

Genome-Wide Identification, Phylogeny and Expression Analysis of Subtilisin (SBT) Gene Family under Wheat Biotic and Abiotic Stress


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification of Wheat SBT Protein Family
2.2. Phylogenetic Analysis of Wheat SBTs
2.3. Collinearity and Chromosome Analysis of Wheat SBT Genes
2.4. Gene structure Characterization and Protein Motif Analysis
2.5. Identification of Cis-Acting Elements in TaSBTs Promoter Region
2.6. Expression Pattern of SBT Genes in Wheat
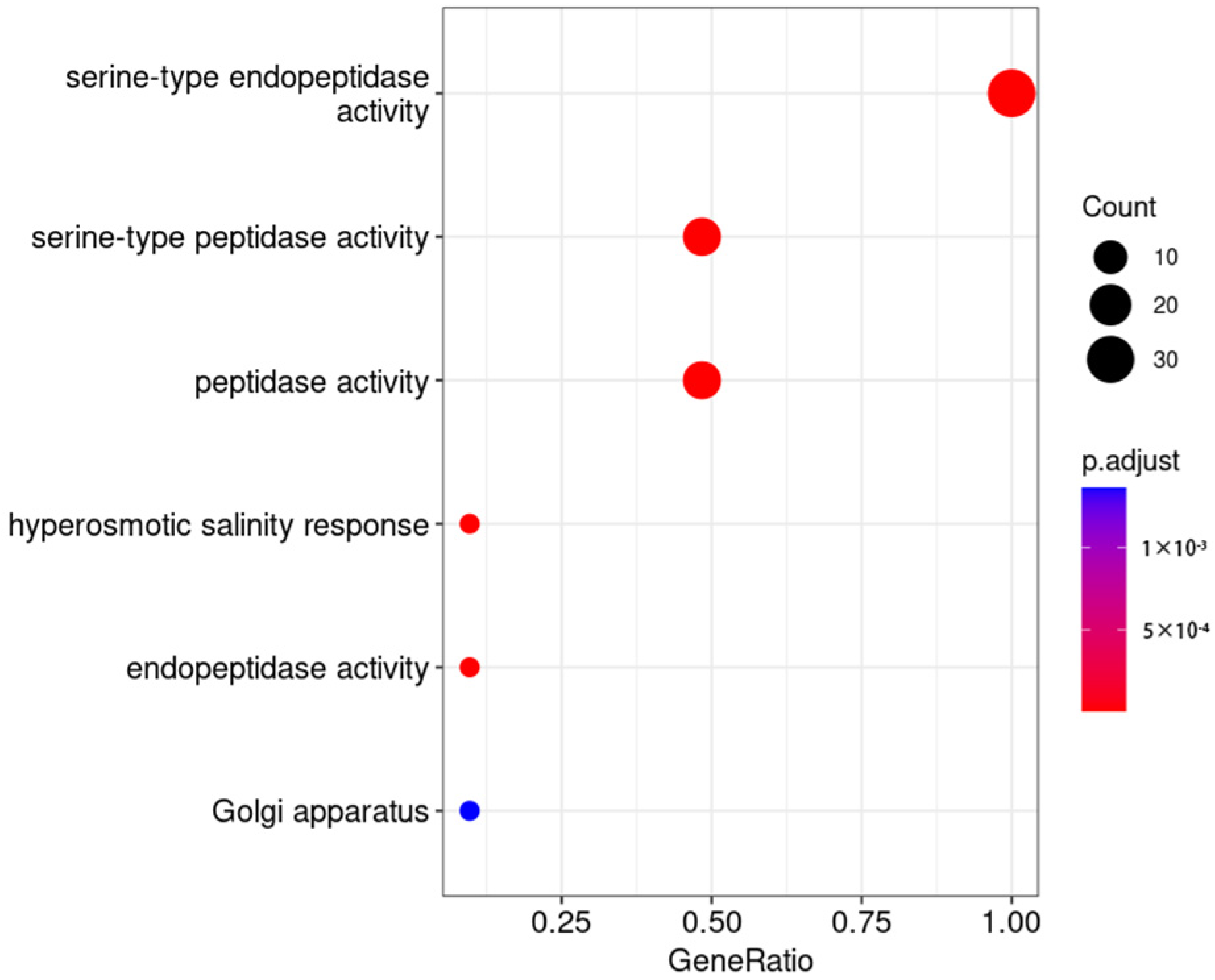
2.7. GO Enrichment Analysis of Differentially Expressed Genes
2.8. qRT-PCR Expression Analysis of TaSBTs during Pst Interaction
3. Materials and Methods
3.1. RNA Extraction, Stripe Rust Inoculation and Plant Materials
3.2. Identification and Classification of Wheat TaSBT Gene Family
3.3. Structure, and Phylogenetic Analysis of Wheat SBT Proteins
3.4. Analysis of TaSBTs Gene Expression Profile and Cis-Acting Elements
3.5. Expression Analysis of TaSBTs under Pst-Wheat Interaction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yi, J.P.; He, J.; Shan, C.J. Principles and Diagnosis and Treatment of Common Plant Diseases; China Agricultural University Press: Beijing, China, 2012. [Google Scholar]
- Yuan, L.Y.; Xie, Y.J.; Ye, H.G. The occurrence and control of wheat stripe rust. Mod. Agric. Sci. Technol. 2012, 6, 167–168. [Google Scholar] [CrossRef]
- Havé, M.; Balliau, T.; Cottyn-Boitte, B.; Dérond, E.; Cueff, G.; Soulay, F.; Lornac, A.; Reichman, P.; Dissmeyer, N.; Avice, J.C.; et al. Increase of proteasome and papain-like cysteine protease activities in autophagy mutants: Backup compensatory effect or pro cell-death effect? Exp. Bot. 2017, 69, 1369–1385. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.J.; Tan, R.; Huang, M.Y.; Du, J.; Yu, Y.; Yang, Y.H.; Bi, C.W. Research progress on the interaction between plants and pathogens using subtilisin like proteases. Mol. Plant Breed. 2018, 16, 8. [Google Scholar]
- Schaller, A.; Stintzi, A.; Rivas, S.; Serrano, I.; Vera, P. From structure to function—A family portrait of plant subtilases. New Phytol. 2017, 218, 901–915. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.H. The biological function of Bacillus subtilis like protease and its application in disease control in agriculture and animal husbandry. Anhui Agric. Sci. 2011, 39, 3. [Google Scholar]
- Yamagata, H.; Masuzawa, T.; Nagaoka, Y.; Ohnishi, T.; Iwasaki, T. Cucumisin, a serine protease from melon fruits, shares structural homology with subtilisin and is generated from a large precursor. Biol. Chem. 1994, 269, 32725. [Google Scholar] [CrossRef]
- Tornero, P.; Conejero, V.; Vera, P. Primary structure and expression of a pathogen-induced protease (PR-P69) in tomato plants: Similarity of functional domains to subtilisin-like endoproteases. Proc. Natl. Acad. Sci. USA 1996, 93, 6332–6337. [Google Scholar] [CrossRef]
