
Microsatellite Markers: A Tool to Assess the Genetic Diversity of Yellow Mustard (Sinapis alba L.)
 , , , , and
, , , , and
Abstract
:1. Introduction
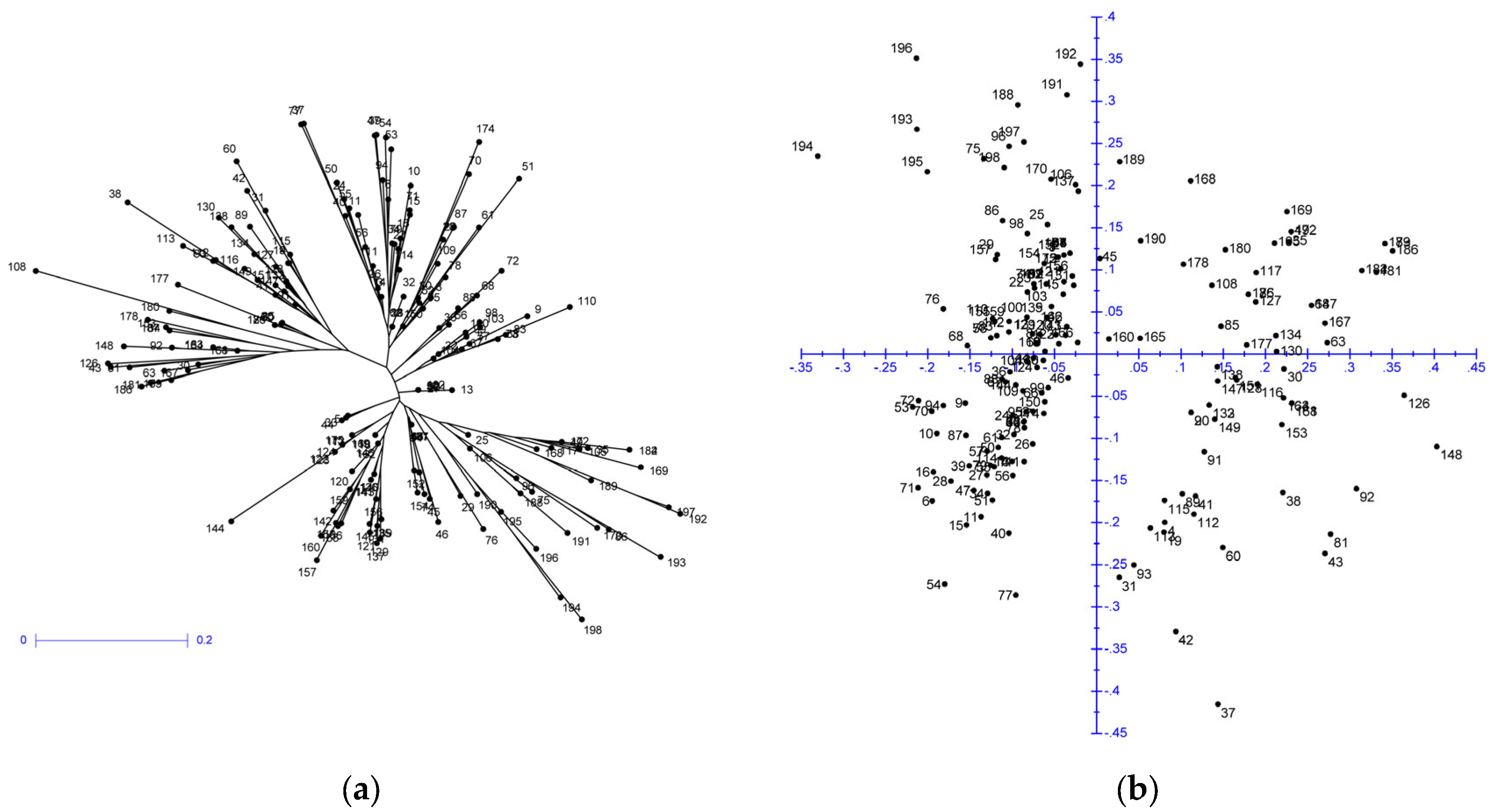
