
MeGLYI-13, a Glyoxalase I Gene in Cassava, Enhances the Tolerance of Yeast and Arabidopsis to Zinc and Copper Stresses

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. MeGLYI-13 Gene Sequence Analysis
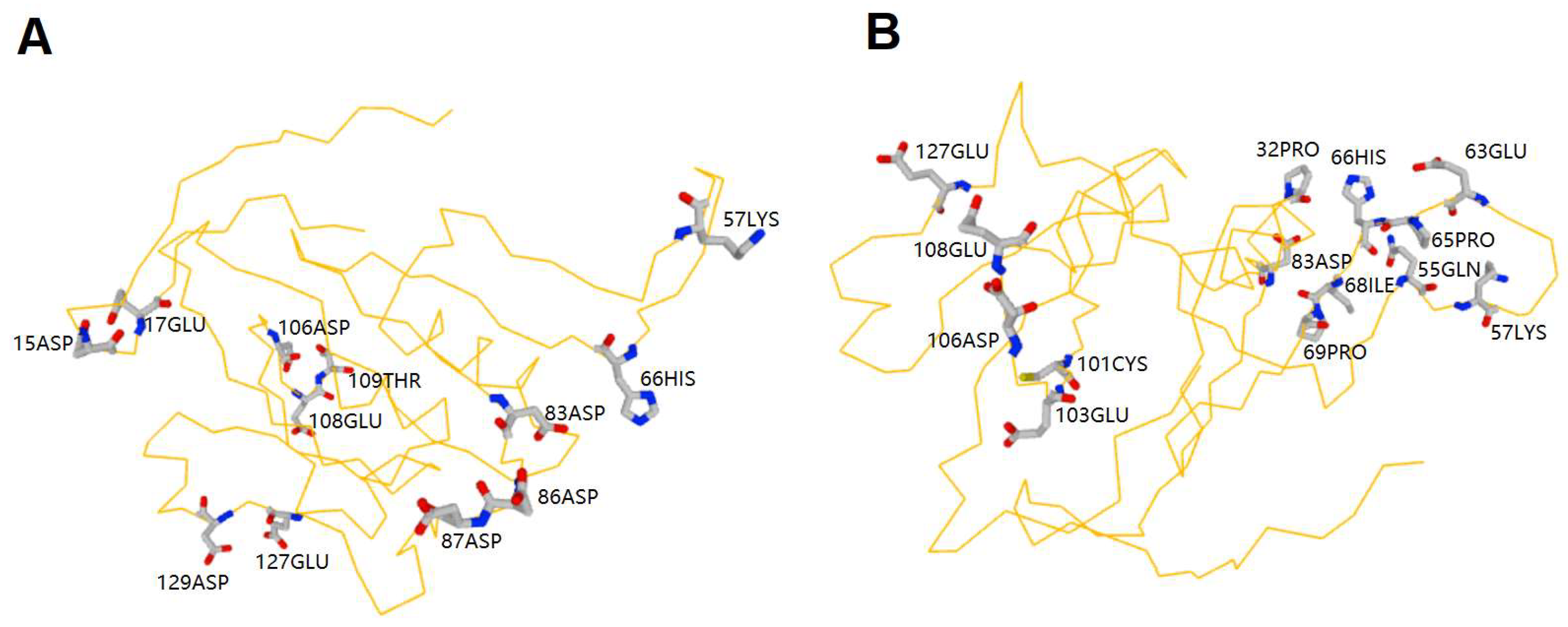
2.2. Prediction of Zn2+ and Cu2+ Binding Sites in MeGLYI-13 Protein Sequence
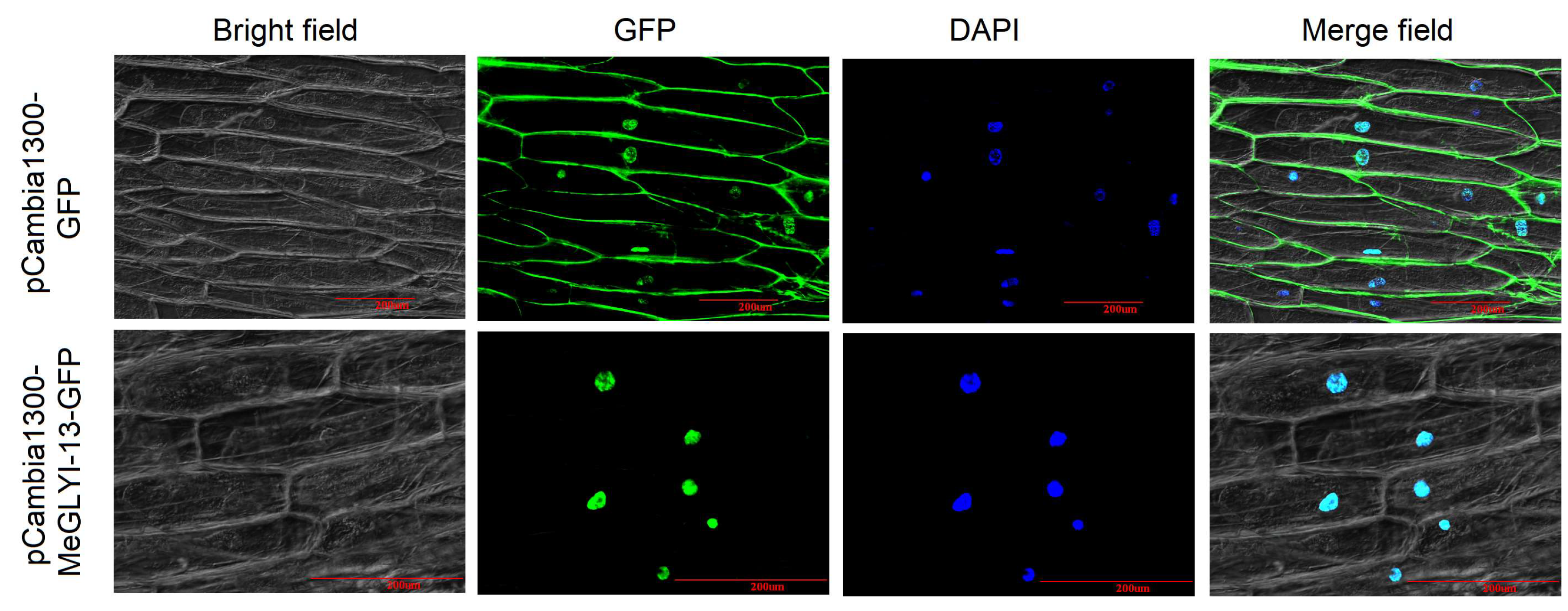
2.3. MeGLYI-13 Protein Was Localized in the Nucleus
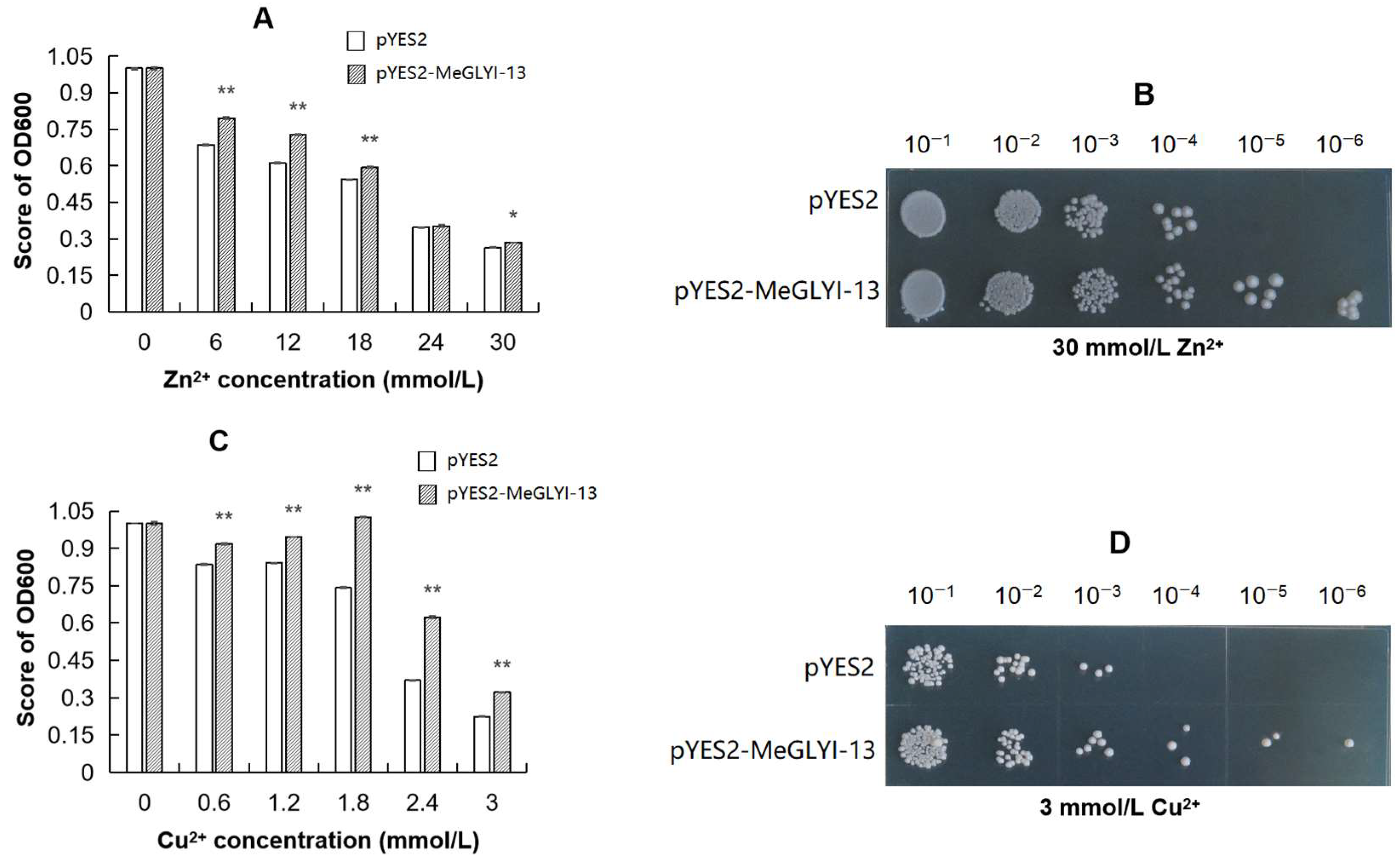
2.4. The Ability of Yeast to Tolerate Cu2+ and Zn2+ Toxicity Is Enhanced by MeGLYI-13
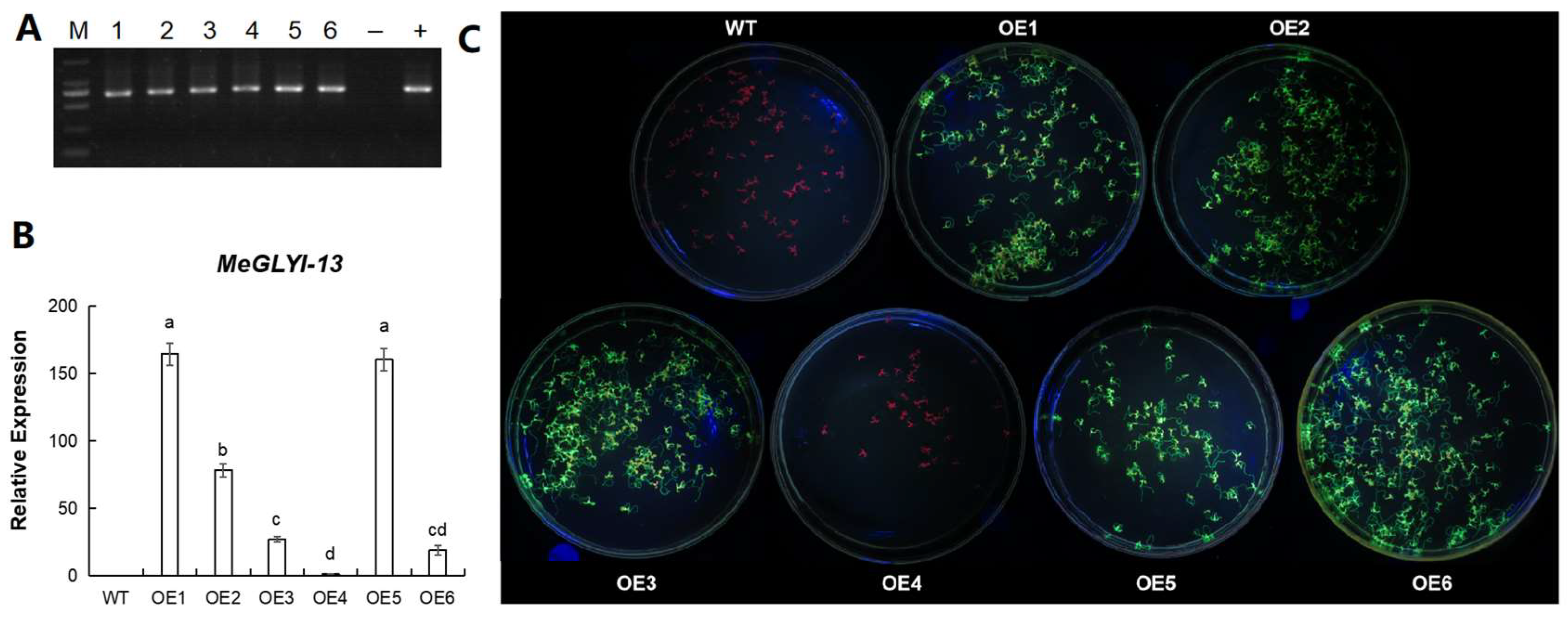
2.5. Expression of MeGLYI-13 in Arabidopsis
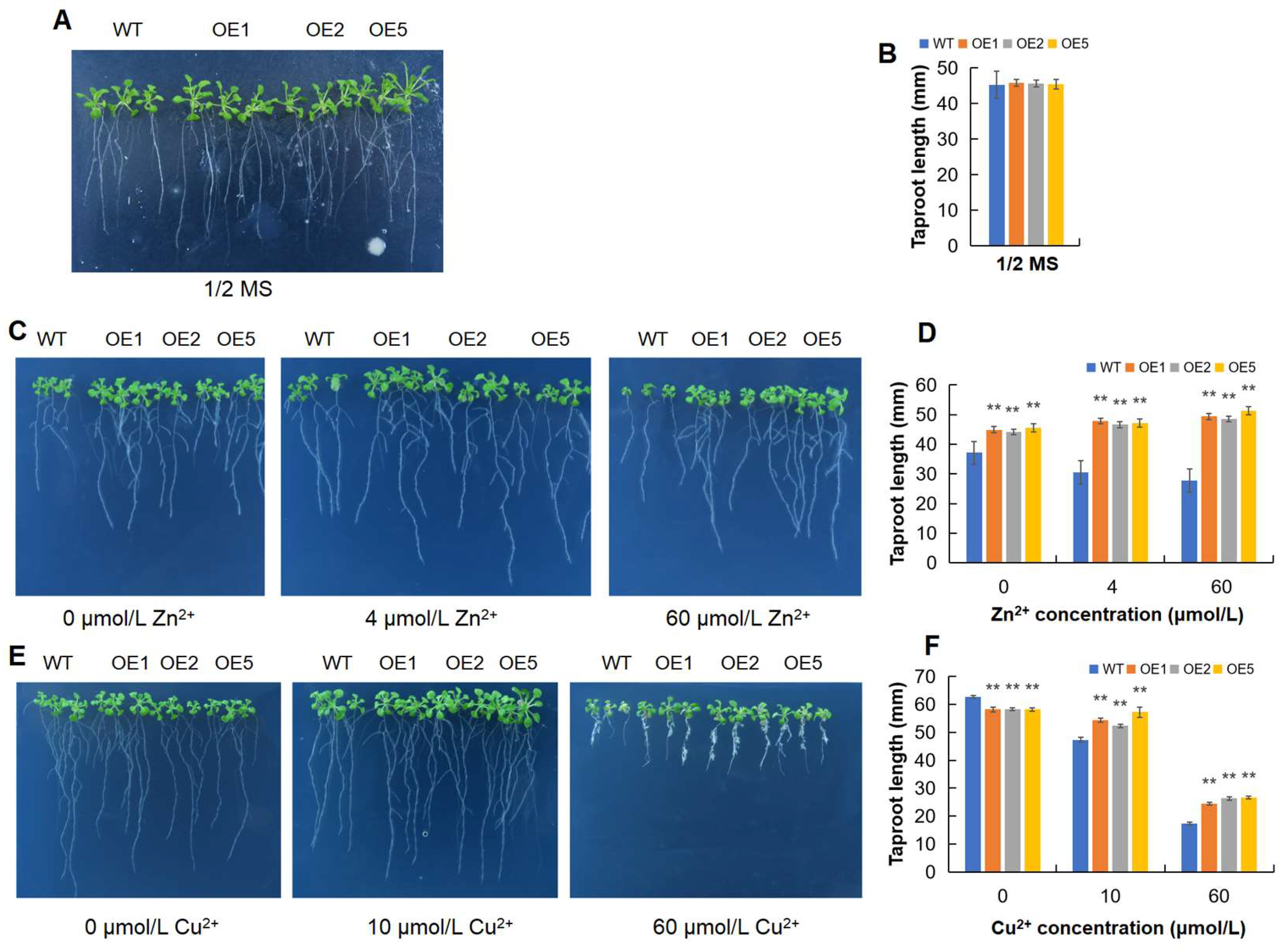
2.6. MeGLYI-13-Overexpressing Arabidopsis Showed Better Tolerance to Zinc Shortage and Toxity
2.7. MeGLYI-13-Overexpressing Arabidopsis Showed Better Tolerance to Copper Toxity
3. Discussion
4. Materials and Methods
4.1. MeGLYI-13 Gene Analysis
4.2. Subcellular Localization of MeGLYI-13
4.3. Expression Analysis of MeGLYI-13 in Response to Zinc and Copper Stress
4.4. Yeast Transformation and Stress Treatments
4.5. Transformation of Arabidopsis with MeGLYI-13
4.6. Detection of Transgenic Arabidopsis
4.7. Expression Level Detection of MeGLYI-13 in Transgenic Arabidopsis
4.8. Fluorescence Detection of Transgenic Arabidopsis
4.9. Znic and Copper Stress Treatment of Transgenic Arabidopsis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ruffing, A.M.; Anthony, S.M.; Strickland, L.M.; Lubkin, I.; Dietz, C.R. Identification of Metal Stresses in Arabidopsis thaliana Using Hyperspectral Reflectance Imaging. Front. Plant Sci. 2021, 12, 624656. [Google Scholar] [CrossRef]
- Alsafran, M.; Saleem, M.H.; Rizwan, M.; Al Jabri, H.; Usman, K.; Fahad, S. An overview of heavy metals toxicity in plants, tolerance mechanism, and alleviation through lysine-chelation with micro-nutrients—A novel approach. Plant Growth Regul. 2023, 100, 337–354. [Google Scholar] [CrossRef]
- Rehman, M.; Liu, L.J.; Wang, Q.; Saleem, M.H.; Bashir, S.; Ullah, S.; Peng, D.X. Copper environmental toxicology, recent advances, and future outlook: A review. Environ. Sci. Pollut. Res. 2019, 26, 18003–18016. [Google Scholar] [CrossRef]
