
Selection and Yield Formation Characteristics of Dry Direct Seeding Rice in Northeast China

 ,
,
Abstract
:1. Introduction
2. Results
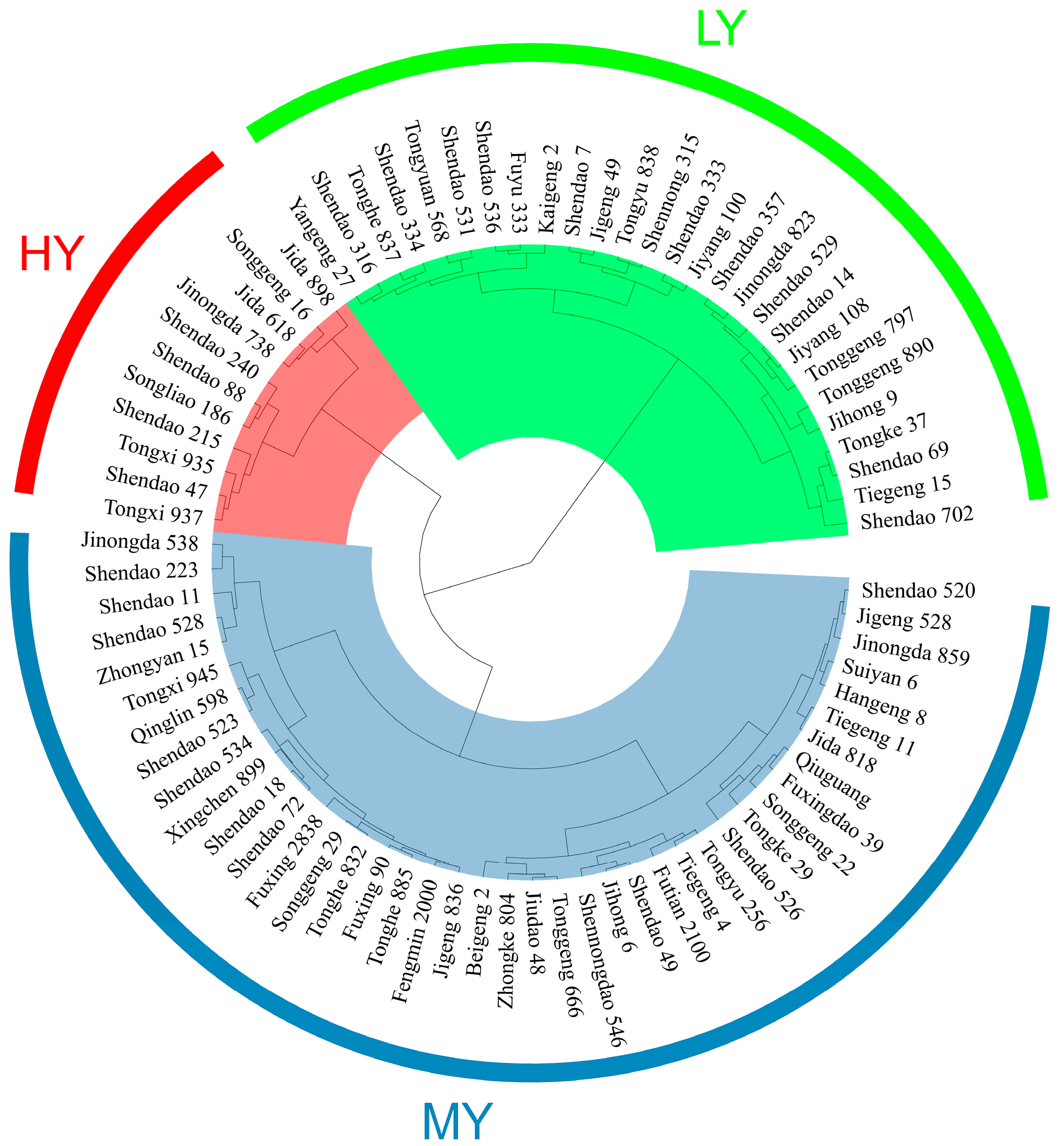
2.1. Grain Yield, Daily Yield and Yield Components
2.2. Growth Duration, Seedling Emergence Rate, Number of Stems, and Productive Stem Rate
2.3. Dry Matter Accumulation and Allocation
2.4. LAI, SPAD, HI, and Grain–Leaf Ratio
2.5. Correlation Analysis
2.6. Regression Analysis between Grain Yield and Growth Duration before Heading
3. Discussion
4. Materials and Methods
4.1. Experimental Sites and Soil Properties
4.2. Tested Materials
4.3. Experimental Design and Management
4.4. Measurements and Sampling
4.4.1. Growth Duration, Seedling Emergence Rate, Number of Stems, and Productive Stem Rate
4.4.2. Dry Matter Accumulation and Allocation
4.4.3. Leaf Area Index, SPAD, and Grain–Leaf Ratio
4.4.4. Grain Yield, Daily Yield, and Yield Components
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Khawar, B.S.C.; Gulshan, J. (Eds.) Rice Production Worldwide; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Peng, S.; Tang, Q.; Zou, Y. Current status and challenges of rice production in China. Plant Prod. Sci. 2009, 12, 3–8. [Google Scholar] [CrossRef]
- Sandhu, N.; Subedi, S.R.; Yadaw, R.B.; Chaudhary, B.; Prasai, H.; Iftekharuddaula, K.; Thanak, T.; Thun, V.; Battan, K.R.; Ram, M.; et al. Root traits enhancing rice grain yield under alternate wetting and drying condition. Front. Plant Sci. 2017, 8, 1879. [Google Scholar] [CrossRef] [PubMed]
- Bouman, B. How Much Water Does Rice Use? Agriscatt 1988 View Project Effects of Land Use on Regional Nitrous Oxide Emissions in the Humid Tropics of Costa Rica Extrapolating Fluxes from Field to Regional Scales View Project. Available online: https://www.researchgate.net/publication/281474989 (accessed on 11 January 2018).
- Tuong, T.P.; Bouman, B.A.M.; Mortimer, M. More rice, less water—Integrated approaches for increasing water productivity in irrigated rice-based systems in Asia. Plant Prod. Sci. 2005, 8, 231–241. [Google Scholar] [CrossRef]
- Mahajan, G.; Chauhan, B.S.; Gill, M.S. Dry-seeded rice culture in Punjab State of India: Lessons learned from farmers. Field Crops Res. 2013, 144, 89–99. [Google Scholar] [CrossRef]
- Deng, F.; He, L.; Chen, D.; Zhang, C.; Tian, Q.; Wu, Z.; Li, Q.; Zeng, Y.; Zhong, X.; Chen, H.; et al. Growth characteristics and grain yield of machine-transplanted medium indica hybrid rice with high daily yield. J. Integr. Agric. 2022, 21, 2547–2558. [Google Scholar] [CrossRef]
- Hossen, M.A.; Hossain, M.M.; Haque, M.E.; Bell, R.W. Transplanting into non-puddled soils with a small-scale mechanical transplanter reduced fuel, labour and irrigation water requirements for rice (Oryza sativa L.) establishment and increased yield. Field Crops Res. 2018, 225, 141–151. [Google Scholar] [CrossRef]
