
Climatic Niche Dynamics of the Astereae Lineage and Haplopappus Species Distribution following Amphitropical Long-Distance Dispersal


Abstract
:1. Introduction
2. Results
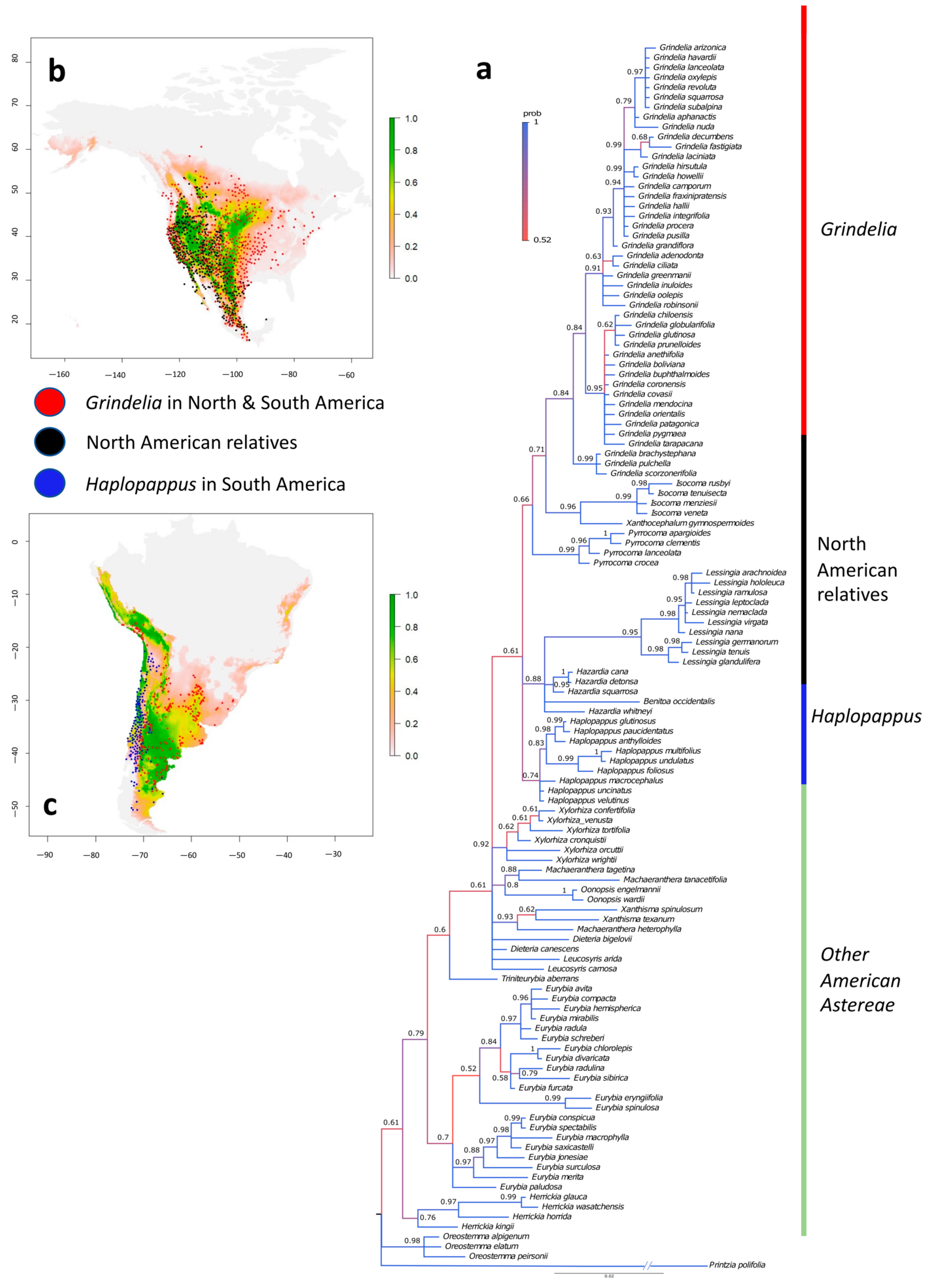
2.1. Identification of Sister Lineages
2.2. Climatic Niche Comparisons
2.3. Richness Structure and Explanatory Variables
3. Discussion
4. Materials and Methods
4.1. Identification of Happlopapus Sister Lineages
4.2. Climate Niche Dynamics
4.3. Richness Structure and Explanatory Variables
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raven, P.H. Amphitropical Relationships in the Floras of North and South America. Q. Rev. Biol. 1963, 38, 151–177. [Google Scholar] [CrossRef]
- Simpson, M.G.; Guilliams, C.M.; Johnson, L.A. Patterns and processes of American amphitropical disjunctions: New insights. Am. J. Bot. 2017, 104, 1597–1599. [Google Scholar] [CrossRef] [Green Version]
- Mooney, H.A.; Dunn, E.L. Convergent Evolution of Mediterranean-Climate Evergreen Sclerophyll Shrubs. Evol. Int. J. Org. Evol. 1970, 24, 292–303. [Google Scholar] [CrossRef]
- Popp, M.; Mirré, V.; Brochmann, C. A single Mid-Pleistocene long-distance dispersal by a bird can explain the extreme bipolar disjunction in crowberries (Empetrum). Proc. Natl. Acad. Sci. USA 2011, 108, 6520–6525. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Muñoz, A. Plant Geography of Chile; Springer: Dordrecht, The Netherlands, 2011; p. 320. [Google Scholar] [CrossRef]
- Pearman, P.B.; Guisan, A.; Broennimann, O.; Randin, C.F. Niche dynamics in space and time. Trends Ecol. Evol. 2008, 23, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Wiens, J.J.; Ackerly, D.D.; Allen, A.P.; Anacker, B.L.; Buckley, L.B.; Cornell, H.V.; Damschen, E.I.; Jonathan Davies, T.; Grytnes, J.A.; Harrison, S.P.; et al. Niche conservatism as an emerging principle in ecology and conservation biology. Ecol. Lett. 2010, 13, 1310–1324. [Google Scholar] [CrossRef] [PubMed]
- Broennimann, O.; Treier, U.A.; Müller-Schärer, H.; Thuiller, W.; Peterson, A.T.; Guisan, A. Evidence of climatic niche shift during biological invasion. Ecol. Lett. 2007, 10, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Crisp, M.D.; Arroyo, M.T.K.; Cook, L.G.; Gandolfo, M.A.; Jordan, G.J.; McGlone, M.S.; Weston, P.H.; Westoby, M.; Wilf, P.; Linder, H.P. Phylogenetic biome conservatism on a global scale. Nature 2009, 458, 754. [Google Scholar] [CrossRef]
- Wiens, J.J. The causes of species richness patterns across space, time, and clades and the role of “ecological limits”. Q. Rev. Biol. 2011, 86, 75–96. [Google Scholar] [CrossRef] [Green Version]
- Pyron, R.A.; Costa, G.C.; Patten, M.A.; Burbrink, F.T. Phylogenetic niche conservatism and the evolutionary basis of ecological speciation. Biol. Rev. 2015, 90, 1248–1262. [Google Scholar] [CrossRef]