- Behtash, F.; Abedini, F.; Ahmadi, H.; Mosavi, S.B.; Aghaee, A.; Morshedloo, M.R.; Lorenzo, J.M. Zinc Application Mitigates Copper Toxicity by Regulating Cu Uptake, Activity of Antioxidant Enzymes, and Improving Physiological Characteristics in Summer Squash. Antioxidants 2022, 11, 1688. [Google Scholar] [CrossRef]
- Saleem, M.H.; Fahad, S.; Adnan, M.; Ali, M.; Rana, M.S.; Kamran, M.; Ali, Q.; Hashem, I.A.; Bhantana, P.; Ali, M.; et al. Foliar application of gibberellic acid endorsed phytoextraction of copper and alleviates oxidative stress in jute (Corchorus capsularis L.) plant grown in highly copper-contaminated soil of China. Environ. Sci. Pollut. Res. 2020, 27, 37121–37133. [Google Scholar] [CrossRef]
- Liao, F.; Lilay, G.H.; Castro, P.H.; Azevedo, H.; Assuncao, A.G.L. Regulation of the Zinc Deficiency Response in the Legume Model Medicago truncatula. Front. Plant Sci. 2022, 13, 916168. [Google Scholar] [CrossRef]
- Stanton, C.; Sanders, D.; Kramer, U.; Podar, D. Zinc in plants: Integrating homeostasis and biofortification. Mol. Plant 2022, 15, 65–85. [Google Scholar] [CrossRef]
- Cabot, C.; Martos, S.; Llugany, M.; Gallego, B.; Tolra, R.; Poschenrieder, C. A Role for Zinc in Plant Defense against Pathogens and Herbivores. Front. Plant Sci. 2019, 10, 1171. [Google Scholar] [CrossRef]
- Paradisone, V.; Navarro-Leon, E.; Albacete, A.; Ruiz, J.M.M.; Esposito, S.; Blasco, B. Improvement of the physiological response of barley plants to both Zinc deficiency and toxicity by the application of calcium silicate. Plant Sci. 2022, 319, 111259. [Google Scholar] [CrossRef]
- Lu, X.; Liu, S.; Zhi, S.; Chen, J.; Ye, G. Comparative transcriptome profile analysis of rice varieties with different tolerance to zinc deficiency. Plant Biol. 2021, 23, 375–390. [Google Scholar] [CrossRef]
- Shakouri, M.; Mandoulakani, B.A.; Norouzi, M. Effect of Soil Zinc Deficiency on Morphological Traits, Yield, and Yield Components and Activity of Zinc-Containing Enzymes in Zn-Efficient and -Inefficient Bread Wheat Cultivars. Commun. Soil. Sci. Plant Anal. 2022, 53, 2529–2542. [Google Scholar] [CrossRef]
- Li, X.; Zhang, L.; Ren, H.; Wang, X.; Mi, F. Zinc toxicity response in Ceratoides arborescens and identification of CaMTP, a novel zinc transporter. Front. Plant Sci. 2022, 13, 976311. [Google Scholar] [CrossRef]