- Devkota, K.P.; Sudhir-Yadav; Khanda, C.M.; Beebout, S.J.; Mohapatra, B.K.; Singleton, G.R.; Puskur, R. Assessing alternative crop establishment methods with a sustainability lens in rice production systems of Eastern India. J. Clean. Prod. 2020, 244, 118835. [Google Scholar] [CrossRef]
- Wang, W.; Du, J.; Zhou, Y.; Zeng, Y.; Tan, X.; Pan, X.; Shi, Q.; Wu, Z.; Zeng, Y. Effects of different mechanical direct seeding methods on grain yield and lodging resistance of early indica rice in South China. J. Integr. Agric. 2021, 20, 1204–1215. [Google Scholar] [CrossRef]
- Jabran, K.; Ullah, E.; Akbar, N.; Yasin, M.; Zaman, U.; Nasim, W.; Riaz, M.; Arjumend, T.; Azhar, M.F.; Hussain, M. Growth and physiology of basmati rice under conventional and water-saving production systems. Arch. Agron. Soil Sci. 2017, 63, 1465–1476. [Google Scholar] [CrossRef]
- Wang, Z.; Gu, D.; Beebout, S.S.; Zhang, H.; Liu, L.; Yang, J.; Zhang, J. Effect of irrigation regime on grain yield, water productivity, and methane emissions in dry direct-seeded rice grown in raised beds with wheat straw incorporation. Crop J. 2018, 6, 495–508. [Google Scholar] [CrossRef]
- Liu, H.; Hussain, S.; Zheng, M.; Peng, S.; Huang, J.; Cui, K.; Nie, L. Dry direct-seeded rice as an alternative to transplanted-flooded rice in Central China. Agron. Sustain. Dev. 2015, 35, 285–294. [Google Scholar] [CrossRef]
- Sandhu, N.; Yadaw, R.B.; Chaudhary, B.; Prasai, H.; Iftekharuddaula, K.; Venkateshwarlu, C.; Annamalai, A.; Xangsayasane, P.; Battan, K.R.; Ram, M.; et al. Evaluating the performance of rice genotypes for improving yield and adaptability under direct seeded aerobic cultivation conditions. Front. Plant Sci. 2019, 10, 159. [Google Scholar] [CrossRef]
- Mahajan, G.; Gill, M.S.; Singh, K. Optimizing seed rate to suppress weeds and to increase yield in aerobic direct-seeded rice in Northwestern Indo-Gangetic plains. J. New Seeds 2010, 11, 225–238. [Google Scholar] [CrossRef]
- Sudhir-Yadav, E.; Humphreys, S.S.; Kukal, G.; Gill, R.R. Effect of water management on dry seeded and puddled transplanted rice. Part 2: Water balance and water productivity. Field Crops Res. 2011, 120, 123–132. [Google Scholar] [CrossRef]
- Wang, W.; Peng, S.; Liu, H.; Tao, Y.; Huang, J.; Cui, K.; Nie, L. The possibility of replacing puddled transplanted flooded rice with dry seeded rice in central China: A review. Field Crops Res. 2017, 214, 310–320. [Google Scholar] [CrossRef]
- Hossain, K.; Timsina, J.; Johnson, D.E.; Gathala, M.K.; Krupnik, T.J. Multi-year weed community dynamics and rice yields as influenced by tillage, crop establishment, and weed control: Implications for rice-maize rotations in the eastern Gangetic plains. Crop Prot. 2020, 138, 105334. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Siddique, K.H.M.; Rehman, H.; Aziz, T.; Lee, D.J.; Wahid, A. Rice direct seeding: Experiences, challenges and opportunities. Soil Tillage Res. 2011, 111, 87–98. [Google Scholar] [CrossRef]
