5 efghijkl | 14.4 | 38.5 | 53.1 bcdefg | 21.3 | 40.2 |
| Diega 2 | EVENA + UPNA | 86.0 abcdef | 16.3 | 19.0 | 28.1 ijklmn | 6.2 | 22.2 | 68.7 abcd | 12.5 | 18.2 |
| Riera 46 | INCAVI | 85.7 abcdef | 16.7 | 19.5 | 32.5 ghijklm | 21.8 | 67.1 | 62.5 abcde | 14.4 | 23.1 |
| Cagarrizo | IMIDRA | 85.2 abcdefg | 20.7 | 24.3 | 36.9 fghijklm | 10.3 | 27.9 | 53.1 bcdefg | 6.2 | 11.8 |
| Arcos | ITVE | 85.0 abcdefg | 30.0 | 35.3 | 25.0 jklmno | 56.6 | 29.8 | 56.2 bcdefg | 22.2 | 59.4 |
| Terriza | IVICAM | 84.2 abcdefg | 11.9 | 14.1 | 56.2 abcdef | 12.5 | 22.2 | 62.5 abcde | 14.4 | 23.1 |
| Planta Nova | ITVE | 83.3 abcdefg | 28.9 | 34.6 | 50.0 bcdefghi | 0.0 | 0.0 | 58.3 bcdef | 28.8 | 49.5 |
| Jarrosuelto | IVICAM | 83.2 abcdefg | 22.2 | 26.7 | 43.7 cdefghijk | 12.0 | 29.5 | 68.7 abcd | 0.0 | 0.0 |
| Sanguina | INCAVI | 83.0 abcdefg | 19.9 | 24.0 | 43.7 cdefghijk | 16.14 | 36.9 | 56.2 bcdefg | 23.9 | 42.5 |
| Tortozona Tinta | IVICAM | 82.2 abcdefg | 12.0 | 14.6 | 37.5 efghijkl | 14.4 | 38.5 | 50.0 bcdefgh | 20.4 | 40.8 |
| Albana | DGA | 81.7 abcdefg | 21.3 | 26.1 | 25.6 jklmno | 9.2 | 36.0 | 50.0 bcdefgh | 20.4 | 40.8 |
| Hebén | IMIDRA | 81.2 abcdefg | 13.5 | 16.2 | 25.6 jklmno | 9.2 | 36.0 | 43.7 dfgh | 12.5 | 28.6 |
| Terriza | IMIDRA | 81.2 abcdefg | 23.9 | 29.5 | 43.7 cdefghijk | 23.9 | 54.7 | 38.7 efghi | 30.34 | 78.4 |
| Maquías | IVICAM | 80.6 abcdefg | 4.1 | 5.1 | 21.9 klmno | 10.3 | 47.0 | 62.5 abcde | 14.4 | 23.1 |
| Xafardán | EVEGA | 79.2 abcdefg | 24.1 | 30.4 | 37.5 efghijkl | 25.0 | 66.7 | 56.2 bcdefg | 37.5 | 66.7 |
| Tinto Jeromo | IVICAM | 77.0 abcdefg | 28.0 | 36.4 | 15.0 mno | 11.5 | 77.0 | 25.0 hi | 0.0 | 0.0 |
| Castellana Blanca | IMIDRA | 76.0 bcdefg | 28.3 | 37.1 | 23.8 klmno | 16.4 | 69.0 | 34.4 fghi | 12.0 | 34.8 |
| Riera 43 | INCAVI | 76.0 bcdefg | 7.4 | 9.8 | 24.4 klmno | 22.9 | 94.1 | 50.0 bcdefgh | 28.9 | 57.7 |
| Sanguina | IVICAM | 72.1 cdefgh | 16.0 | 22.2 | 25.0 hijklmn | 0.0 | 0.0 | 25.0 abcdef | 0.0 | 0.0 |
| Planta Mula | ITVE | 71.5 defgh | 34.6 | 48.4 | 7.5 jklmno | 5.0 | 66.7 | 32.5 hi | 29.9 | 91.9 |
| Bastardo blanco | CICYTEX | 72.2 efgh | 26.9 | 37.7 | 29.8 no | 18.8 | 63.3 | 59.4 ghi | 12.0 | 20.2 |
| Rayada Melonera | IMIDRA | 65.2 fgh | 11.7 | 17.9 | 28.1 ijklmn | 6.2 | 22.2 | 50.0 bcdefgh | 20.4 | 40.8 |
| Zamarrica | EVEGA | 64.5 fgh | 23.1 | 35.7 | 17.5 lmno | 15.0 | 85.7 | 25.0 hi | 0.0 | 0.0 |
| Cariñena Roja | INCAVI | 62.0 hg | 26.9 | 43.4 | 26.2 jklmno | 25.5 | 97.2 | 32.5 ghi | 29.9 | 91.9 |
| Mandrègue | DGA | 51.7 hi | 14.20 | 27.4 | 10.0 no | 10.0 | 100.0 | 43.7 dfgh | 12.5 | 28.6 |
| Morate | IMIDRA | 34.0 ij | 14.28 | 42.0 | 17.5 lmno | 15.0 | 85.7 | 37.5 efghi | 14.4 | 38.5 |
| 110-Richter | MBG | 18.3 jk | 11.25 | 62.1 | 15.0 mno | 0.0 | 0.0 | 15.0 i | 11.5 | 77.0 |
| SO4 | MBG | 5.0 k | 0.00 | 0.0 | 5.0 o | 0.0 | 0.0 | 15.0 i | 11.5 | 77.0 |
| LSD (0.05) | 23.5 | 22.4 | 25.4 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boso, S.; Gago, P.; Santiago, J.-L.; Muñoz-Organero, G.; Cabello, F.; Puertas, B.; Puig, A.; Domingo, C.; Valdés, M.E.; Moreno, D.; et al. Variation in Susceptibility to Downy Mildew Infection in Spanish Minority Vine Varieties. Plants 2023, 12, 2638. https://doi.org/10.3390/plants12142638
Boso S, Gago P, Santiago J-L, Muñoz-Organero G, Cabello F, Puertas B, Puig A, Domingo C, Valdés ME, Moreno D, et al. Variation in Susceptibility to Downy Mildew Infection in Spanish Minority Vine Varieties. Plants. 2023; 12(14):2638. https://doi.org/10.3390/plants12142638
Chicago/Turabian StyleBoso, Susana, Pilar Gago, José-Luis Santiago, Gregorio Muñoz-Organero, Félix Cabello, Belén Puertas, Anna Puig, Carme Domingo, M. Esperanza Valdés, Daniel Moreno, and et al. 2023. "Variation in Susceptibility to Downy Mildew Infection in Spanish Minority Vine Varieties" Plants 12, no. 14: 2638. https://doi.org/10.3390/plants12142638
APA StyleBoso, S., Gago, P., Santiago, J.-L., Muñoz-Organero, G., Cabello, F., Puertas, B., Puig, A., Domingo, C., Valdés, M. E., Moreno, D., Diaz-Losada, E., Cibriain, J. F., Dañobeitia-Artabe, O., Rubio-Cano, J.-A., Martínez-Gascueña, J., Mena-Morales, A., Chirivella, C., Usón, J.-J., & Martínez, M.-C. (2023). Variation in Susceptibility to Downy Mildew Infection in Spanish Minority Vine Varieties. Plants, 12(14), 2638. https://doi.org/10.3390/plants12142638