- Chen, G.; Li, J.; Han, H.; Du, R.; Wang, X. Physiological and Molecular Mechanisms of Plant Responses to Copper Stress. Int. J. Mol. Sci. 2022, 23, 12950. [Google Scholar] [CrossRef] [PubMed]
- Nazir, F.; Hussain, A.; Fariduddin, Q. Hydrogen peroxide modulate photosynthesis and antioxidant systems in tomato (Solanum lycopersicum L.) plants under copper stress. Chemosphere 2019, 230, 544–558. [Google Scholar] [CrossRef] [PubMed]
- Leng, X.; Mu, Q.; Wang, X.; Li, X.; Zhu, X.; Shangguan, L.; Fang, J. Transporters, chaperones, and P-type ATPases controlling grapevine copper homeostasis. Funct. Integr. Genom. 2015, 15, 673–684. [Google Scholar] [CrossRef]
- Hunter, C.; Ware, M.A.; Gleason, S.M.; Pilon-Smits, E.; Pilon, M. Recovery after deficiency: Systemic copper prioritization and partitioning in the leaves and stems of hybrid poplar. Tree Physiol. 2022, 42, 1776–1785. [Google Scholar] [CrossRef]
- Schulten, A.; Pietzenuk, B.; Quintana, J.; Scholle, M.; Feil, R.; Krause, M.; Romera-Branchat, M.; Wahl, V.; Severing, E.; Coupland, G.; et al. Energy status-promoted growth and development of Arabidopsis require copper deficiency response transcriptional regulator SPL7. Plant Cell 2022, 34, 3873–3898. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, Y.; Ma, X.; Ouyang, Z.; Deng, L.; Shen, S.; Dong, X.; Du, N.; Dong, H.; Guo, Z.; et al. Melatonin Alleviates Copper Toxicity via Improving ROS Metabolism and Antioxidant Defense Response in Tomato Seedlings. Antioxidants 2022, 11, 758. [Google Scholar] [CrossRef]
- Raza Khan, A.; Fan, X.; Salam, A.; Azhar, W.; Ulhassan, Z.; Qi, J.; Liaquat, F.; Yang, S.; Gan, Y. Melatonin-mediated resistance to copper oxide nanoparticles-induced toxicity by regulating the photosynthetic apparatus, cellular damages and antioxidant defense system in maize seedlings. Environ. Pollut. 2022, 316 Pt 2, 120639. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.; Saleem, M.H.; Fahad, S.; Bashir, S.; Peng, D.X.; Deng, G.; Alamri, S.; Siddiqui, M.H.; Khan, S.M.; Shah, R.A.; et al. Effects of rice straw biochar and nitrogen fertilizer on ramie (Boehmeria nivea L.) morpho-physiological traits, copper uptake and post-harvest soil characteristics, grown in an aged-copper contaminated soil. J. Plant Nutr. 2021, 45, 11–24. [Google Scholar] [CrossRef]