- Yadav, S.; Singh, U.M.; Naik, S.M.; Venkateshwarlu, C.; Ramayya, P.J.; Raman, K.A.; Sandhu, N.; Kumar, A. Molecular mapping of QTLs associated with lodging resistance in dry direct-seeded rice (Oryza sativa L.). Front. Plant Sci. 2017, 8, 1431. [Google Scholar] [CrossRef]
- Sagare, D.B.; Abbai, R.; Jain, A.; Jayadevappa, P.K.; Dixit, S.; Singh, A.K.; Challa, V.; Alam, S.; Singh, U.M.; Yadav, S.; et al. More and more of less and less: Is genomics-based breeding of dry direct-seeded rice (DDSR) varieties the need of hour? Plant Biotechnol. J. 2020, 18, 2173–2186. [Google Scholar] [CrossRef]
- Liu, H.; Won, P.L.P.; Banayo, N.P.M.; Nie, L.; Peng, S.; Kato, Y. Late-season nitrogen applications improve grain yield and fertilizer-use efficiency of dry direct-seeded rice in the tropics. Field Crops Res. 2019, 233, 114–120. [Google Scholar] [CrossRef]
- Sandhu, N.; Torres, R.O.; Cruz, M.T.S.; Maturan, P.C.; Jain, R.; Kumar, A.; Henry, A. Traits and QTLs for development of dry direct-seeded rainfed rice varieties. J. Exp. Bot. 2015, 66, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, G.; Singh, K.; Singh, N.; Kaur, R.; Chauhan, B.S. Screening of water-efficient rice genotypes for dry direct seeding in South Asia. Arch. Agron. Soil Sci. 2018, 64, 103–115. [Google Scholar] [CrossRef]
- Sun, L.; Hussain, S.; Liu, H.; Peng, S.; Huang, J.; Cui, K.; Nie, L. Implications of low sowing rate for hybrid rice varieties under dry direct-seeded rice system in Central China. Field Crops Res. 2015, 175, 87–95. [Google Scholar] [CrossRef]
- Wang, W.; Chen, Q.; Hussain, S.; Mei, J.; Dong, H.; Peng, S.; Huang, J.; Cui, K.; Nie, L. Pre-sowing Seed Treatments in Direct-seeded Early Rice: Consequences for Emergence, Seedling Growth and Associated Metabolic Events under Chilling Stress. Sci. Rep. 2016, 6, 19637. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Jat, M.L.; Ganie, Z.A.; Chauhan, B.S.; Gupta, R.K. Herbicide options for effective weed management in dry direct-seeded rice under scented rice-wheat rotation of western Indo-Gangetic Plains. Crop. Prot. 2016, 81, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Hyun, D.Y.; Oh, M.W.; Choi, Y.M.; Lee, S.; Lee, M.C.; Oh, S. Morphological and molecular evaluation for germinability in rice varieties under low-temperature and anaerobic conditions. J. Crop Sci. Biotechnol. 2017, 20, 21–27. [Google Scholar] [CrossRef]
- Pal, R.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Impact of sowing date on yield, dry matter and nitrogen accumulation, and nitrogen translocation in dry-seeded rice in North-West India. Field Crops Res. 2017, 206, 138–148. [Google Scholar] [CrossRef]
- Mahajan, G.; Chauhan, B.S. Performance of dry direct-seeded rice in response to genotype and seeding rate. Agron. J. 2016, 108, 257–265. [Google Scholar] [CrossRef]
- Wei, H.; Zhang, H.; Blumwald, E.; Li, H.; Cheng, J.; Dai, Q.; Huo, Z.; Xu, K.; Guo, B. Different characteristics of high yield formation between inbred japonica super rice and inter-sub-specific hybrid super rice. Field Crops Res. 2016, 198, 179–187. [Google Scholar] [CrossRef]