- Cao, J.; Han, X.; Zhang, T.; Yang, Y.; Huang, J.; Hu, X. Genome-wide and molecular evolution analysis of the subtilase gene family in Vitis vinifera. BMC Genom. 2014, 15, 1116. [Google Scholar] [CrossRef]
- Ekchaweng, K.; Khunjan, U.; Churngchow, N. Molecular cloning and characterization of three novel subtilisin-like serine protease genes from Hevea brasiliensis. Physiol. Mol. Plant Pathol. 2017, 97, 79–95. [Google Scholar] [CrossRef]
- Tripathi, L.P.; Sowdhamini, R. Cross genome comparisons of serine proteases in Arabidopsis and rice. BMC Genom. 2006, 7, 200. [Google Scholar] [CrossRef]
- Schaller, A.; Stintzi, A.; Graff, L. Subtilases—Versatile tools for protein turnover, plant development, and interactions with the environment. Physiol. Plant 2012, 145, 52–66. [Google Scholar] [CrossRef]
- Dai, M.; Zhou, N.; Zhang, Y.; Zhang, Y.; Ni, K.; Wu, Z.; Liu, L.; Wang, X.; Chen, Q. Genome-wide analysis of the SBT gene family involved in drought tolerance in cotton. Front. Plant Sci. 2023, 13, 1097732. [Google Scholar] [CrossRef]
- Neuteboom, L.W.; Veth-Tello, L.M.; Clijdesdale, O.R.; Hooykaas, P.J.J.; van der Zaal, B.J. A Novel Subtilisin-like Protease Gene from Arabidopsis thaliana is Expressed at Sites of Lateral Root Emergence. DNA Res. 1999, 26, 13. [Google Scholar] [CrossRef]
- Zhao, C. Exploiting Secondary Growth in Arabidopsis. Construction of Xylem and Bark cDNA Libraries and Cloning of Three Xylem Endopeptidases. Plant Physiol. 2000, 123, 1185–1196. [Google Scholar] [CrossRef]
- Liu, P.L.; Chen, G.Q.; Shen, X.H. Cloning and Expression Characteristics Analysis of the ApSBT Gene of Bacillus subtilis Protease from Baizilian. Plant Physiol. 2020, 56, 2725–2735. [Google Scholar] [CrossRef]
- Ramírez, V.; López, A.; Mauch-Mani, B.; Gil, M.; Vera, P. An extracellular subtilase switch for immune priming in Arabidopsis. PLoS Pathog. 2013, 9, e1003445. [Google Scholar] [CrossRef]
- Bai, X.X.; Zhan, G.M.; Tian, S.X.; Peng, H.; Cui, X.Y.; Islam, M.A.; Farhan, G.; Ma, Y.Z.; Kang, Z.S.; Xu, Z.S. Transcription factor BZR2 activates chitinase Cht20. 2 transcription to confer resistance to wheat stripe rust. Plant Physiol. 2021, 187, 2749–2762. [Google Scholar] [CrossRef]
- Bolser, D.M.; Kerhornou, A.; Walts, B.; Kersey, P. Editor’s Choice: Triticeae Resources in Ensembl Plants. Plant Cell Physiol. 2015, 56, e3. [Google Scholar] [CrossRef]
- Finn, R.D.; Tate, J.; Mistry, J.; Coggill, P.C.; Sammut, S.J.J.; Hotz, H.R.; Ceric, G.; Forslund, K.; Eddy, S.R.; Sonnhammer, E.L. The Pfam protein families database. Nucleic Acids Res. 2008, 32, D138. [Google Scholar] [CrossRef]
- Wen, S.; Chen, G.; Huang, T.; Shen, X. Genome-wide Study of SBT Genes in Eight Plant Species: Evolution and Expression Outlines for Development and Stress. Res. Sq. 2020. preprint. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef]
- Aron, M.B.; Lu, S.; Anderson, J.B.; Farideh, C.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.F.; Geer, R.C.; Gonzales, N.R. CDD: A Conserved Domain Database for the functional annotation of proteins. Nucleic Acids Res. 2011, 39, D225–D229. [Google Scholar]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Hochstrasser, D.F. Protein Identification and Analysis Tools in the ExPASy Server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar]