- Yemets, A.; Horiunova, I.; Blume, Y. Cadmium, nickel, copper, and zinc influence on microfilament organization in Arabidopsis root cells. Cell Biol. Int. 2021, 45, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Abdulmajeed, A.M.; Alnusairi, G.S.H.; Alharbi, M.H.; Almushhin, A.; Hasan, M.M.; Soliman, M.H. Alleviation of copper phytotoxicity by acetylsalicylic acid and nitric oxide application in mung bean involves the up-regulation of antioxidants, osmolytes and glyoxalase system. J. Plant Interact. 2021, 16, 201–212. [Google Scholar] [CrossRef]
- Zaid, A.; Mohammad, F.; Wani, S.H.; Siddique, K.M.H. Salicylic acid enhances nickel stress tolerance by up-regulating antioxidant defense and glyoxalase systems in mustard plants. Ecotoxicol. Environ. Saf. 2019, 180, 575–587. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Hossain, M.A.; Fujita, M.; Tran, L.S. Physiological and biochemical mechanisms associated with trehalose-induced copper-stress tolerance in rice. Sci. Rep. 2015, 5, 11433. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Tripathi, A.K.; Nutan, K.K.; Sharma, S.; Ghosh, A.; Tripathi, J.K.; Pareek, A.; Singla-Pareek, S.L.; Sopory, S.K. A nuclear-localized rice glyoxalase I enzyme, OsGLYI-8, functions in the detoxification of methylglyoxal in the nucleus. Plant J. 2017, 89, 565–576. [Google Scholar] [CrossRef]
- Lin, F.; Xu, J.; Shi, J.; Li, H.; Li, B. Molecular cloning and characterization of a novel glyoxalase I gene TaGly I in wheat (Triticum aestivum L.). Mol. Biol. Rep. 2010, 37, 729–735. [Google Scholar] [CrossRef]
- Tan, Y.X.; Liang, J.; Zhang, Z.; Dong, S.Q.; Cao, J.F.; Qin, Q.P.; Zhang, Z.G.; Ni, D.A. Cloning and molecular characterisation of a putative glyoxalase I Gene (HfGlX I-1) of Daylily (Hemerocallis spp.). J. Plant Biochem. Biotechnol. 2022, 32, 265–273. [Google Scholar] [CrossRef]
- Li, Z.G. Methylglyoxal and Glyoxalase System in Plants: Old Players, New Concepts. Bot. Rev. 2016, 82, 183–203. [Google Scholar] [CrossRef]
- Kharbech, O.; Ben Massoud, M.; Sakouhi, L.; Djebali, W.; Mur, L.A.J.; Chaoui, A. Exogenous application of hydrogen sulfide reduces chromium toxicity in maize seedlings by suppressing NADPH oxidase activities and methylglyoxal accumulation. Plant Physiol. Biochem. 2020, 154, 646–656. [Google Scholar] [CrossRef]
