Abstract
Over the centuries, human society has evolved based on the ability to select and use more adapted species for food supply, which means making plant species tastier and more productive in particular environmental conditions. However, nowadays, this scenario is highly threatened by climate change, especially by the changes in temperature and greenhouse gasses that directly affect photosynthesis, which highlights the need for strategic studies aiming at crop breeding and guaranteeing food security. This is especially worrying for crops with complex phenology, genomes with low variability, and the ones that support a large production chain, such as Coffea sp. L. In this context, recent advances shed some light on the genome function and transcriptional control, revealing small RNAs (sRNAs) that are responsible for environmental cues and could provide variability through gene expression regulation. Basically, sRNAs are responsive to environmental changes and act on the transcriptional and post-transcriptional gene silencing pathways that regulate gene expression and, consequently, biological processes. Here, we first discuss the predicted impact of climate changes on coffee plants and coffee chain production and then the role of sRNAs in response to environmental changes, especially temperature, in different species, together with their potential as tools for genetic improvement. Very few studies in coffee explored the relationship between sRNAs and environmental cues; thus, this review contributes to understanding coffee development in the face of climate change and towards new strategies of crop breeding.
1. Introduction
Due to climate changes and the importance of crops to human life, a lot of studies have been carried out toward understanding plant plasticity and the development of more tolerant varieties in biotic and abiotic stresses, together with higher productivity [1,2,3,4,5]. One of these important crops is Coffea sp L., which presents a complex phenological cycle and whose coffee fruit production is mainly affected by temperature changes, rainfall, and altitude [6,7,8,9]. Climate projections indicate that by the end of this century, global temperature may increase by 0.2 to 1.0 °C in the best scenario and from 2.4 to 4.8 °C in the most pessimistic scenario in comparison to the period from 1986 to 2005 [10]. In the long term, climate change, combined with such phenomena as longer and more unpredictable droughts or excessive rainfall, threatens the sustainability of coffee production on a global scale [11,12], which is also expected for other important crops [13,14].
The Coffea genus belongs to the Rubiaceae family with 130 described species [15]. Coffea arabica (Arabica coffee) and Coffea canephora are responsible for almost all the coffee bean production in the world [16]. Interestingly, the vast majority of coffee species are diploid and allogamous, while C. arabica is an allotetraploid and autogamous species that originated from the natural hybridization of non-reduced gametes of C. eugenioides and C. canephora [17,18]. Furthermore, C. arabica has a narrow genetic base, as described in the topic below, and due to favorable climatic conditions, together with genetic improvement programs, Brazil has become its largest producer in the world [19]. In this way, it is clear that a need arises for intelligent strategies to promote diversity and support genetic improvement, guaranteeing coffee production and, in a broader aspect, the adaptation of crops to climate change. In other species, one of these strategies includes the identification of new regulatory molecules such as small RNAs (sRNAs) [20,21,22]. These small RNAs act as post-transcriptional regulators of gene expression, degrading or blocking the target mRNA and regulating different processes, such as development, differentiation, flowering, metabolism, response to abiotic stress, and plant defense [23,24,25].
Elevated temperatures and heat stress negatively affect the distribution and yield of commercially important plants worldwide [26,27,28]. When the temperature rises 5 °C or more above the ideal, which is specific for each species, it is experienced as heat stress by all living organisms [29]. In the coffee tree, at the beginning of flowering, high temperatures associated with the intense water deficit cause the death of the pollen tubes by dehydration, causing the abortion of the flowers [30] or premature opening of the floral buds [31], which ends up generating losses in production. Furthermore, continuous exposure of C. arabica to temperatures close to 30 °C or more can result not only in repressed growth but also in anomalies such as yellowing of leaves [32], growth of tumors at the base of the stem [33], changes in the enzymes of the photosynthetic apparatus [34], and worsening of the grain quality [35].
Some studies have already been carried out in which microRNAs (miRNAs), a class of sRNAs responsive to heat stress by high temperatures, have been characterized, such as in Populus tomentosa [36], Arabidopsis thaliana [29], Oryza sativa [37], Saccharina japonica [38]; Solanum lycopersicum [39], Gossypium hirsutum [40], and Musa spp. AAA [41]. However, despite some miRNAs in common, the list of miRNAs with altered expression varies with the degree of stress and between species. Although sRNAs in coffee plants have already been addressed in some studies, such as the identification of miRNAs and their biogenesis pathways [42,43,44,45], miRNAs in floral development [46] and reproductive development [47], there is still a gap relating the role of sRNAs in temperature control in these species. Therefore, considering the peculiar aspect of C. arabica origin related to the narrow diversity and the need for more adapted genotypes in the face of climate changes, this review contributes to the understanding of the roles of sRNAs in coffee plants and discusses perspectives on its use as tools for crop breeding.
2. Coffee Plant: Origin, Phenology, and Environment-Related Aspects
In order to understand how the coffee tree behaves in the face of climate change, it is important to know how this plant emerged and spread throughout the world and how its very peculiar phenological aspects can contribute to this. The coffee tree belongs to the Rubiaceae family, genus Coffea, with 130 described species [15], the vast majority being diploid and allogamous; however, only C. arabica (Arabica coffee) and C. canephora (Robusta/Conilon coffee) are cultivated expressively. The other two species are cultivated on a smaller scale, C. liberica (Liberica coffee) and C. dewevrei (Excelsa coffee) [48]. In Brazil, 80% of the total production is C. arabica [16].
The origin and center of genetic diversity of C. arabica is in Southwestern Ethiopia [49,50], while C. canephora originated in Central and Western Africa [51,52,53]. C. arabica originated from the natural hybridization of non-reduced gametes of the diploid species, C. eugenioides and C. canephora [17,18,54], forming an allotetraploid species, 2n = 4x = 44, and it is considered autogamous with about 10% of allogamy [55]. This very rare hybridization event was estimated to have occurred from a single plant around 20,000 years ago [18] or, considering other predictions, between 100,000 and 665,000 years ago [56,57]. Particular events of that period seem to be related to thermal shocks that promoted the fusion of gametes and the shutdown of self-incompatibility systems of parent species, allowing for autogamy and reproduction [18], which can be related to the insertion of retrotransposons [56]. Transposons are endogenous DNA sequences capable of moving and multiplying in the host genome, impairing genes but also contributing to genome evolution [58,59,60], and in agreement with the stated before, these elements present higher activities under temperature changes [61,62]. Although the genetic base of C. arabica is narrow, the species has cultivars with great variability due to factors such as crossings, mutations, agronomic practices, and environment [63,64]. Currently, this species is widely distributed, being cultivated in regions with higher altitudes and milder temperatures, between 18 °C and 21 °C, in the American and Asian continents, as well as in some regions of Africa [64].
On the other hand, C. canephora is a diploid species, with 2n = 22 chromosomes, allogamous, originating from a wide hot, humid, and low-altitude region of the African continent, which extends from Guinea to the Democratic Republic of Congo [17,65]. C. canephora is widely cultivated in the African, American, and Asian continents, in places of low altitude and higher temperatures, with an annual average between 22 °C and 26 °C [66]. In regions with higher temperatures and abundant humidity, plants of the C. canephora species can reach up to 5 m in height. They are usually multi-stemmed, even in commercial crops with frequent thinning [64]. It is a highly polymorphic species with extensive geographical distribution, showing great adaptability to different environmental conditions [64]. This species was introduced in Brazil around 1920, in Espírito Santo, and its varieties have great variability in relation to the different agronomic and morphological characteristics due to the origin of the species and the fact that it reproduces by cross-self-fertilization [67].
Regarding its phenology, contrary to most angiosperms, which emit inflorescences and bear fruit in the same phenological year, coffee needs two years to complete its cycle [30]. The first year begins with vegetative growth characterized by the development of plagiotropic branches from orthotropic branches. The first phase, which runs from September to March, begins with the formation of axillary buds at the nodes of the primary plagiotropic branches [30,68]. From January to July, the differentiation of floral buds between early and late cultivars of C. arabica begins; it is known that the quality and intensity of light, as well as the photoperiod, affect plant growth, directly influencing floral development and many other characteristics; however, the inductive stimulus of the coffee reproductive cycle is still unclear and may be affected by the interaction of different environmental factors, in which molecular aspects are little explored [68]. Recently, López et al. [68] reviewed the endogenous and environmental factors related to the flowering process of C. arabica and proposed a model for the induction and floral development in Brazilian environmental conditions, where the floral induction starts in January, extending until July for early and late cultivars. However, based on gene expression data, López et al. [69] suggest a continuum of floral induction that allows different starting points for floral activation, which explains developmental asynchronicity and prolonged anthesis events in tropical perennial species.