- Guerrero, P.C.; Rosas, M.; Arroyoa, M.T.K.; Wiens, J.J. Evolutionary lag times and recent origin of the biota of an ancient desert (Atacama–Sechura). Proc. Natl. Acad. Sci. USA 2013, 110, 11469–11474. [Google Scholar] [CrossRef] [PubMed]
- Wiens, J.J.; Graham, C.H. Niche Conservatism: Integrating Evolution, Ecology, and Conservation Biology. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 519–539. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Jarvie, S.; Guo, W.-Y.; Deng, T.; Mao, L.; Zhang, M.; Chu, C.; Qian, H.; Svenning, J.-C.; He, F. Niche overlap and divergence times support niche conservatism in eastern Asia–eastern North America disjunct plants. Glob. Ecol. Biogeogr. 2021, 30, 1990–2003. [Google Scholar] [CrossRef]
- Heibl, C.; Renner, S.S. Distribution Models and a Dated Phylogeny for Chilean Oxalis Species Reveal Occupation of New Habitats by Different Lineages, not Rapid Adaptive Radiation. Syst. Biol. 2012, 61, 823–834. [Google Scholar] [CrossRef] [PubMed]
- Donoghue, M.J.; Edwards, E.J. Biome Shifts and Niche Evolution in Plants. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 547–572. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Petitpierre, B.; Broennimann, O.; Daehler, C.; Kueffer, C. Unifying niche shift studies: Insights from biological invasions. Trends Ecol. Evol. 2014, 29, 260–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiroga, R.E.; Premoli, A.C.; Fernández, R.J. Niche dynamics in amphitropical desert disjunct plants: Seeking for ecological and species-specific influences. Glob. Ecol. Biogeogr. 2021, 30, 370–383. [Google Scholar] [CrossRef]
- Peña-Gómez, F.T.; Guerrero, P.C.; Bizama, G.; Duarte, M.; Bustamante, R.O. Climatic Niche Conservatism and Biogeographical Non-Equilibrium in Eschscholzia californica (Papaveraceae), an Invasive Plant in the Chilean Mediterranean Region. PLoS ONE 2014, 9, e105025. [Google Scholar] [CrossRef] [Green Version]
- Brouillet, L.; Lowrey, T.K.; Urbatsch, L.; Karaman-Castro, V.; Sancho, G.; Wagstaff, S.; Semple, J. Astereae. In Systematics, Evolution, and Biogeography of Compositae; Funk, V.A., Susanna, A., Stuessy, T.F., Bayer, R.J., Eds.; Smithsonian National Museum of Natural History: Washington, DC, USA, 2009; pp. 589–629. [Google Scholar]
- Nesom, G.; Robinson, H. Tribe Astereae Cass. In The Families and Genera of Vascular Plants. VIII Flowering Plants. Eudicots. Asterales; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; Volume VIII, pp. 284–341. [Google Scholar]
- Klingenberg, L. Monographie der südamerikanischen Gattungen Haplopappus Cass. und Notopappus L. Klingenberg (Asteracea–Astereae). Bibl. Bot. 2007, 157, 351. [Google Scholar]
- Schneider, A.C.; Moore, A.J. Parallel Pleistocene amphitropical disjunctions of a parasitic plant and its host. Am. J. Bot. 2017, 104, 1745–1755. [Google Scholar] [CrossRef] [Green Version]
- Moore, A.J.; Bartoli, A.; Tortosa, R.D.; Baldwin, B.G. Phylogeny, biogeography, and chromosome evolution of the amphitropical genus Grindelia (Asteraceae) inferred from nuclear ribosomal and chloroplast sequence data. Taxon 2012, 61, 211–230. [Google Scholar] [CrossRef]