- Mustafiz, A.; Ghosh, A.; Tripathi, A.K.; Kaur, C.; Ganguly, A.K.; Bhavesh, N.S.; Tripathi, J.K.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. A unique Ni2+-dependent and methylglyoxal-inducible rice glyoxalaseI possesses a single active site and functions in abiotic stress response. Plant J. 2014, 78, 951–963. [Google Scholar] [CrossRef]
- Singla-Pareek, S.L.; Yadav, S.K.; Pareek, A.; Reddy, M.K.; Sopory, S.K. Transgenic tobacco overexpressing glyoxalase pathway enzymes grow and set viable seeds in zinc-spiked soils. Plant Physiol. 2006, 140, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, S.M.; Hasanuzzaman, M.; Parvin, K.; Hossain, M.S.; Fujita, M. Protective role of tebuconazole and trifloxystrobin in wheat (Triticum aestivum L.) under cadmium stress via enhancement of antioxidant defense and glyoxalase systems. Physiol. Mol. Biol. Plants 2021, 27, 1043–1057. [Google Scholar] [CrossRef]
- Tang, F.; Li, R.; Zhou, Y.; Wang, S.; Zhou, Q.; Ding, Z.; Yao, Y.; Liu, J.; Wang, Y.; Hu, X.; et al. Genome-Wide Identification of Cassava Glyoxalase I Genes and the Potential Function of MeGLYⅠ-13 in Iron Toxicity Tolerance. Int. J. Mol. Sci. 2022, 23, 5212. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Corpas, F.J.; Ahmad, P. Salicylic acid-induced nitric oxide enhances arsenic toxicity tolerance in maize plants by upregulating the ascorbate-glutathione cycle and glyoxalase system. J. Hazard. Mater. 2020, 399, 123020. [Google Scholar] [CrossRef] [PubMed]
- Altaf, M.M.; Diao, X.-P.; Altaf, M.A.; Ur Rehman, A.; Shakoor, A.; Khan, L.U.; Jan, B.L.; Ahmad, P. Silicon-mediated metabolic upregulation of ascorbate glutathione (AsA-GSH) and glyoxalase reduces the toxic effects of vanadium in rice. J. Hazard. Mater. 2022, 436, 129145. [Google Scholar] [CrossRef]
- Shen, S.L.; Chen, J.Q.; Chang, J.J.; Xia, B.C. Using bioenergy crop cassava (Manihot esculenta) for reclamation of heavily metal-contaminated land. Int. J. Phytoremediat. 2020, 22, 1313–1320. [Google Scholar] [CrossRef]
- Gasparatos, D. Soil Contamination by Heavy Metals and Metalloids. Environments 2022, 9, 32. [Google Scholar] [CrossRef]
- Chen, Y.; Chao, Z.-F.; Jin, M.; Wang, Y.-L.; Li, Y.; Wu, J.-C.; Xiao, Y.; Peng, Y.; Lv, Q.-Y.; Gui, S.; et al. A heavy metal transporter gene ZmHMA3a promises safe agricultural production on cadmium-polluted arable land. J. Genet. Genom. 2022, 50, 130–134. [Google Scholar] [CrossRef]