3. Impacts of Increased Temperature and CO2 on Coffee Growing
Drought and unfavorable temperatures have always been described as the main climatic limitations for coffee production [70]. The main effects of climate change are displacement of optimal growing zones, changes in precipitation (amount and distribution), changes in crop disease/pest dynamics, and loss of agricultural land due to sea level rise and/or desertification [71]. As changes in the global climate are recognized, and coffee cultivation spreads to different regions where it was not cultivated before, concerns are increasing about these factors and the possible consequences for the production of this crop [32,70,71,72,73].
Indeed, temperature can limit the economic exploitation of coffee, in part because coffee growth is particularly affected by high and low temperatures [74]. The optimum average temperature range for C. arabica is reported to be between 18 and 21 °C; however, it should be noted that cultivars selected under intensive management conditions have allowed C. arabica plantations to be spread to regions with average annual temperatures of up to 24–25 °C, with satisfactory yields [35,73,75,76,77].
On the other hand, in regions with an average annual temperature below 18 °C [13,74,78], growth is largely repressed; in any case, it is clear that large variations in temperature cause defects in the beans, modify their biochemical composition, and alter the final quality of the beverage [13,74,78]. However, the pessimistic data obtained so far on how climate change affects the coffee crop had not considered the potential positive effects of high atmospheric CO2 concentration on coffee plant photosynthesis or the role of CO2 on heat tolerance and recognized resilience of elite coffee genotypes to acclimatize to stressful conditions [31].
In the physiological aspect, in general, plants feel and respond directly to the increase in atmospheric CO2 concentration through an increase in the rate of net photosynthesis (A) and reduction in stomatal conductance (gs); this is the basis for the effect of CO2 fertilization with a corresponding increase in yields [79,80]. Increases in A and CO2 concentration in the chloroplast of C3 plants are associated with stimulation of the rate of carboxylation of ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO) [80]. Furthermore, it has been shown systematically, but not universally, that gs decreases with high levels of CO2 concentration [81]. In general, decreases in gs often lead to lower transpiration rates and greater water use efficiency. However, this also results in latent heat loss and increasing leaf temperatures, which may impair crop performance in a global warming scenario [31].
Regarding productivity, the beneficial effect of increasing CO2 was demonstrated in a study carried out with C. arabica plants monitored for 3 years, where a higher yield of grain production was verified than in plants grown under ambient CO2 concentration [82]. However, the effects of the combination of high temperatures and CO2 concentrations on production yield and grain quality still need to be better explored.
In a study of C. arabica and C. canephora plants cultivated for 10 months at 25/20 °C (day/night) under 380 and 700 μL of CO2 L−1 and then subjected to an increase in temperature (0.5 °C per day) up to 42/34 °C (day/night), higher concentrations of CO2 strongly contributed to the impact of temperature on both species, promoting greater efficiency in water use, not causing negative photosynthetic regulation, in addition to increasing the protection molecules, as well as the activity of some antioxidant enzymes [76]. High concentrations of CO2, therefore, play a role in the sustainability of the coffee crop under future scenarios of climate change [75,76], as demonstrated by recent works on strengthening the capacity of moderate and severe drought tolerance [83,84,85,86].
Thus, although the studies are focused on a better understanding of how the physiological aspects respond to climatic variations and the importance of CO2 as a mitigator of the damage caused by the increase in temperature, crucial issues still need to be elucidated, such as, for example, how the concentration increased CO2 levels together with increased temperature can affect the plant–pest/disease balance, abnormal flower formation, and fruit development and quality. In addition, associating these results with molecular and genetic data may help to develop new ways to improve coffee, and in this sense, exploring sRNAs, in general, can be a good way to fill this gap since they are fine regulators of gene expression.
4. How Do Genetic and Molecular Components Contribute to the Coffee Plant’s Physiological Responses to Climate Change?
Genetic and molecular aspects are crucial to determining how organisms respond to different environmental conditions; however, studies relating these factors to physiological responses are limited. In the genetic and molecular aspect, the adaptive and evolutionary capacities of C. arabica and its parents (C. eugenioides and C. canephora) were evaluated under four thermal regimes (RTs; 18–14 °C, 23–19 °C, 28–24 °C, and 33–29 °C) [87]. The data indicate that C. arabica activates the genome inherited from each parent for different temperature conditions. For example, the growth rate was similar to that of C. canephora under the hottest RT and that of C. eugenioides under the coldest RT. For metabolite contents, C. arabica showed a response similar to that of C. canephora in the hottest RT [87].
At the level of gene expression, few differences between the allopolyploid and its parents were observed for studied genes linked to photosynthesis, respiration, and the circadian clock, while genes linked to redox activity showed a greater capacity of the allopolyploid for homeostasis [87]. In this way, the global transcriptional response to RTs of C. arabica was more homeostatic compared to its parents, and this provided greater phenotypic homeostasis when faced with unsuitable environments for the diploid parent species [87].
In another study, two genotypes of C. arabica (Acauã and Catuaí) were compared regarding leaf physiology, transcriptome, and carbohydrate/protein levels under optimal and elevated temperatures. It was demonstrated that the Acauã cultivar had lower leaf temperatures in both conditions than cv. Catuaí, while little or no difference was observed for the other leaf physiological parameters [88]. However, the genotypes showed a transcriptional restriction in the warmer temperatures. Differentially expressed genes responsive to temperature elevation revealed shared and genotype-specific genes, mainly related to carbohydrate metabolism [88]. These findings revealed intraspecific differences in the interconnected transcriptional and metabolic pathways that respond to warmer temperatures, potentially linked to coffee thermotolerance [88].
5. The sRNA Characteristics and Their Roles in Plant Stress Mediation
Most of the transcribed sequences of a genome do not encode proteins and are called non-coding RNAs (ncRNAs). Among the non-coding RNAs, we highlight three major groups of small RNAs (sRNAs) that act repressing gene expression through the transcriptional and post-transcriptional gene silencing pathways, TGS and PTGS, respectively, miRNA, small interfering RNAs (siRNAs), and phased secondary small interfering RNAs (phasiRNAs) [23,89,90]. Although sRNAs are biochemically very close, they differ from each other in their biogenesis to their mode of action. For example, microRNAs (miRNAs) are transcribed from MIR genes, while siRNAs are cleavage products of other RNAs [23,40,91], and phasiRNAs constitute an important category of small RNAs in plants, but most of their functions are still poorly defined [90].
MiRNAs are an extensive class of endogenous, small, non-coding RNAs (sncRNAs), ranging from 20 to 24 nucleotides in length, which are involved in mediating and silencing post-transcriptional gene expression. Although inhibition of translation by cleavage is a process classically mediated by siRNAs, miRNAs also play this role [23,92]. MiRNAs are encoded by MIR, which are transcribed by RNA polymerase II (POL II), while sRNAs are generally transcribed from the action of polymerases IV and V exclusive to plants [93]. The primary miRNA transcripts (pri-miRNA) are 5′-encapsulated and polyadenylated and are capable of forming imperfect folding structures called “hairpins”, which have a 5p and a 3p arm, with a size similar to the transcripts that encode proteins [92,94].
Although MIR genes often encode a single RNA transcript, there are few documented polycistronic miRNAs in plants, a phenomenon that is much more common in animals. Transcription of some miRNA loci can generate transcripts with different hairpin structures and sequences, but which can still be processed by the miRNA biogenesis machinery, resulting in different “mature” sequences [23,92,94]. In plants, the primary transcript (pri-miRNA) is stabilized by DAWDLE-binding proteins (DDL) and is further processed in the nucleus by the endonuclease activity of DICER-LIKE1 (DCL1) with the help of other proteins, such as RNA-binding proteins double-stranded (dsRNA) HYPONASTIC LEAVES 1 (HYL1) and SERRATE (SE) in precursors (pre-miRNAs) [95,96]. Then, the pre-miRNAs undergo subsequent cleavage by the DCL complex, forming a duplex structure [95].
Pre-miRNA in plants ranges from 49 to 900 nucleotides in length and undergoes processing by DCL1 or alternatively by DCL2, DCL3, and DCL4, forming a duplex with two laterally projecting 3′ nucleotides [95]. MiRNAs generally have 21 nucleotides, but the size varies depending on the DCL that performs the cleavage, with the distance between the RNase III and PAZ domains being suggested as a determinant of miRNA length [92,96]. The duplex is then methylated at the 3′ end by a methyltransferase, called HUA ENCHANCER1 (HEN1), to prevent further modification and degradation by exonucleases [97]. The HASTY protein (HST) transports the duplex to the cytoplasm [98], where one of the arms of the duplex is chosen and incorporated into a protein from the ARGONAUTA family (AGO) to form the RISC (RNA-Induced Silencing Complex). One of the domains of the AGO protein, called PIWI, has endonuclease activity and is capable of cleaving miRNA targets [98,99,100].