- Koppen, W. Climatología; Fondo de Cultura Económica: Mexico City, Mexico, 1948; Available online: https://cir.nii.ac.jp/crid/1130282272370970880 (accessed on 10 May 2023).
- Luebert, F.; Pliscoff, P. Sinopsis Bioclimática y Vegetacional de Chile; Universidad de Chile: Santiago, Chile, 2006; p. 384. [Google Scholar] [CrossRef]
- Araújo, M.B.; Pearson, R.G. Equilibrium of species’ distributions with climate. Ecography 2005, 28, 693–695. [Google Scholar] [CrossRef]
- Rundel, P.W.; Dillon, M.O.; Palma, B.; Mooney, H.A.; Gulmon, S.L.; Ehleringer, J.R. The phytogeography and ecology of the coastal Atacama and Peruvian Deserts. Aliso 1991, 13, 1–49. [Google Scholar] [CrossRef] [Green Version]
- Garreaud, R.D.; Vuille, M.; Compagnucci, R.; Marengo, J. Present-day South American climate. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 281, 180–195. [Google Scholar] [CrossRef]
- Constance, L. Introduction and Historical Review. Amphitropical relationships in the herbaceous flora of the pacific coast of north and south america: A symposium. Q. Rev. Biol. 1963, 38, 109–116. [Google Scholar] [CrossRef]
- Moore, M.J.; Jansen, R.K. Molecular evidence for the age, origin, and evolutionary history of the American desert plant genus Tiquilia (Boraginaceae). Mol. Phylogenetics Evol. 2006, 39, 668–687. [Google Scholar] [CrossRef]
- Wen, J.; Lowry, P.P.; Walck, J.L.; Yoo, K.-O. Phylogenetic and Biogeographic Diversification in Osmorhiza (Apiaceae). Ann. Mo. Bot. Gard. 2002, 89, 414–428. [Google Scholar] [CrossRef] [Green Version]
- Scherson, R.A.; Vidal, R.; Sanderson, M.J. Phylogeny, biogeography, and rates of diversification of New World Astragalus (Leguminosae) with an emphasis on South American radiations. Am. J. Bot. 2008, 95, 1030–1039. [Google Scholar] [CrossRef]
- Ickert-Bond, S.M.; Rydin, C.; Renner, S.S. A fossil-calibrated relaxed clock for Ephedra indicates an Oligocene age for the divergence of Asian and New World clades and Miocene dispersal into South America. J. Syst. Evol. 2009, 47, 444–456. [Google Scholar] [CrossRef] [Green Version]
- Simpson, B.B.; Tate, J.A.; Weeks, A. The biogeography of Hoffmannseggia (Leguminosae, Caesalpinioideae, Caesalpinieae): A tale of many travels. J. Biogeogr. 2005, 32, 15–27. [Google Scholar] [CrossRef]
- Lia, V.V.; Confalonieri, V.A.; Comas, C.I.; Hunziker, J.H. Molecular Phylogeny of Larrea and Its Allies (Zygophyllaceae): Reticulate Evolution and the Probable Time of Creosote Bush Arrival to North America. Mol. Phylogenetics Evol. 2001, 21, 309–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, J.; Ickert-Bond, S.M. Evolution of the Madrean–Tethyan disjunctions and the North and South American amphitropical disjunctions in plants. J. Syst. Evol. 2009, 47, 331–348. [Google Scholar] [CrossRef]