- Liu, Y.; Nour-Eldin, H.H.; Zhang, L.; Li, Z.; Fernie, A.R.; Ren, M. Biotechnological detoxification: An unchanging source-sink balance strategy for crop improvement. Trends Plant Sci. 2022, 28, 135–138. [Google Scholar] [CrossRef]
- Balparda, M.; Schmitz, J.; Duemmel, M.; Wuthenow, I.C.; Schmidt, M.; Alseekh, S.; Fernie, A.R.; Lercher, M.J.; Maurino, V.G. Two plant glyoxalase systems with different evolutionary origins detoxify diverse reactive carbonyl species. Plant Physiol. 2022, 191, 1214–1233. [Google Scholar] [CrossRef]
- Manoj, V.M.; Anunanthini, P.; Swathik, P.C.; Dharshini, S.; Narayan, J.A.; Manickavasagam, M.; Sathishkumar, R.; Suresha, G.S.; Hemaprabha, G.; Ram, B.; et al. Comparative analysis of glyoxalase pathway genes in Erianthus arundinaceus and commercial sugarcane hybrid under salinity and drought conditions. BMC Genom. 2019, 19, 986. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Xiao, X.; Wang, N.; Zhang, F.; Gao, G.; Xu, K.; Chen, B.; Qiao, J.; Wu, X. Genome-wide analysis and expression profiles of glyoxalase gene families in Chinese cabbage (Brassica rapa L). PLoS ONE 2018, 13, e0191159. [Google Scholar] [CrossRef]
- Li, T.; Cheng, X.; Wang, Y.; Yin, X.; Li, Z.; Liu, R.; Liu, G.; Wang, Y.; Xu, Y. Genome-wide analysis of glyoxalase-like gene families in grape (Vitis vinifera L.) and their expression profiling in response to downy mildew infection. BMC Genom. 2019, 20, 362. [Google Scholar] [CrossRef] [PubMed]
- Jana, G.A.; Yaish, M.W. Functional characterization of the Glyoxalase-I (PdGLX1) gene family in date palm under abiotic stresses. Plant Signal Behav. 2020, 15, 1811527. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Ma, C.; Pan, Y.; Gong, S.; Zhao, C.; Chen, S.; Li, H. Sugar beet M14 glyoxalase I gene can enhance plant tolerance to abiotic stresses. J. Plant Res. 2013, 126, 415–425. [Google Scholar] [CrossRef]
- Zeng, Z.; Xiong, F.; Yu, X.; Gong, X.; Luo, J.; Jiang, Y.; Kuang, H.; Gao, B.; Niu, X.; Liu, Y. Overexpression of a glyoxalase gene, OsGly I, improves abiotic stress tolerance and grain yield in rice (Oryza sativa L.). Plant Physiol. Biochem. 2016, 109, 62–71. [Google Scholar] [CrossRef]