- Xing, Z.P.; Wu, P.; Zhu, M.; Qian, H.J.; Hu, Y.J.; Guo, B.W.; Wei, H.Y.; Xu, K.; Huo, Z.Y.; Dai, Q.G.; et al. Temperature and solar radiation utilization of rice for yield formation with different mechanized planting methods in the lower reaches of the Yangtze River, China. J. Integr. Agric. 2017, 16, 1923–1935. [Google Scholar] [CrossRef]
- Zhang, C.; He, L.H.; Liao, S.; Gao, Y.T.; Zhu, S.L.; Li, B.; Zhou, W.; Chen, Y.; Hu, J.F.; Xiang, Z.F.; et al. Effect of sowing date on daily yield of mechanical indica hybrid rice under different ecological conditions. Acta Agron. Sin. 2020, 46, 1579–1590. [Google Scholar]
- Chauhan, B.S.; Prabhjyot, K.; Mahajan, G.; Randhawa, R.K.; Singh, H.; Kang, M.S. Global Warming and Its Possible Impact on Agriculture in India. In Advances in Agronomy; Academic Press Inc.: Cambridge, MA, USA, 2014; pp. 65–121. [Google Scholar] [CrossRef]
- Huang, L.; Yang, D.; Li, X.; Peng, S.; Wang, F. Coordination of high grain yield and high nitrogen use efficiency through large sink size and high post-heading source capacity in rice. Field Crops Res. 2019, 233, 49–58. [Google Scholar] [CrossRef]
- Mahender, A.; Anandan, A.; Pradhan, S.K. Early seedling vigour, an imperative trait for direct-seeded rice: An overview on physio-morphological parameters and molecular markers. Planta 2015, 241, 1027–1050. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Li, S.; Cheng, S.; Liu, Q.; Zhou, L.; Tao, Y.; Xing, Z.; Hu, Y.; Guo, B.; Wei, H.; et al. Increasing the appropriate seedling density for higher yield in dry direct-seeded rice sown by a multifunctional seeder after wheat-straw return. J. Integr. Agric. 2023, 22, 400–416. [Google Scholar] [CrossRef]
- Zhou, C.; Huang, Y.; Jia, B.; Wang, S.; Dou, F.; Samonte, S.O.P.B.; Chen, K.; Wang, Y. Optimization of nitrogen rate and planting density for improving the grain yield of different rice genotypes in Northeast China. Agronomy 2019, 9, 555. [Google Scholar] [CrossRef]
- Yang, W.; Peng, S.; Laza, R.C.; Visperas, R.M.; Dionisio-Sese, M.L. Grain yield and yield attributes of new plant type and hybrid rice. Crop Sci. 2007, 47, 1393–1400. [Google Scholar] [CrossRef]
- Peng, S.; Cassman, K.G.; Virmani, S.S.; Sheehy, J.; Khush, G.S. Yield Potential Trends of Tropical Rice since the Release of IR8 and the Challenge of Increasing Rice Yield Potential. Crop Sci. 1999, 39, 1552–1559. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, X.; Cao, J.; Li, Y.; Huang, J.; Peng, S. High nitrogen input reduces yield loss from low temperature during the seedling stage in early-season rice. Field Crops Res. 2018, 228, 68–75. [Google Scholar] [CrossRef]
- Deng, F.; Wang, L.; Mei, X.F.; Li, S.X.; Pu, S.L.; Ren, W.J. Polyaspartate urea and nitrogen management affect nonstructural carbohydrates and yield of rice. Crop Sci. 2016, 56, 3272–3285. [Google Scholar] [CrossRef]
- Yang, J.-C.; Yong, D.U.; Liu, H. Cultivation Approaches and Techniques for Annual Super-High-Yielding of Rice and Wheat in the Lower Reaches of Yangtze River. Sci. Agric. Sin. 2008, 41, 1611–1621. [Google Scholar] [CrossRef]