- Hughes, C.; Eastwood, R. Island radiation on a continental scale: Exceptional rates of plant diversification after uplift of the Andes. Proc. Natl. Acad. Sci. USA 2006, 103, 10334–10339. [Google Scholar] [CrossRef] [PubMed]
- Villaverde, T.; González-Moreno, P.; Rodríguez-Sánchez, F.; Escudero, M. Niche shifts after long-distance dispersal events in bipolar sedges (Carex, Cyperaceae). Am. J. Bot. 2017, 104, 1765–1774. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, P.C.; Durán, A.P.; Walter, H.E. Latitudinal and altitudinal patterns of the endemic cacti from the Atacama desert to Mediterranean Chile. J. Arid. Environ. 2011, 75, 991–997. [Google Scholar] [CrossRef]
- Jara-Arancio, P.; Arroyo, M.T.K.; Guerrero, P.C.; Hinojosa, L.F.; Arancio, G.; Méndez, M.A. Phylogenetic perspectives on biome shifts in Leucocoryne (Alliaceae) in relation to climatic niche evolution in western South America. J. Biogeogr. 2014, 41, 328–338. [Google Scholar] [CrossRef]
- Pulliam, H.R. Sources, Sinks, and Population Regulation. Am. Nat. 1988, 132, 652–661. [Google Scholar] [CrossRef]
- Ulrich, W.; Almeida-Neto, M.; Gotelli, N.J. A consumer’s guide to nestedness analysis. Oikos 2009, 118, 3–17. [Google Scholar] [CrossRef]
- Díaz, F.P.; Latorre, C.; Carrasco-Puga, G.; Wood, J.R.; Wilmshurst, J.M.; Soto, D.C.; Cole, T.L.; Gutiérrez, R.A. Multiscale climate change impacts on plant diversity in the Atacama Desert. Glob. Chang. Biol. 2019, 25, 1733–1745. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information; National Library of Medicine (US); National Center for Biotechnology Information. 2020. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 10 May 2023).
- Xia, X. DAMBE7: New and Improved Tools for Data Analysis in Molecular Biology and Evolution. Mol. Biol. Evol. 2018, 35, 1550–1552. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. raxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol. Evol. 2021, 12, 373–377. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree v1.4.4; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2016; Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 14 May 2023).
- GBIF.org GBIF. Available online: https://www.gbif.org/occurrence/download/0084489-230530130749713 (accessed on 14 May 2023).
- GBIF.org GBIF. Available online: https://www.gbif.org/occurrence/download/0084505-230530130749713 (accessed on 14 May 2023).
- GBIF.org GBIF. Available online: https://www.gbif.org/occurrence/download/0084478-230530130749713 (accessed on 14 May 2023).
- GBIF.org GBIF. Available online: https://www.gbif.org/occurrence/download/0084408-230530130749713 (accessed on 14 May 2023).
- GBIF.org GBIF. Available online: https://www.gbif.org/occurrence/download/0084485-230530130749713 (accessed on 14 May 2023).
- GBIF.org GBIF. Available online: https://www.gbif.org/occurrence/download/0084496-230530130749713 (accessed on 14 May 2023).