- Wu, Q.; Gao, S.; Pan, Y.B.; Su, Y.; Grisham, M.P.; Guo, J.; Xu, L.; Que, Y. Heterologous expression of a Glyoxalase I gene from sugarcane confers tolerance to several environmental stresses in bacteria. PeerJ 2018, 6, e5873. [Google Scholar] [CrossRef]
- Schmitz, J.; Dittmar, I.C.; Brockmann, J.D.; Schmidt, M.; Hüdig, M.; Rossoni, A.W.; Maurino, V.G. Defense against Reactive Carbonyl Species Involves at Least Three Subcellular Compartments Where Individual Components of the System Respond to Cellular Sugar Status. Plant Cell 2017, 29, 3234–3254. [Google Scholar] [CrossRef]
- Lentz, E.M.; Kuon, J.E.; Alder, A.; Mangel, N.; Zainuddin, I.M.; McCallum, E.J.; Anjanappa, R.B.; Gruissem, W.; Vanderschuren, H. Cassava geminivirus agroclones for virus-induced gene silencing in cassava leaves and roots. Plant Methods 2018, 14, 73. [Google Scholar] [CrossRef]
- Luo, X.Q.; An, F.F.; Xue, J.J.; Zhu, W.L.; Wei, Z.W.; Ou, W.J.; Li, K.M.; Chen, S.B.; Cai, J. Integrative analysis of metabolome and transcriptome reveals the mechanism of color formation in cassava (Manihot esculenta Crantz) leaves. Front. Plant Sci. 2023, 14, 1181257. [Google Scholar] [CrossRef]
- Tuo, D.C.; Zhou, P.; Yan, P.; Cui, H.G.; Liu, Y.; Wang, H.; Yang, X.K.; Liao, W.B.; Sun, D.; Li, X.Y.; et al. A cassava common mosaic virus vector for virus-induced gene silencing in cassava. Plant Methods 2021, 17, 74. [Google Scholar] [CrossRef]
- Zeng, H.Q.; Xie, Y.W.; Liu, G.Y.; Wei, Y.X.; Hu, W.; Shi, H.T. Agrobacterium-Mediated Gene Transient Overexpression and Tobacco Rattle Virus (TRV)-Based Gene Silencing in Cassava. Int. J. Mol. Sci. 2019, 20, 3976. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Lv, X.; Gao, G.; Li, F.; Li, J.; Qiao, J.; Xu, K.; Chen, B.; Wang, L.; Xiao, X.; et al. Identification and Characterization of a Glyoxalase I Gene in a Rapeseed Cultivar with Seed Thermotolerance. Front. Plant Sci. 2016, 7, 150. [Google Scholar] [CrossRef]
- Sas-Nowosielska, H.; Pawlas, N. Heavy metals in the cell nucleus—Role in pathogenesis. Acta Biochim. Pol. 2015, 62, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Gichner, T.; Patkova, Z.; Szakova, J.; Znidar, I.; Mukherjee, A. DNA damage in potato plants induced by cadmium, ethyl methanesulphonate and gamma-rays. Environ. Exp. Bot. 2008, 62, 113–119. [Google Scholar] [CrossRef]