- Wu, G.; Zhang, H.; Huo, Z.; Xu, K.; Gao, H.; Wei, H.; Sha, A.; Xu, Z.; Qian, Z.; Sun, J. Characteristics of dry matter production and accumulation and Super-High Yield of japonica super rice in South China. Acta Agron. Sin. 2010, 36, 1921–1930. [Google Scholar]
- Wei, H.; Meng, T.; Li, X.; Dai, Q.; Zhang, H.; Yin, X. Sink-source relationship during rice grain filling is associated with grain nitrogen concentration. Field Crops Res. 2018, 215, 23–38. [Google Scholar] [CrossRef]
- Xu, L.; Zhan, X.; Yu, T.; Nie, L.; Huang, J.; Cui, K.; Wang, F.; Li, Y.; Peng, S. Yield performance of direct-seeded, double-season rice using varieties with short growth durations in central China. Field Crops Res. 2018, 227, 49–55. [Google Scholar] [CrossRef]
- Namikawa, M.; Matsunami, T.; Yabiku, T.; Takahashi, T.; Matsunami, M.; Hasegawa, T. Analysis of yield constraints and seasonal solar radiation and temperature limits for stable cultivation of dry direct-seeded rice in northeastern Japan. Field Crops Res. 2023, 295, 108896. [Google Scholar] [CrossRef]
- Ahmed, S.; Humphreys, E.; Chauhan, B.S. Optimum sowing date and cultivar duration of dry-seeded boro on the High Ganges River Floodplain of Bangladesh. Field Crops Res. 2016, 190, 91–102. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Grain Yield (t ha−1) | Daily Yield (kg ha−1 d−1) | Effective Panicles m−2 | Spikelets per Panicle | Spikelets m−2 (×103) | Grain Filling Rate (%) | 1000-Grain Weight (g) |
|---|---|---|---|---|---|---|---|
| HY | 10.05 a | 64.03 a | 458.03 a | 111.00 a | 49.98 a | 89.05 a | 24.88 a |
| MY | 7.74 b | 49.63 b | 414.63 b | 101.52 b | 41.18 b | 88.55 a | 24.06 b |
| LY | 5.75 c | 36.81 c | 307.16 c | 101.84 b | 30.85 c | 88.09 a | 25.16 a |
| Mean | 7.38 | 47.25 | 383.95 | 102.95 | 38.87 | 88.46 | 24.55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, C.; Li, Y.; Zhang, K.; Wu, Z.; Liu, J.; Liu, J.; Zhou, C.; Wang, S.; Li, F.; Sui, G. Selection and Yield Formation Characteristics of Dry Direct Seeding Rice in Northeast China. Plants 2023, 12, 3496. https://doi.org/10.3390/plants12193496
Liang C, Li Y, Zhang K, Wu Z, Liu J, Liu J, Zhou C, Wang S, Li F, Sui G. Selection and Yield Formation Characteristics of Dry Direct Seeding Rice in Northeast China. Plants. 2023; 12(19):3496. https://doi.org/10.3390/plants12193496
Chicago/Turabian StyleLiang, Chao, Yimeng Li, Kunhao Zhang, Zhouzhou Wu, Jiaxin Liu, Junfeng Liu, Chanchan Zhou, Shu Wang, Fenghai Li, and Guomin Sui. 2023. "Selection and Yield Formation Characteristics of Dry Direct Seeding Rice in Northeast China" Plants 12, no. 19: 3496. https://doi.org/10.3390/plants12193496
APA StyleLiang, C., Li, Y., Zhang, K., Wu, Z., Liu, J., Liu, J., Zhou, C., Wang, S., Li, F., & Sui, G. (2023). Selection and Yield Formation Characteristics of Dry Direct Seeding Rice in Northeast China. Plants, 12(19), 3496. https://doi.org/10.3390/plants12193496