- GBIF.org GBIF. Available online: https://www.gbif.org/occurrence/download/0084498-230530130749713 (accessed on 14 May 2023).
- Chamberlain, S.; Barve, V.; Mcglinn, D.; Oldoni, D.; Desmet, P.; Geffert, L.; Ram, K. rgbif: Interface to the Global Biodiversity Information Facility API. R Package Version 3.7.7. 2023. Available online: https://CRAN.R-project.org/package=rgbif (accessed on 14 May 2023).
- Zizka, A.; Silvestro, D.; Andermann, T.; Azevedo, J.; Duarte Ritter, C.; Edler, D.; Farooq, H.; Herdean, A.; Ariza, M.; Scharn, R.; et al. CoordinateCleaner: Standardized cleaning of occurrence records from biological collection databases. Methods Ecol. Evol. 2019, 10, 744–751. [Google Scholar] [CrossRef]
- Chapman, A.D. Principles and Methods of Data Cleaning—Primary Species and Species Occurrence Data, Version 1.0.; GBIF: Copenhagen, Danmark, 2005. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- R: A Language and Environment for Statistical Computing. Available online: http://www.Rproject.org/ (accessed on 10 January 2022).
- Di Cola, V.; Broennimann, O.; Petitpierre, B.; Breiner, F.T.; D’Amen, M.; Randin, C.; Engler, R.; Pottier, J.; Pio, D.; Dubuis, A.; et al. ecospat: An R package to support spatial analyses and modeling of species niches and distributions. Ecography 2017, 40, 774–787. [Google Scholar] [CrossRef]
- Broennimann, O.; Fitzpatrick, M.C.; Pearman, P.B.; Petitpierre, B.; Pellissier, L.; Yoccoz, N.G.; Thuiller, W.; Fortin, M.-J.; Randin, C.; Zimmermann, N.E.; et al. Measuring ecological niche overlap from occurrence and spatial environmental data. Glob. Ecol. Biogeogr. 2012, 21, 481–497. [Google Scholar] [CrossRef] [Green Version]
- Broennimann, O.; Mráz, P.; Petitpierre, B.; Guisan, A.; Müller-Schärer, H. Contrasting spatio-temporal climatic niche dynamics during the eastern and western invasions of spotted knapweed in North America. J. Biogeogr. 2014, 41, 1126–1136. [Google Scholar] [CrossRef] [Green Version]
- Schoener, T.W. The Anolis Lizards of Bimini: Resource Partitioning in a Complex Fauna. Ecology 1968, 49, 704–726. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evol. Int. J. Org. Evol. 2008, 62, 2868–2883. [Google Scholar] [CrossRef] [PubMed]
- Petitpierre, B.; Kueffer, C.; Broennimann, O.; Randin, C.; Daehler, C.; Guisan, A. Climatic niche shifts are rare among terrestrial plant invaders. Science 2012, 335, 1344–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippi, R.A. Viage al Desierto de Atacama Hecho de Orden del Gobierno de Chile en el Verano de 1853–1854. 1860. Available online: http://www.memoriachilena.gob.cl/602/w3-article-7825.html (accessed on 10 May 2023).
- Muñoz-Chick, M.P.; Aníbal, O. Nota aclaratoria sobre especies colectadas por Federico Philippi en su viaje a Tarapaca. Not. Mens. Mus. Nac. Hist. Nat. Chile 1987, 313, 2–17. [Google Scholar]
- Jiles, C. Field Notebook. CONC Herbarium: 1947–1981. Available online: https://sites.google.com/a/uv.cl/herbarium/qf-carlos-jiles-pizarro (accessed on 10 January 2023).