- Doncheva, S.; Nikolov, B.; Ogneva, V. Effect of copper excess on the morphology of the nucleus in maize root meristem cells. Physiol. Plant. 1996, 96, 118–122. [Google Scholar] [CrossRef]
- Shen, J.; Su, Y.; Zhou, C.; Zhang, F.; Zhou, H.; Liu, X.; Wu, D.L.; Yin, X.C.; Xie, Y.J.; Yuan, X.X. A putative rice L-cysteine desulfhydrase encodes a true L-cysteine synthase that regulates plant cadmium tolerance. Plant Growth Regul. 2019, 89, 217–226. [Google Scholar] [CrossRef]
- Shi, Y.; Jiang, N.; Wang, M.T.; Du, Z.Y.; Chen, J.; Huang, Y.Y.; Li, M.Y.; Jin, Y.F.; Li, J.H.; Wan, J.; et al. OsHIPP17 is involved in regulating the tolerance of rice to copper stress. Front. Plant Sci. 2023, 14, 1183445. [Google Scholar] [CrossRef]
- Lin, X.; Li, R.; Zhou, Y.; Tang, F.; Wang, Y.; Lu, X.; Wang, S.; Yao, Y.; Liu, J.; Hu, X.; et al. Overexpression of Cassava MeAnn2 Enhances the Salt and IAA Tolerance of Transgenic Arabidopsis. Plants 2021, 10, 941. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, R.; Tang, F.; Che, Y.; Fernie, A.R.; Zhou, Q.; Ding, Z.; Yao, Y.; Liu, J.; Wang, Y.; Hu, X.; et al. MeGLYI-13, a Glyoxalase I Gene in Cassava, Enhances the Tolerance of Yeast and Arabidopsis to Zinc and Copper Stresses. Plants 2023, 12, 3375. https://doi.org/10.3390/plants12193375
Li R, Tang F, Che Y, Fernie AR, Zhou Q, Ding Z, Yao Y, Liu J, Wang Y, Hu X, et al. MeGLYI-13, a Glyoxalase I Gene in Cassava, Enhances the Tolerance of Yeast and Arabidopsis to Zinc and Copper Stresses. Plants. 2023; 12(19):3375. https://doi.org/10.3390/plants12193375
Chicago/Turabian StyleLi, Ruimei, Fenlian Tang, Yannian Che, Alisdair R. Fernie, Qin Zhou, Zhongping Ding, Yuan Yao, Jiao Liu, Yajie Wang, Xinwen Hu, and et al. 2023. "MeGLYI-13, a Glyoxalase I Gene in Cassava, Enhances the Tolerance of Yeast and Arabidopsis to Zinc and Copper Stresses" Plants 12, no. 19: 3375. https://doi.org/10.3390/plants12193375
APA StyleLi, R., Tang, F., Che, Y., Fernie, A. R., Zhou, Q., Ding, Z., Yao, Y., Liu, J., Wang, Y., Hu, X., & Guo, J. (2023). MeGLYI-13, a Glyoxalase I Gene in Cassava, Enhances the Tolerance of Yeast and Arabidopsis to Zinc and Copper Stresses. Plants, 12(19), 3375. https://doi.org/10.3390/plants12193375