Elevated levels of CO2 and temperature can affect plant growth and development in general, but the signaling pathways that regulate these processes and which molecules are regulated by miRNAs under these conditions are still poorly understood. miRNAs can function in a complex regulatory network developed to deal with stressful conditions; several studies demonstrated that conserved miRNAs, such as miR160 and miR175, and new ones, had their expression altered by the elevation of temperature, which consequently resulted in the alteration of the expression of their genes target [36,39,101,102,103,104]; in addition, temperature rise can induce intron splicing of miRNAs [105].
In A. thaliana, the increase in temperature and CO2 were evaluated separately, and through small RNA sequencing, miRNAs whose expression is significantly altered in both situations were identified. Notably, almost all CO2-influenced miRNAs were inversely affected by elevated temperature. Furthermore, this study points out that the miR156/miR172 pathway, reported to act in the transition from vegetative to reproductive phase, probably has an important role in early flowering induced by high CO2 concentrations [105].
In sunflower (Helianthus annuus), a regulatory interaction between the transcription factor HaWRKY6 and miR396 was established. This miRNA, which normally acts in plant development through the regulation of GRF transcription factors, was activated for protection at high temperatures in sunflower (45 °C), a plant particularly well adapted to this type of stress. The identification of a young and divergent transcription factor under the negative control of a highly conserved regulatory element indicates that caution is needed when making inferences about the role of known miRNAs in non-model plants [106].
In the genus Coffea, several families of conserved and new microRNAs have been identified [42,43,45,107,108,109,110,111,112] in addition to the main proteins in the biosynthesis of sRNAs, such as DICER and Argonaute [44,113]. More recently, several types of non-coding RNAs were identified in C. canephora, in addition to miRNAs and their targets [114], and in C. arabica during floral development [46]. Hernández-Castellano et al. [115] identified some families of miRNAs in embryogenic callus; the data suggest a differential and cooperative role for miR535, miR164, miR2119, and miR157a during the initial stages of somatic embryogenesis in C. canephora, supporting their regulatory role in cellular totipotency of plants, which can bring benefits to genetic improvement programs. Regarding abiotic stress, a transcriptional profile of mRNAs and sRNAs in C. arabica roots has already been carried out in response to nitrogen lack [116]. In general, these data are important for the advancement of research with sRNAs in coffee plants; however, more studies are needed investigating the role of these molecules in adverse conditions such as high temperatures and high CO2 and their applicability as breeding tools.
SiRNAs, on the other hand, are generated from double-stranded RNAs (dsRNA) that can originate from different sources, such as RNAs transcribed from inverted repeats, natural cis-antisense transcription pairs, the action of RNA-dependent RNA polymerases (RDRs) that convert single-stranded RNA in dsRNA, the replication of RNA viruses, and regions of the genome rich in retroelements. DsRNA is cleaved into siRNAs of 21 to 24 nt by DCL proteins, and the size of siRNAs released depends on the specific catalytic activity of the respective DCL protein. DsRNA is usually cleaved by several DCL proteins, generating classes of siRNA with different sizes. Like miRNAs, siRNAs are loaded into RISC containing AGO protein that directs target regulation at either the post-transcriptional level or the transcriptional level through a pathway called RNA-directed DNA methylation (RdDM) [117].
DNA methylation is also an important epigenetic modification and plays a key role in regulating plant growth and development. In tobacco, the expression of methylated CycD3-1 and Nt-EXPA5 was found to be altered during heat stress [117]. In Populus simonii, the MIR393a, MIR156i, MIR167h, MIR396e, and MIR396g genes were methylated in CNG regions in heat-treated plants, while they were methylated in CG regions in cold-treated systems. These data demonstrate that methylation can regulate the expression of miRNAs under heat stress, further affecting the expression level of their targets, probably through the gene-silencing function of miRNAs [118].
The epigenetic marks are also being interpreted as part of the cell’s transcriptional memory in response to abiotic stress, so a deep comprehension of such mechanisms could assist in improving plant resilience. In agreement, a growing body of evidence suggests that the accumulation of signaling compounds and transcription factors (TFs) [118,119], together with epigenetic modification [120,121], play fundamental roles in the interplay between plant development and environmental signals. Very few examples are found in coffee; for example, Guedes et al. [122] studied the drought responses of tolerant and sensitive clones of C. canephora subjected to three drought cycles. The investigation of transcriptional memory to drought in the tolerant clone revealed genes related to ABA and a possible interaction between drought and biotic stress and memory genes. Furthermore, MYB proteins and miRNAs were found to modulate expression in the drought response. The drought-responsive genes identified in this work constitute valuable genomic resources to improve coffee cultivation and develop tolerant crops.
Finally, phasiRNAs constitute an important category of small RNAs in plants, but most of their functions are still poorly defined. The biogenesis of phasiRNAs occurs after cleavage of target mRNA or long non-coding RNA (lncRNA), typically (but not exclusively) by a 22-nucleotide miRNA [20]. After cleavage, the 5′ fragment of the target mRNA is rapidly degraded by a 3′→5′ exonucleolytic complex. The 3′ fragment is converted to double-stranded RNA (dsRNA) through the activity of (RDR6), which can be recruited by AGO1-RISC or AGO7-RISC and assisted by SUPPRESSOR OF GENE SILENCING3 (SGS3), which, in turn, can prevent degradation of the 3′ fragment of a 5′→3′ exoribonuclease. The resulting dsRNA is iteratively cleaved by a Dicer protein from the 5′ end of the “top” strand (derived from Pol II) containing the cleavage site, producing phasiRNA duplexes. The function of the Dicer family member DCL4 requires the assistance of a DOUBLE-STRANDED RNA BINDING FACTOR (DRB) protein to produce 21-nucleotide phasiRNAs. RISC containing phasiRNA subsequently interacts with target RNAs in a homology-dependent manner, such as with miRNAs [20].
Although phasiRNAs in plants are not yet fully elucidated as to their biological role, recent studies have shown that they play an important role in male fertility, such as in the development of anthers in maize (Zea mays) [123,124], wheat (Triticum aestivum), barley (Hordeum vulgare) [125], and rice (Oryza sativa) [126]; in addition, heat stress can negatively affect the reproduction of several species; recently, phasiRNAs have been found to be the most downregulated in response to heat stress in flax (Linum usitatissimum) reproductive tissues [126]. Recently, de Oliveira et al. [47] identified an increase in a specific group of phasiRNAs and the molecules involved in their biogenesis in the meiotic phase of coffee anthers. The meiotic phase of coffee anthers coincides with the G4 phase established by Moraes et al. [127], in which a latency stage of these buds is observed during the cold and dry periods in Brazil [68]. Furthermore, the pattern of accumulation of these molecules changes again after the rainy season and temperature increase, which demonstrates that these small molecules may be associated with the regulation of reproductive development in coffee plants in response to climate change.
As discussed above, sRNAs are molecules with great potential to be explored in the genetic improvement of coffee plants in the face of climate change. A good strategy for this is the use of RNA interference (RNAi) technology, which has already been extensively studied in plant disease control and plant protection strategies based on the knowledge that double-stranded RNA (dsRNA) molecules derived from plant or exogenous genes can silence essential or virulence genes in microbial pathogens and pests [128,129,130,131]. In this strategy, dsRNA induces the organism’s RISC silencing complex to silence target genes, as illustrated in Figure 1. These strategies are advantageous due to the highly specific, environmentally friendly, and malleable nature of dsRNAs [129].
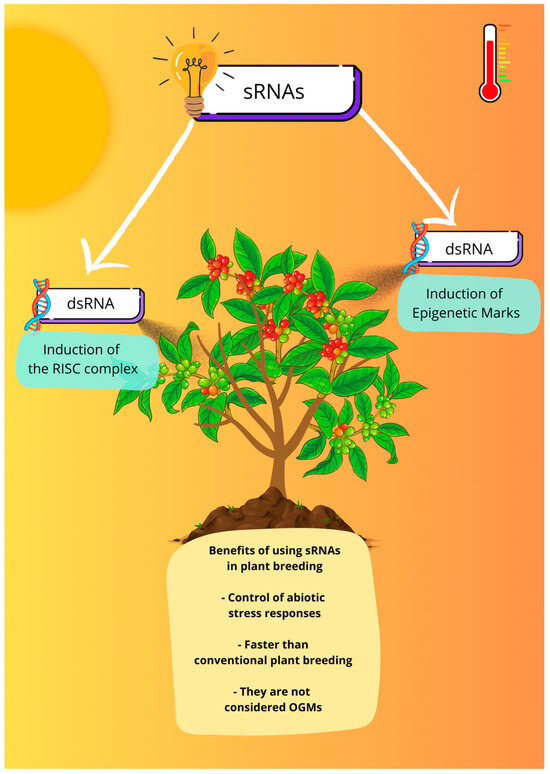
Figure 1.