- McCain, C.M. The mid-domain effect applied to elevational gradients: Species richness of small mammals in Costa Rica. J. Biogeogr. 2004, 31, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Colwell, R.K.; Hurtt, G.C. Nonbiological Gradients in Species Richness and a Spurious Rapoport Effect. Am. Nat. 1994, 144, 570–595. [Google Scholar] [CrossRef]
- Clarke, K.R.; Somerfield, P.J.; Gorley, R.N. Testing of null hypotheses in exploratory community analyses: Similarity profiles and biota-environment linkage. J. Exp. Mar. Biol. Ecol. 2008, 366, 56–69. [Google Scholar] [CrossRef]
- Ulrich, W.; Gotelli, N.J. Null model analysis of species nestedness patterns. Ecology 2007, 88, 1824–1831. [Google Scholar] [CrossRef]
- Almeida-Neto, M.; Guimarães, P.; Guimarães, P.R., Jr.; Loyola, R.D.; Ulrich, W. A consistent metric for nestedness analysis in ecological systems: Reconciling concept and measurement. Oikos 2008, 117, 1227–1239. [Google Scholar] [CrossRef]
- Gotelli, N.J. Null model analysis of species co-occurrence patterns. Ecology 2000, 81, 2606–2621. [Google Scholar] [CrossRef]
- Almeida-Neto, M.; Ulrich, W. A straightforward computational approach for measuring nestedness using quantitative matrices. Environ. Model. Softw. 2011, 26, 173–178. [Google Scholar] [CrossRef]
- Strona, G.; Galli, P.; Seveso, D.; Montano, S.; Fattorini, S. Nestedness for Dummies (NeD): A User-Friendly Web Interface for Exploratory Nestedness Analysis. J. Stat. Softw. 2014, 59, 9. [Google Scholar] [CrossRef] [Green Version]
- Pagel, M.; Meade, A.; Barker, D. Bayesian Estimation of Ancestral Character States on Phylogenies. Syst. Biol. 2004, 53, 673–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Software Bayes Trees V.1.3. 2009. Available online: http://www.evolution.reading.ac.uk/BayesTrees (accessed on 14 May 2023).
- Rahbek, C.; Gotelli, N.J.; Colwell, R.K.; Entsminger, G.L.; Rangel, T.F.L.V.B.; Graves, G.R. Predicting continental-scale patterns of bird species richness with spatially explicit models. Proc. R. Soc. B Biol. Sci. 2007, 274, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Rangel, T.F.; Diniz-Filho, J.A.F.; Bini, L.M. SAM: A comprehensive application for Spatial Analysis in Macroecology. Ecography 2010, 33, 46–50. [Google Scholar] [CrossRef]
- DIVA-GIS v. 4. 2004. Available online: https://www.diva-gis.org/gdata (accessed on 14 May 2023).




{kind=link}
{kind=link}
{kind=link}
| Locus | Total Length | Ingroup Variable Characters | Total Variable Characters | Parsimony Informative Characters | Ingroup Coverage (%) | Outgroup Coverage (%) | % Variability |
|---|---|---|---|---|---|---|---|
| ETS | 539 | 128 | 128 | 63 | 1 | 98 | 35 |
| ITS1&2 | 645 | 94 | 209 | 123 | 1 | 99 | 57 |
| Concat. Matrix | 1184 | 337 | 367 | 197 |
| Variables | SAR Coeff. |
|---|---|
| Mean Diurnal Range (Bio2) | 0.465 |
| Isothermality (Bio3) | 0.156 |
| Temperature Seasonality (Bio4) | 0.036 * |
| Temperature Annual Range (Bio7) | 0.852 |
| Annual Mean Temperature (Bio1) | 0.192 |
| Mean Temperature of Wettest Quarter (Bio8) | 0.181 * |
| Annual Precipitation (Bio12) | 0.001 |
| R2 | 0.469 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosas, M.R.; Segovia, R.A.; Guerrero, P.C. Climatic Niche Dynamics of the Astereae Lineage and Haplopappus Species Distribution following Amphitropical Long-Distance Dispersal. Plants 2023, 12, 2721. https://doi.org/10.3390/plants12142721
Rosas MR, Segovia RA, Guerrero PC. Climatic Niche Dynamics of the Astereae Lineage and Haplopappus Species Distribution following Amphitropical Long-Distance Dispersal. Plants. 2023; 12(14):2721. https://doi.org/10.3390/plants12142721
Chicago/Turabian StyleRosas, Marcelo R., Ricardo A. Segovia, and Pablo C. Guerrero. 2023. "Climatic Niche Dynamics of the Astereae Lineage and Haplopappus Species Distribution following Amphitropical Long-Distance Dispersal" Plants 12, no. 14: 2721. https://doi.org/10.3390/plants12142721
APA StyleRosas, M. R., Segovia, R. A., & Guerrero, P. C. (2023). Climatic Niche Dynamics of the Astereae Lineage and Haplopappus Species Distribution following Amphitropical Long-Distance Dispersal. Plants, 12(14), 2721. https://doi.org/10.3390/plants12142721