- Horton, P.; Park, K.J.; Obayashi, T.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein Localization Prediction Software. Nucleic Acids Res. 2007, 35, 585–587. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Granziol, D.; Ru, B.; Zohren, S.; Dong, X.; Roberts, S. MEMe: An Accurate Maximum Entropy Method for Efficient Approximations in Large-Scale Machine Learning. Entropy 2019, 21, 551. [Google Scholar] [CrossRef]
- Sudhir, K.; Glen, S.; Koichiro, T. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar]
- Guo, B.J.; Li, Y.; Yuan, Z.C.; Lu, C.; Zhang, X.Z.; Xu, R.G. Genome wide analysis of barley ARF gene family. Wheat Crops 2016, 36, 7. [Google Scholar]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL): An online tool for phylogenetic tree display and annotation. FEBS Lett. 1988, 232, 78–82. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.J.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Ramírez-González, R.; Borrill, P.; Lang, D.; Harrington, S.; Brinton, J.; Venturini, L.; Davey, M.; Jacobs, J.; Van, E.F.; Pasha, A.; et al. The transcriptional landscape of polyploid wheat. Science 2018, 361, eaar6089. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Dominguez, R.; Holmes, K.C. Actin structure and function. Annu. Rev. Biophys. 2011, 40, 169–186. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Thomas, D.S. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Rautengarten, C.; Steinhauser, D.; Büssis, D.; Stintzi, A.; Schaller, A.; Kopka, J.; Altmann, T. Inferring Hypotheses on Functional Relationships of Genes: Analysis of the Arabidopsis thaliana Subtilase Gene Family. PLoS Comput. Biol. 2005, 1, e40. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Ellendorff, U.; Kemp, B.; Mansfield, J.W.; Forsyth, A.; Mitchell, K.; Bastas, K.; Liu, C.M.; Woods-Tör, A.; Zipfel, C.; et al. A genome-wide functional investigation into the roles of receptor-like proteins in Arabidopsis. Plant Physiol. 2008, 147, 503–517. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Goher, F.; Chen, L.; Song, J.; Zhao, J. Genome-Wide Identification, Phylogeny and Expression Analysis of Subtilisin (SBT) Gene Family under Wheat Biotic and Abiotic Stress. Plants 2023, 12, 3065. https://doi.org/10.3390/plants12173065
Zhao X, Goher F, Chen L, Song J, Zhao J. Genome-Wide Identification, Phylogeny and Expression Analysis of Subtilisin (SBT) Gene Family under Wheat Biotic and Abiotic Stress. Plants. 2023; 12(17):3065. https://doi.org/10.3390/plants12173065
Chicago/Turabian StyleZhao, Xiaotong, Farhan Goher, Lei Chen, Jiancheng Song, and Jiqiang Zhao. 2023. "Genome-Wide Identification, Phylogeny and Expression Analysis of Subtilisin (SBT) Gene Family under Wheat Biotic and Abiotic Stress" Plants 12, no. 17: 3065. https://doi.org/10.3390/plants12173065