Small RNAs as a tool for plant breeding. Understanding the role of sRNAs in response to abiotic stress can help in the development of new products based on RNAi technology, such as the exogenous application of dsRNA that can induce the silencing of target genes or be used to induce epigenetic marks to stimulate the transcriptional memory.
6. Conclusions
Considering the negative impacts of climate change on crop production and the particular characteristics of species, extensive studies are needed to deeply comprehend the regulatory mechanism of plant development to direct breeding programs. This is particularly important because our diet largely depends on fruits, and reproductive development is strongly affected by temperature changes. Despite the identification of several sRNAs responsive to temperature and CO2 elevation, studies that explore these molecules in response to these changes in coffee plants are scarce. Here, we explored this gap, starting with a discussion on how the origin and phenology of the coffee tree are related to its low genetic variability. Then, we discussed how the physiological and molecular mechanisms respond to increased temperature and CO2 levels and are related to sRNA pathways. The role of sRNAs in different species and how they could be useful for crop breeding was also discussed, although it has been poorly explored in coffee plants. In conclusion, sRNAs and their related mechanisms must be better explored in crops due to their potential, which will allow for the development of new products based on RNAi technology that can be used exogenously, applied at specific periods of the development, and are better accepted by society than transgenics, in addition to overcoming the time barrier that is required in assisted breeding, especially for perennial species with complex phenology, such as coffee plants.
Author Contributions
R.R.d.O. and A.C.-J. conceptualized the project; K.K.P.d.O. wrote the first version of this manuscript; K.K.P.d.O. and R.R.d.O. reviewed and wrote the final version of this manuscript; A.C.-J. supported the project. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the Instituto Nacional de Ciência e Tecnologia do Café (INCT/Café) under the grant of the Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG, CAP APQ 03605/17) and scholarship research for KKPO (5316/15), the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for scholarship research for ACJ under grant number 309005/2022-1, and Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, #21/06968-3).
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank the members of the Laboratory of Plant Molecular Physiology (LFMP, UFLA/Brazil) for discussions of the process and the funding agencies listed above.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tonietto, Â.; Sato, J.H.; Teixeira, J.B.; de Souza, E.M.; Pedrosa, F.O.; Franco, O.L.; Mehta, A. Proteomic Analysis of Developing Somatic Embryos of Coffea arabica. Plant Mol. Biol. Rep. 2012, 30, 1393–1399. [Google Scholar] [CrossRef]
- Bertrand, B.; Villegas Hincapié, A.M.; Marie, L.; Breitler, J.-C. Breeding for the Main Agricultural Farming of Arabica Coffee. Front. Sustain. Food Syst. 2021, 5, 709901. [Google Scholar] [CrossRef]
- Campuzano-Duque, L.F.; Blair, M.W. Strategies for Robusta Coffee (Coffea canephora) Improvement as a New Crop in Colombia. Agriculture 2022, 12, 1576. [Google Scholar] [CrossRef]
- Breitler, J.-C.; Etienne, H.; Léran, S.; Marie, L.; Bertrand, B. Description of an Arabica Coffee Ideotype for Agroforestry Cropping Systems: A Guideline for Breeding More Resilient New Varieties. Plants 2022, 11, 2133. [Google Scholar] [CrossRef]
- Zaidan, I.R.; Ferreira, A.; Noia, L.R.; Santos, J.G.; de Arruda, V.C.; do Couto, D.P.; Braz, R.A.; de Brites Senra, J.F.; Partelli, F.L.; Azevedo, C.F.; et al. Diversity and Structure of Coffea canephora from Old Seminal Crops in Espírito Santo, Brazil: Genetic Resources for Coffee Breeding. Tree Genet. Genomes 2023, 19, 19. [Google Scholar] [CrossRef]
- Mouen Bedimo, J.A.; Bieysse, D.; Nyassé, S.; Nottéghem, J.L.; Cilas, C. Role of Rainfall in the Development of Coffee Berry Disease in Coffea arabica Caused by Colletotrichum kahawae, in Cameroon. Plant Pathol. 2010, 59, 324–329. [Google Scholar] [CrossRef]
- Venancio, L.P.; Filgueiras, R.; Mantovani, E.C.; do Amaral, C.H.; da Cunha, F.F.; dos Santos Silva, F.C.; Althoff, D.; dos Santos, R.A.; Cavatte, P.C. Impact of Drought Associated with High Temperatures on Coffea canephora Plantations: A Case Study in Espírito Santo State, Brazil. Sci. Rep. 2020, 10, 19719. [Google Scholar] [CrossRef]
- Wagner, S.; Jassogne, L.; Price, E.; Jones, M.; Preziosi, R. Impact of Climate Change on the Production of Coffea arabica at Mt. Kilimanjaro, Tanzania. Agriculture 2021, 11, 53. [Google Scholar] [CrossRef]
- Dinh, T.L.A.; Aires, F.; Rahn, E. Statistical Analysis of the Weather Impact on Robusta Coffee Yield in Vietnam. Front. Environ. Sci. 2022, 10, 820916. [Google Scholar] [CrossRef]
- Lee, J.Y.; Marotzke, J.; Bala, G.; Cao, L.; Corti, S.; Dunne, J.P.; Engelbrecht, F.; Fischer, E.; Fyfe, J.C.; Jones, C.; et al. Chapter 4: Future Global Climate: Scenario-Based Projections and Near-Term Information. In Climate Change 2021: The Physical Science Basis; Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- DaMatta, F.M.; Grandis, A.; Arenque, B.C.; Buckeridge, M.S. Impacts of Climate Changes on Crop Physiology and Food Quality. Food Res. Int. 2010, 43, 1814–1823. [Google Scholar] [CrossRef]
- Bunn, C.; Läderach, P.; Ovalle Rivera, O.; Kirschke, D. A Bitter Cup: Climate Change Profile of Global Production of Arabica and Robusta Coffee. Clim. Chang. 2015, 129, 89–101. [Google Scholar] [CrossRef]
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate Trends and Global Crop Production Since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature Increase Reduces Global Yields of Major Crops in Four Independent Estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.P.; Rakotonasolo, F. Six New Species of Coffee (Coffea) from Northern Madagascar. Kew Bull. 2021, 76, 497–511. [Google Scholar] [CrossRef]
- Conab: Companhia Nacional De Abastecimento. Acomp. safra brasileira de café—Safra 2022-Quarto levantamento, n.4, v. 9, Brasília, 2022, p. 1–52. Available online: https://www.icafe.cr/wp-content/uploads/informacion_mercado/reportes_mercado/entidades_cafetaleras/Estimado%20de%20Cosecha%20de%20Brasil.pdf (accessed on 27 September 2023).
- Lashermes, P.; Combes, M.-C.; Robert, J.; Trouslot, P.; D’Hont, A.; Anthony, F.; Charrier, A. Molecular Characterisation and Origin of the Coffea arabica L. Genome. Mol. Genet. Genom. 1999, 261, 259–266. [Google Scholar] [CrossRef]
- Scalabrin, S.; Toniutti, L.; Di Gaspero, G.; Scaglione, D.; Magris, G.; Vidotto, M.; Pinosio, S.; Cattonaro, F.; Magni, F.; Jurman, I.; et al. A Single Polyploidization Event at the Origin of the Tetraploid Genome of Coffea arabica Is Responsible for the Extremely Low Genetic Variation in Wild and Cultivated Germplasm. Sci. Rep. 2020, 10, 4642. [Google Scholar] [CrossRef]
- ICO: International Cofee Organization. Estatísticas Do Comércio; ICO: International Cofee Organization: London, UK, 2022. [Google Scholar]
- Liu, X.; Chu, S.; Sun, C.; Xu, H.; Zhang, J.; Jiao, Y.; Zhang, D. Genome-Wide Identification of Low Phosphorus Responsive microRNAs in Two Soybean Genotypes by High-Throughput Sequencing. Funct. Integr. Genom. 2020, 20, 825–838. [Google Scholar] [CrossRef]
- Tahaei, S.A.; Nasri, M.; Soleymani, A.; Ghooshchi, F.; Oveysi, M. Plant Growth Regulators Affecting Corn (Zea mays L.) Physiology and Rab17 Expression under Drought Conditions. Biocatal. Agric. Biotechnol. 2022, 41, 102288. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, X. Endoplasmic Reticulum Stress-Responsive microRNAs Are Involved in the Regulation of Abiotic Stresses in Wheat. Plant Cell Rep. 2023, 42, 1433–1452. [Google Scholar] [CrossRef]
- Voinnet, O. Origin, Biogenesis, and Activity of Plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef]
- Tang, J.; Chu, C. MicroRNAs in Crop Improvement: Fine-Tuners for Complex Traits. Nat. Plants 2017, 3, 17077. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Sinha, P.; Singh, V.K.; Kumar, A.; Zhang, Q.; Bennetzen, J.L. 5Gs for Crop Genetic Improvement. Curr. Opin. Plant Biol. 2020, 56, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Habib-ur-Rahman, M.; Ahmad, A.; Raza, A.; Hasnain, M.U.; Alharby, H.F.; Alzahrani, Y.M.; Bamagoos, A.A.; Hakeem, K.R.; Ahmad, S.; Nasim, W.; et al. Impact of Climate Change on Agricultural Production; Issues, Challenges, and Opportunities in Asia. Front. Plant Sci. 2022, 13, 925548. [Google Scholar] [CrossRef] [PubMed]
- Lachaud, M.; Bravo-Ureta, B.E.; Ludena, C.E. Economic Effects of Climate Change on Agricultural Production and Productivity in Latin America and the Caribbean (LAC). Agric. Econ. 2022, 53, 321–332. [Google Scholar] [CrossRef]
- Mutengwa, C.S.; Mnkeni, P.; Kondwakwenda, A. Climate-Smart Agriculture and Food Security in Southern Africa: A Review of the Vulnerability of Smallholder Agriculture and Food Security to Climate Change. Sustainability 2023, 15, 2882. [Google Scholar] [CrossRef]
- Guan, Q.; Lu, X.; Zeng, H.; Zhang, Y.; Zhu, J. Heat Stress Induction of miR398 Triggers a Regulatory Loop That Is Critical for Thermotolerance in Arabidopsis. Plant J. 2013, 74, 840–851. [Google Scholar] [CrossRef]
- Camargo, Â.P.D.; Camargo, M.B.P.D. Definição e esquematização das fases fenológicas do cafeeiro arábica nas condições tropicais do Brasil. Bragantia 2001, 60, 65–68. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Rahn, E.; Läderach, P.; Ghini, R.; Ramalho, J.C. Why Could the Coffee Crop Endure Climate Change and Global Warming to a Greater Extent than Previously Estimated? Clim. Chang. 2019, 152, 167–178. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Ramalho, J.D.C. Impacts of Drought and Temperature Stress on Coffee Physiology and Production: A Review. Braz. J. Plant Physiol. 2006, 18, 55–81. [Google Scholar] [CrossRef]
- Franco, C.M. Influence of Temperature on Growth of Coffee Plant; IBEC Research Institute: New York, NY, USA, 1958. [Google Scholar]
- Dubberstein, D.; Lidon, F.C.; Rodrigues, A.P.; Semedo, J.N.; Marques, I.; Rodrigues, W.P.; Gouveia, D.; Armengaud, J.; Semedo, M.C.; Martins, S.; et al. Resilient and Sensitive Key Points of the Photosynthetic Machinery of Coffea spp. to the Single and Superimposed Exposure to Severe Drought and Heat Stresses. Front. Plant Sci. 2020, 11, 1049. [Google Scholar] [CrossRef]
- Ramalho, J.C.; Pais, I.P.; Leitão, A.E.; Guerra, M.; Reboredo, F.H.; Máguas, C.M.; Carvalho, M.L.; Scotti-Campos, P.; Ribeiro-Barros, A.I.; Lidon, F.J.C.; et al. Can Elevated Air [CO2] Conditions Mitigate the Predicted Warming Impact on the Quality of Coffee Bean? Front. Plant Sci. 2018, 9, 287. [Google Scholar] [CrossRef]
- Chen, L.; Ren, Y.; Zhang, Y.; Xu, J.; Sun, F.; Zhang, Z.; Wang, Y. Genome-Wide Identification and Expression Analysis of Heat-Responsive and Novel microRNAs in Populus Tomentosa. Gene 2012, 504, 160–165. [Google Scholar] [CrossRef]
- Sailaja, B.; Voleti, S.R.; Subrahmanyam, D.; Sarla, N.; Prasanth, V.V.; Bhadana, V.P.; Mangrauthia, S.K. Prediction and Expression Analysis of miRNAs Associated with Heat Stress in Oryza Sativa. Rice Sci. 2014, 21, 3–12. [Google Scholar] [CrossRef]
- Liu, F.; Wang, W.; Sun, X.; Liang, Z.; Wang, F. Conserved and Novel Heat Stress-Responsive microRNAs Were Identified by Deep Sequencing in Saccharina japonica (Laminariales, Phaeophyta). Plant Cell Environ. 2015, 38, 1357–1367. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Wang, Q.; Jiang, F.; Cao, X.; Sun, M.; Liu, M.; Wu, Z. Identification of miRNAs and Their Targets in Wild Tomato at Moderately and Acutely Elevated Temperatures by High-Throughput Sequencing and Degradome Analysis. Sci. Rep. 2016, 6, 33777. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, N.; Yang, X.; Tu, L.; Zhang, X. Small RNA-Mediated Responses to Low- and High-Temperature Stresses in Cotton. Sci. Rep. 2016, 6, 35558. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Zhang, Y.; Tang, R.; Qu, H.; Duan, X.; Jiang, Y. Banana sRNAome and Degradome Identify microRNAs Functioning in Differential Responses to Temperature Stress. BMC Genom. 2019, 20, 33. [Google Scholar] [CrossRef] [PubMed]
- Loss-Morais, G.; Ferreira, D.C.R.; Margis, R.; Alves-Ferreira, M.; Corrêa, R.L. Identification of Novel and Conserved microRNAs in Coffea canephora and Coffea arabica. Genet. Mol. Biol. 2014, 37, 671–682. [Google Scholar] [CrossRef]
- Chaves, S.S.; Fernandes-Brum, C.N.; Silva, G.F.F.; Ferrara-Barbosa, B.C.; Paiva, L.V.; Nogueira, F.T.S.; Cardoso, T.C.S.; Amaral, L.R.; de Souza Gomes, M.; Chalfun-Junior, A. New Insights on Coffea miRNAs: Features and Evolutionary Conservation. Appl. Biochem. Biotechnol. 2015, 177, 879–908. [Google Scholar] [CrossRef]
- Fernandes-Brum, C.N.; Rezende, P.M.; Ribeiro, T.H.C.; de Oliveira, R.R.; Cunha de Sousa Cardoso, T.; do Amaral, L.R.; de Souza Gomes, M.; Chalfun-Junior, A. A Genome-Wide Analysis of the RNA-Guided Silencing Pathway in Coffee Reveals Insights into Its Regulatory Mechanisms. PLoS ONE 2017, 12, e0176333. [Google Scholar] [CrossRef]
- BIBI, F.; BAROZAI, M.; DIN, M. Bioinformatics Profiling and Characterization of potentialmicroRNAs and Their Targets in the Genus Coffea. Turk. J. Agric. For. 2017, 41, 191–200. [Google Scholar] [CrossRef]
- Ribeiro, T.H.C.; Baldrich, P.; de Oliveira, R.R.; Fernandes-Brum, C.N.; Mathioni, S.M.; Cunha de Sousa Cardoso, T.; de Souza Gomes, M.; do Amaral, L.R.; de Oliveira, K.K.P.; dos Reis, G.L.; et al. The Floral Development of the Allotetraploid Coffea arabica L. Correlates with a Small RNA Dynamic Reprogramming. bioRxiv 2023. [Google Scholar] [CrossRef]
- de Oliveira, K.K.P.; de Oliveira, R.R.; de Campos Rume, G.; Ribeiro, T.H.C.; Fernandes-Brum, C.N.; Kakrana, A.; Mathioni, S.; Meyers, B.C.; de Souza Gomes, M.; Chalfun-Junior, A. Coffee Microsporogenesis and Related Small Interfering RNAs Biosynthesis Have a Unique Pattern among Eudicots Suggesting a Sensitivity to Climate Changes. bioRxiv 2023. [Google Scholar] [CrossRef]
- Davis, A.P.; Govaerts, R.; Bridson, D.M.; Stoffelen, P. An Annotated Taxonomic Conspectus of the Genus Coffea (Rubiaceae). Bot. J. Linn. Soc. 2006, 152, 465–512. [Google Scholar] [CrossRef]
- Aga, E.; Bekele, E.; Bryngelsson, T. Inter-Simple Sequence Repeat (ISSR) Variation in Forest Coffee Trees (Coffea arabica L.) Populations from Ethiopia. Genetica 2005, 124, 213–221. [Google Scholar] [CrossRef]
- Silvestrini, M.; Junqueira, M.G.; Favarin, A.C.; Guerreiro-Filho, O.; Maluf, M.P.; Silvarolla, M.B.; Colombo, C.A. Genetic Diversity and Structure of Ethiopian, Yemen and Brazilian Coffea arabica L. Accessions Using Microsatellites Markers. Genet. Resour. Crop Evol. 2007, 54, 1367–1379. [Google Scholar] [CrossRef]
- Ferwerda, F.P. Coffee. In Evolution of Crop Plants; Longman: London, UK, 1976; pp. 257–260. [Google Scholar]
- Gomez, C.; Dussert, S.; Hamon, P.; Hamon, S.; de Kochko, A.; Poncet, V. Current Genetic Differentiation of Coffea canephoraPierre ex A. Froehn in the Guineo-Congolian African Zone: Cumulative Impact of Ancient Climatic Changes and Recent Human Activities. BMC Evol. Biol. 2009, 9, 167. [Google Scholar] [CrossRef]
- Musoli, P.; Cubry, P.; Aluka, P.; Billot, C.; Dufour, M.; De Bellis, F.; Pot, D.; Bieysse, D.; Charrier, A.; Leroy, T. Genetic Differentiation of Wild and Cultivated Populations: Diversity of Coffea canephora Pierre in Uganda. Genome 2009, 52, 634–646. [Google Scholar] [CrossRef]
- Sakiyama, N.S.; Pereira, A.A.; Zambolim, L. Melhoramento Do Café Arábica. Melhoramento de Espécies Cultivadas; Imprensa Universitária: Viçosa, Brazil, 1999. [Google Scholar]
- Carvalho, A.; Mônaco, L.C.; Campana, M.P. Melhoramento do cafeeiro: XXVII—Ensaio de seleções regionais de Jaú. Bragantia 1964, 23, 129–142. [Google Scholar] [CrossRef]
- Yu, Q.; Guyot, R.; de Kochko, A.; Byers, A.; Navajas-Pérez, R.; Langston, B.J.; Dubreuil-Tranchant, C.; Paterson, A.H.; Poncet, V.; Nagai, C.; et al. Micro-Collinearity and Genome Evolution in the Vicinity of an Ethylene Receptor Gene of Cultivated Diploid and Allotetraploid Coffee Species (Coffea). Plant J. 2011, 67, 305–317. [Google Scholar] [CrossRef]
- Cenci, A.; Combes, M.-C.; Lashermes, P. Genome Evolution in Diploid and Tetraploid Coffea Species as Revealed by Comparative Analysis of Orthologous Genome Segments. Plant Mol. Biol. 2012, 78, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Feschotte, C.; Pritham, E.J. DNA Transposons and the Evolution of Eukaryotic Genomes. Annu. Rev. Genet. 2007, 41, 331–368. [Google Scholar] [CrossRef] [PubMed]
- Mirouze, M.; Vitte, C. Transposable Elements, a Treasure Trove to Decipher Epigenetic Variation: Insights from Arabidopsis and Crop Epigenomes. J. Exp. Bot. 2014, 65, 2801–2812. [Google Scholar] [CrossRef] [PubMed]
- Catlin, N.S.; Josephs, E.B. The Important Contribution of Transposable Elements to Phenotypic Variation and Evolution. Curr. Opin. Plant Biol. 2022, 65, 102140. [Google Scholar] [CrossRef]
- Lv, Y.; Hu, F.; Zhou, Y.; Wu, F.; Gaut, B.S. Maize Transposable Elements Contribute to Long Non-Coding RNAs That Are Regulatory Hubs for Abiotic Stress Response. BMC Genom. 2019, 20, 864. [Google Scholar] [CrossRef]
- Pargana, A.; Musacchia, F.; Sanges, R.; Russo, M.T.; Ferrante, M.I.; Bowler, C.; Zingone, A. Intraspecific Diversity in the Cold Stress Response of Transposable Elements in the Diatom Leptocylindrus Aporus. Genes 2019, 11, 9. [Google Scholar] [CrossRef]
- Anthony, F.; Bertrand, B.; Quiros, O.; Wilches, A.; Lashermes, P.; Berthaud, J.; Charrier, A. Genetic Diversity of Wild Coffee (Coffea arabica L.) Using Molecular Markers. Euphytica 2001, 118, 53–65. [Google Scholar] [CrossRef]
- Guerreiro-Filho, O. Origem e Classificação Botânica Do Cafeeiro. In Cultivares de Café: Origem, Características e Recomendações; Embrapa Café: Brasília, Brazil, 2008; pp. 23–30. [Google Scholar]
- Conagin, C.H.T.M.; Mendes, A.J.T. Pesquisas citológicas e genéticas em três espécies de Coffea: Auto-incompatibilidade em Coffea canephora pierre ex froehner. Bragantia 1961, 20, 788–804. [Google Scholar] [CrossRef]
- Mendes, A.N.G.; Guimarães, R.J.; Souza, C.A.S. Classificação Botânica, Origem e Distribuição Geográfica Do Cafeeiro. In Cafeicultura; UFLA/FAEPE: Lavras, Brazil, 2002; pp. 285–300. [Google Scholar]
- da Eira Aguiar, A.T.; Guerreiro-Filho, O.; Maluf, M.P.; Gallo, P.B.; Fazuoli, L.C. Caracterização de cultivares de Coffea arabica mediante utilização de descritores mínimos. Bragantia 2004, 63, 179–192. [Google Scholar] [CrossRef]
- López, M.E.; Santos, I.S.; de Oliveira, R.R.; Lima, A.A.; Cardon, C.H.; Chalfun-Junior, A.; López, M.E.; Santos, I.S.; de Oliveira, R.R.; Lima, A.A.; et al. An Overview of the Endogenous and Environmental Factors Related to the Coffea arabica Flowering Process. Beverage Plant Res. 2021, 1, 13. [Google Scholar] [CrossRef]
- Cardon, C.H.; de Oliveira, R.R.; Lesy, V.; Ribeiro, T.H.C.; Fust, C.; Pereira, L.P.; Colasanti, J.; Chalfun-Junior, A. Expression of Coffee Florigen CaFT1 Reveals a Sustained Floral Induction Window Associated with Asynchronous Flowering in Tropical Perennials. Plant Sci. 2022, 325, 111479. [Google Scholar] [CrossRef] [PubMed]
- Cheserek, J.J.; Gichimu, B.M. Drought and Heat Tolerance in Coffee: A Review; University of Embu: Embu, Kenya, 2012. [Google Scholar]
- Kimemia, J.K. Effect of Global Warming on Coffee Production. In Proceedings of the Ugandan Coffee Traders Federation Breakfast Fellowship, Kampala, Uganda, 15 June 2010. [Google Scholar]
- Ovalle-Rivera, O.; Läderach, P.; Bunn, C.; Obersteiner, M.; Schroth, G. Projected Shifts in Coffea arabica Suitability among Major Global Producing Regions Due to Climate Change. PLoS ONE 2015, 10, e0124155. [Google Scholar] [CrossRef] [PubMed]
- Läderach, P.; Ramirez–Villegas, J.; Navarro-Racines, C.; Zelaya, C.; Martinez–Valle, A.; Jarvis, A. Climate Change Adaptation of Coffee Production in Space and Time. Clim. Chang. 2017, 141, 47–62. [Google Scholar] [CrossRef]
- Silva, E.A.; DaMatta, F.M.; Ducatti, C.; Regazzi, A.J.; Barros, R.S. Seasonal Changes in Vegetative Growth and Photosynthesis of Arabica Coffee Trees. Field Crops Res. 2004, 89, 349–357. [Google Scholar] [CrossRef]
- Martins, M.Q.; Rodrigues, W.P.; Fortunato, A.S.; Leitão, A.E.; Rodrigues, A.P.; Pais, I.P.; Martins, L.D.; Silva, M.J.; Reboredo, F.H.; Partelli, F.L.; et al. Protective Response Mechanisms to Heat Stress in Interaction with High [CO2] Conditions in Coffea spp. Front Plant Sci. 2016, 7, 947. [Google Scholar] [CrossRef]
- Rodrigues, W.P.; Martins, M.Q.; Fortunato, A.S.; Rodrigues, A.P.; Semedo, J.N.; Simões-Costa, M.C.; Pais, I.P.; Leitão, A.E.; Colwell, F.; Goulao, L.; et al. Long-Term Elevated Air [CO2] Strengthens Photosynthetic Functioning and Mitigates the Impact of Supra-Optimal Temperatures in Tropical Coffea arabica and C. canephora Species. Glob. Chang. Biol. 2016, 22, 415–431. [Google Scholar] [CrossRef] [PubMed]
- Scotti-Campos, P.; Pais, I.P.; Ribeiro-Barros, A.I.; Martins, L.D.; Tomaz, M.A.; Rodrigues, W.P.; Campostrini, E.; Semedo, J.N.; Fortunato, A.S.; Martins, M.Q.; et al. Lipid Profile Adjustments May Contribute to Warming Acclimation and to Heat Impact Mitigation by Elevated [CO2] in Coffea spp. Environ. Exp. Bot. 2019, 167, 103856. [Google Scholar] [CrossRef]
- Carr, M. The Water Relations and Irrigation Requirements of Coffee. Exp. Agric. 2001, 37, 1–36. [Google Scholar] [CrossRef]
- Long, S.P.; Ainsworth, E.A.; Leakey, A.D.B.; Nösberger, J.; Ort, D.R. Food for Thought: Lower-Than-Expected Crop Yield Stimulation with Rising CO2 Concentrations. Science 2006, 312, 1918–1921. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The Response of Photosynthesis and Stomatal Conductance to Rising [CO2]: Mechanisms and Environmental Interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Engineer, C.B.; Hashimoto-Sugimoto, M.; Negi, J.; Israelsson-Nordström, M.; Azoulay-Shemer, T.; Rappel, W.-J.; Iba, K.; Schroeder, J.I. CO2 Sensing and CO2 Regulation of Stomatal Conductance: Advances and Open Questions. Trends Plant Sci. 2016, 21, 16–30. [Google Scholar] [CrossRef]
- Ghini, R.; Neto, A.; Dentzien, A.; Gerreiro-Filho, O.; Jost, R.; Patrício, F.; Marinho-Prado, J.; Thomaziello, R.A.; Bettiol, W.; DaMatta, F. Coffee Growth, Pest and Yield Responses to Free-Air CO2 Enrichment. Clim. Chang. 2015, 132, 307–320. [Google Scholar] [CrossRef]
- Avila, R.; De Almeida, W.; Cavalcante, L.; Machado, K.; Barbosa, M.; Sousa, R.; Martino, P.; Toral, M.; Marçal, D.; Martins, S.; et al. Elevated Air [CO2] Improves Photosynthetic Performance and Alters Biomass Accumulation and Partitioning in Drought-Stressed Coffee Plants. Environ. Exp. Bot. 2020, 177, 104137. [Google Scholar] [CrossRef]
- Semedo, J.N.; Rodrigues, A.P.; Lidon, F.C.; Pais, I.P.; Marques, I.; Gouveia, D.; Armengaud, J.; Silva, M.J.; Martins, S.; Semedo, M.C.; et al. Intrinsic Non-Stomatal Resilience to Drought of the Photosynthetic Apparatus in Coffea spp. Is Strengthened by Elevated Air [CO2]. Tree Physiol. 2021, 41, 708–727. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, I.; Marques, I.; Paulo, O.S.; Batista, D.; Partelli, F.L.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C.; Ribeiro-Barros, A.I. Understanding the Impact of Drought in Coffea Genotypes: Transcriptomic Analysis Supports a Common High Resilience to Moderate Water Deficit but a Genotype Dependent Sensitivity to Severe Water Deficit. Agronomy 2021, 11, 2255. [Google Scholar] [CrossRef]
- Marques, I.; Rodrigues, A.P.; Gouveia, D.; Lidon, F.C.; Martins, S.; Semedo, M.C.; Gaillard, J.-C.; Pais, I.P.; Semedo, J.N.; Scotti-Campos, P.; et al. High-Resolution Shotgun Proteomics Reveals That Increased Air [CO2] Amplifies the Acclimation Response of Coffea Species to Drought Regarding Antioxidative, Energy, Sugar, and Lipid Dynamics. J. Plant Physiol. 2022, 276, 153788. [Google Scholar] [CrossRef]
- Bertrand, B.; Bardil, A.; Baraille, H.; Dussert, S.; Doulbeau, S.; Dubois, E.; Severac, D.; Dereeper, A.; Etienne, H. The Greater Phenotypic Homeostasis of the Allopolyploid Coffea arabica Improved the Transcriptional Homeostasis over That of Both Diploid Parents. Plant Cell Physiol. 2015, 56, 2035–2051. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, R.R.; Ribeiro, T.H.C.; Cardon, C.H.; Fedenia, L.; Maia, V.A.; Barbosa, B.C.F.; Caldeira, C.F.; Klein, P.E.; Chalfun-Junior, A. Elevated Temperatures Impose Transcriptional Constraints and Elicit Intraspecific Differences between Coffee Genotypes. Front. Plant Sci. 2020, 11, 1113. [Google Scholar] [CrossRef]
- Brodersen, P.; Sakvarelidze-Achard, L.; Bruun-Rasmussen, M.; Dunoyer, P.; Yamamoto, Y.Y.; Sieburth, L.; Voinnet, O. Widespread Translational Inhibition by Plant miRNAs and siRNAs. Science 2008, 320, 1185–1190. [Google Scholar] [CrossRef]
- Zheng, J.; Chen, C.; Li, G.; Chen, P.; Liu, Y.; Xia, R. Biogenesis of Reproductive PhasiRNAs: Exceptions to the Rules. Plant Biotechnol. J. 2023, 21, 241–243. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, H.; Fang, S.; Kang, Y.; Wu, W.; Hao, Y.; Li, Z.; Bu, D.; Sun, N.; Zhang, M.Q.; et al. NONCODE 2016: An Informative and Valuable Data Source of Long Non-Coding RNAs. Nucleic Acids Res. 2016, 44, D203–D208. [Google Scholar] [CrossRef] [PubMed]
- Budak, H.; Akpinar, B.A. Plant miRNAs: Biogenesis, Organization and Origins. Funct. Integr. Genom. 2015, 15, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Law, J.A.; Jacobsen, S.E. Establishing, Maintaining and Modifying DNA Methylation Patterns in Plants and Animals. Nat. Rev. Genet. 2010, 11, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Axtell, M.J.; Westholm, J.O.; Lai, E.C. Vive La Différence: Biogenesis and Evolution of microRNAs in Plants and Animals. Genome Biol. 2011, 12, 221. [Google Scholar] [CrossRef]
- Margis, R.; Fusaro, A.F.; Smith, N.A.; Curtin, S.J.; Watson, J.M.; Finnegan, E.J.; Waterhouse, P.M. The Evolution and Diversification of Dicers in Plants. FEBS Lett. 2006, 580, 2442–2450. [Google Scholar] [CrossRef]
- Bologna, N.G.; Voinnet, O. The Diversity, Biogenesis, and Activities of Endogenous Silencing Small RNAs in Arabidopsis. Annu. Rev. Plant Biol. 2014, 65, 473–503. [Google Scholar] [CrossRef]
- Ramachandran, V.; Chen, X. Degradation of microRNAs by a Family of Exoribonucleases in Arabidopsis. Science 2008, 321, 1490–1492. [Google Scholar] [CrossRef]
- Park, M.Y.; Wu, G.; Gonzalez-Sulser, A.; Vaucheret, H.; Poethig, R.S. Nuclear Processing and Export of microRNAs in Arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 3691–3696. [Google Scholar] [CrossRef]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef]
- Debat, H.J.; Ducasse, D.A. Plant microRNAs: Recent Advances and Future Challenges. Plant Mol. Biol. Rep. 2014, 32, 1257–1269. [Google Scholar] [CrossRef]
- Yu, X.; Wang, H.; Lu, Y.; de Ruiter, M.; Cariaso, M.; Prins, M.; van Tunen, A.; He, Y. Identification of Conserved and Novel microRNAs That Are Responsive to Heat Stress in Brassica Rapa. J. Exp. Bot. 2012, 63, 1025–1038. [Google Scholar] [CrossRef] [PubMed]
- Kruszka, K.; Pacak, A.; Swida-Barteczka, A.; Nuc, P.; Alaba, S.; Wroblewska, Z.; Karlowski, W.; Jarmolowski, A.; Szweykowska-Kulinska, Z. Transcriptionally and Post-Transcriptionally Regulated microRNAs in Heat Stress Response in Barley. J. Exp. Bot. 2014, 65, 6123–6135. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Yang, L.; Fan, D.; Li, L.; Hao, Q. Integration Analysis of miRNA-mRNA Pairs between Two Contrasting Genotypes Reveals the Molecular Mechanism of Jujube (Ziziphus jujuba Mill.) Response to High-Temperature Stress; Research Square: Durham, NC, USA, 2023. [Google Scholar]
- Amini, Z.; Salehi, H.; Chehrazi, M.; Etemadi, M.; Xiang, M. miRNAs and Their Target Genes Play a Critical Role in Response to Heat Stress in Cynodon dactylon (L.) Pers. Mol. Biotechnol. 2023, 1–14. [Google Scholar] [CrossRef] [PubMed]
- May, P.; Liao, W.; Wu, Y.; Shuai, B.; McCombie, W.R.; Zhang, M.Q.; Liu, Q.A. The Effects of Carbon Dioxide and Temperature on microRNA Expression in Arabidopsis Development. Nat. Commun. 2013, 4, 2145. [Google Scholar] [CrossRef]
- Giacomelli, J.I.; Weigel, D.; Chan, R.L.; Manavella, P.A. Role of Recently Evolved miRNA Regulation of Sunflower HaWRKY6 in Response to Temperature Damage. New Phytol. 2012, 195, 766–773. [Google Scholar] [CrossRef]
- Nellikunnumal, S.M.; Chandrashekar, A. Computational Identification of Conserved MicroRNA and Their Targets in Coffea canephora by EST Analysis. Dyn. Biochem. Process Biotechnol. Mol. Biol. 2012, 6, 70–76. [Google Scholar]
- Mohanan, S.; Gowda, K.; Kandukuri, S.V.; Chandrashekar, A. Involvement of a Novel Intronic microRNA in Cross Regulation of N-Methyltransferase Genes Involved in Caffeine Biosynthesis in Coffea canephora. Gene 2013, 519, 107–112. [Google Scholar] [CrossRef]
- Akter, A.; Islam, M.M.; Mondal, S.I.; Mahmud, Z.; Jewel, N.A.; Ferdous, S.; Amin, M.R.; Rahman, M.M. Computational Identification of miRNA and Targets from Expressed Sequence Tags of Coffee (Coffea arabica). Saudi J. Biol. Sci. 2014, 21, 3–12. [Google Scholar] [CrossRef]
- Rebijith, K.B.; Asokan, R.; Ranjitha, H.H.; Krishna, V.; Nirmalbabu, K. In Silico Mining of Novel microRNAs from Coffee (Coffea arabica) Using Expressed Sequence Tags. J. Hortic. Sci. Biotechnol. 2013, 88, 325–337. [Google Scholar] [CrossRef]
- Devi, K.J.; Chakraborty, S.; Deb, B.; Rajwanshi, R. Computational Identification and Functional Annotation of microRNAs and Their Targets from Expressed Sequence Tags (ESTs) and Genome Survey Sequences (GSSs) of Coffee (Coffea arabica L.). Plant Gene 2016, 6, 30–42. [Google Scholar] [CrossRef]
- Ivamoto, S.T.; Júnior, O.R.; Domingues, D.S.; dos Santos, T.B.; de Oliveira, F.F.; Pot, D.; Leroy, T.; Vieira, L.G.E.; Carazzolle, M.F.; Pereira, G.A.G.; et al. Transcriptome Analysis of Leaves, Flowers and Fruits Perisperm of Coffea arabica L. Reveals the Differential Expression of Genes Involved in Raffinose Biosynthesis. PLoS ONE 2017, 12, e0169595. [Google Scholar] [CrossRef]
- Parvez Mosharaf, M.; Akond, Z.; Hadiul Kabir, M.; Nurul Haque Mollah, M. Genome-Wide Identification, Characterization and Phylogenetic Analysis of Dicer-like (DCL) Gene Family in Coffea arabica. Bioinformation 2019, 15, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Lemos, S.M.C.; Fonçatti, L.F.C.; Guyot, R.; Paschoal, A.R.; Domingues, D.S. Genome-Wide Screening and Characterization of Non-Coding RNAs in Coffea canephora. Non-Coding RNA 2020, 6, 39. [Google Scholar] [CrossRef]
- Hernández-Castellano, S.; Andrade-Marcial, M.; Aguilar-Méndez, E.D.; Loyola-Vargas, V.M.; de Folter, S.; De-la-Peña, C. MiRNA Expression Analysis during Somatic Embryogenesis in Coffea canephora. Plant Cell Tissue Organ Cult. 2022, 150, 177–190. [Google Scholar] [CrossRef]
- dos Santos, T.B.; Soares, J.D.M.; Lima, J.E.; Silva, J.C.; Ivamoto, S.T.; Baba, V.Y.; Souza, S.G.H.; Lorenzetti, A.P.R.; Paschoal, A.R.; Meda, A.R.; et al. An Integrated Analysis of mRNA and sRNA Transcriptional Profiles in Coffea arabica L. Roots: Insights on Nitrogen Starvation Responses. Funct. Integr. Genom. 2019, 19, 151–169. [Google Scholar] [CrossRef]
- Centomani, I.; Sgobba, A.; D’Addabbo, P.; Dipierro, N.; Paradiso, A.; De Gara, L.; Dipierro, S.; Viggiano, L.; de Pinto, M.C. Involvement of DNA Methylation in the Control of Cell Growth during Heat Stress in Tobacco BY-2 Cells. Protoplasma 2015, 252, 1451–1459. [Google Scholar] [CrossRef]
- Conrath, U. Molecular Aspects of Defence Priming. Trends Plant Sci. 2011, 16, 524–531. [Google Scholar] [CrossRef]
- Santos, A.P.; Serra, T.; Figueiredo, D.D.; Barros, P.; Lourenço, T.; Chander, S.; Oliveira, M.M.; Saibo, N.J.M. Transcription Regulation of Abiotic Stress Responses in Rice: A Combined Action of Transcription Factors and Epigenetic Mechanisms. OMICS J. Integr. Biol. 2011, 15, 839–857. [Google Scholar] [CrossRef]
- Vriet, C.; Hennig, L.; Laloi, C. Stress-Induced Chromatin Changes in Plants: Of Memories, Metabolites and Crop Improvement. Cell. Mol. Life Sci. 2015, 72, 1261–1273. [Google Scholar] [CrossRef]
- Crisp, P.A.; Ganguly, D.; Eichten, S.R.; Borevitz, J.O.; Pogson, B.J. Reconsidering Plant Memory: Intersections between Stress Recovery, RNA Turnover, and Epigenetics. Sci. Adv. 2016, 2, e1501340. [Google Scholar] [CrossRef]
- de Freitas Guedes, F.A.; Nobres, P.; Rodrigues Ferreira, D.C.; Menezes-Silva, P.E.; Ribeiro-Alves, M.; Correa, R.L.; DaMatta, F.M.; Alves-Ferreira, M. Transcriptional Memory Contributes to Drought Tolerance in Coffee (Coffea canephora) Plants. Environ. Exp. Bot. 2018, 147, 220–233. [Google Scholar] [CrossRef]
- Yadava, P.; Tamim, S.; Zhang, H.; Teng, C.; Zhou, X.; Meyers, B.C.; Walbot, V. Transgenerational Conditioned Male Fertility of HD-ZIP IV Transcription Factor Mutant Ocl4: Impact on 21-Nt phasiRNA Accumulation in Pre-Meiotic Maize Anthers. Plant Reprod. 2021, 34, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Zhai, J.; Zhang, H.; Arikit, S.; Huang, K.; Nan, G.-L.; Walbot, V.; Meyers, B.C. Spatiotemporally Dynamic, Cell-Type–Dependent Premeiotic and Meiotic phasiRNAs in Maize Anthers. Proc. Natl. Acad. Sci. USA 2015, 112, 3146–3151. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, S.; Pokhrel, S.; Czymmek, K.; Meyers, B.C. Premeiotic, 24-Nucleotide Reproductive PhasiRNAs Are Abundant in Anthers of Wheat and Barley But Not Rice and Maize. Plant Physiol. 2020, 184, 1407–1423. [Google Scholar] [CrossRef] [PubMed]
- Pokhrel, S.; Meyers, B.C. Heat-responsive microRNAs and Phased Small Interfering RNAs in Reproductive Development of Flax. Plant Direct 2022, 6, e385. [Google Scholar] [CrossRef]
- Morais, H.; Caramori, P.H.; Koguishi, M.S.; de Arruda Ribeiro, A.M. Escala fenológica detalhada da fase reprodutiva de Cooffea arabica. Bragantia 2008, 67, 257–260. [Google Scholar] [CrossRef]
- Wytinck, N.; Manchur, C.L.; Li, V.H.; Whyard, S.; Belmonte, M.F. dsRNA Uptake in Plant Pests and Pathogens: Insights into RNAi-Based Insect and Fungal Control Technology. Plants 2020, 9, 1780. [Google Scholar] [CrossRef]
- Šečić, E.; Kogel, K.-H. Requirements for Fungal Uptake of dsRNA and Gene Silencing in RNAi-Based Crop Protection Strategies. Curr. Opin. Biotechnol. 2021, 70, 136–142. [Google Scholar] [CrossRef]
- Lucena-Leandro, V.S.; Abreu, E.F.A.; Vidal, L.A.; Torres, C.R.; Junqueira, C.I.C.V.F.; Dantas, J.; Albuquerque, É.V.S. Current Scenario of Exogenously Induced RNAi for Lepidopteran Agricultural Pest Control: From dsRNA Design to Topical Application. Int. J. Mol. Sci. 2022, 23, 15836. [Google Scholar] [CrossRef]
- Qiao, L.; Niño-Sánchez, J.; Hamby, R.; Capriotti, L.; Chen, A.; Mezzetti, B.; Jin, H. Artificial Nanovesicles for dsRNA Delivery in Spray-Induced Gene Silencing for Crop Protection. Plant Biotechnol. J. 2023, 21, 854–865. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).