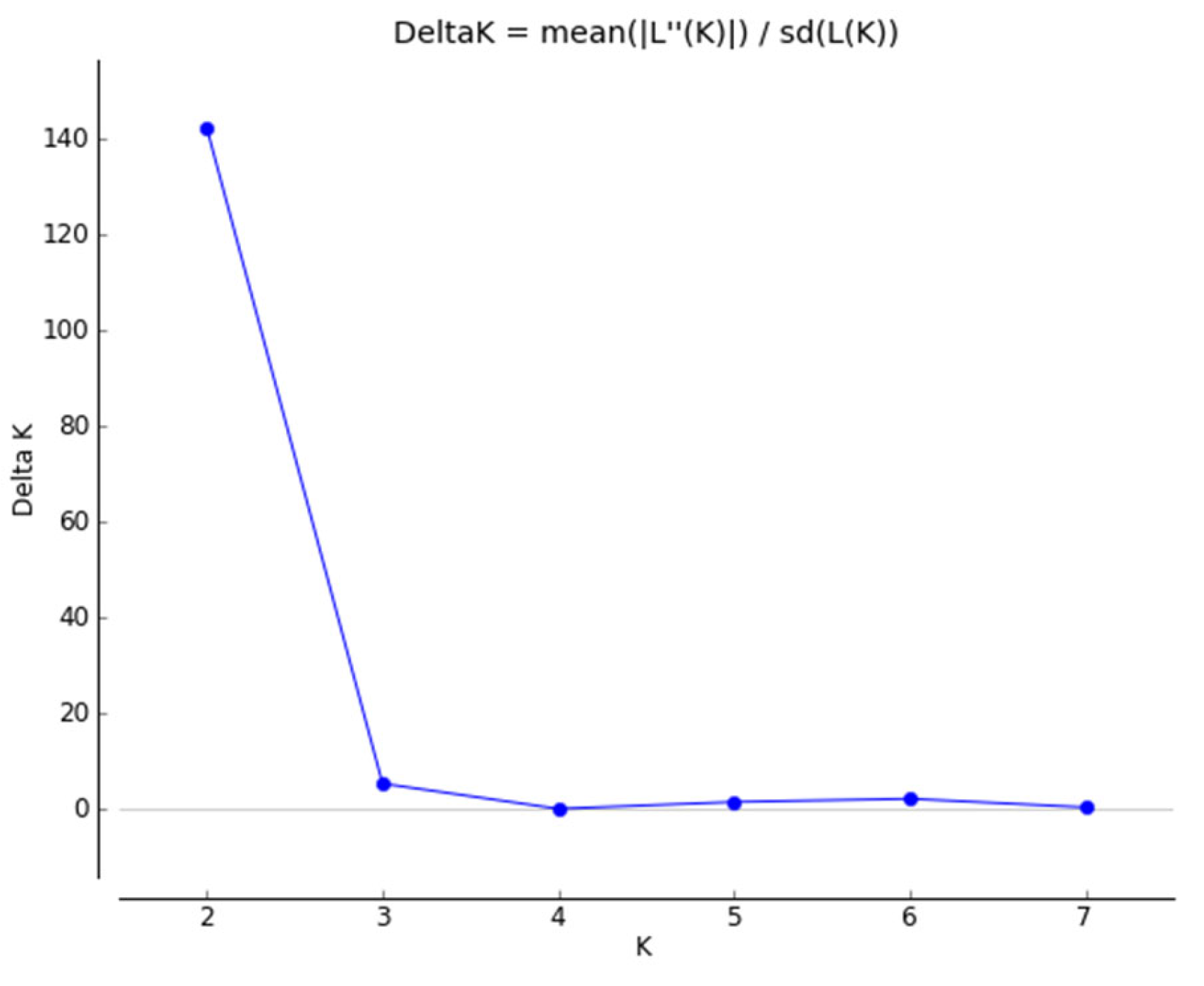
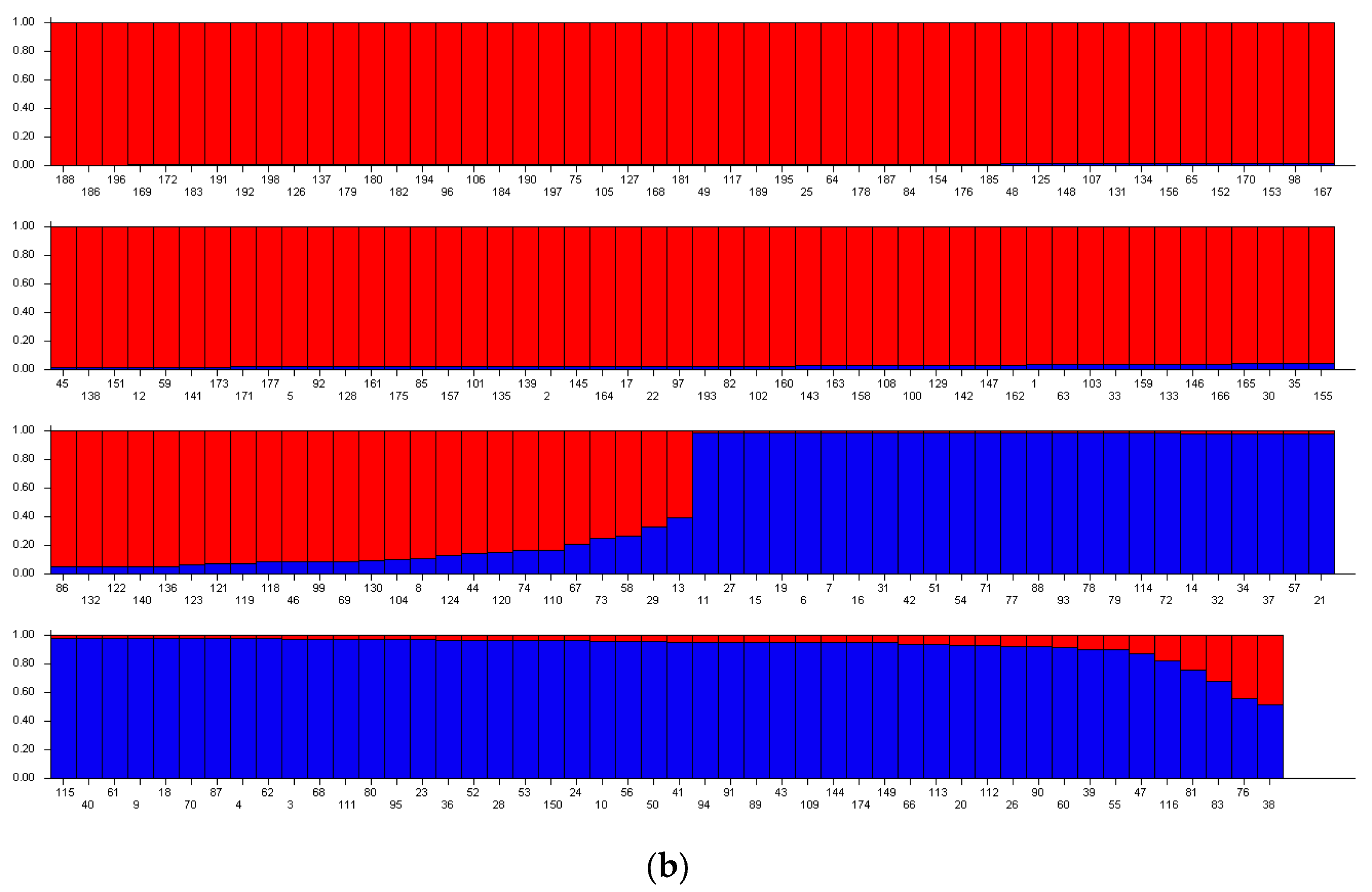
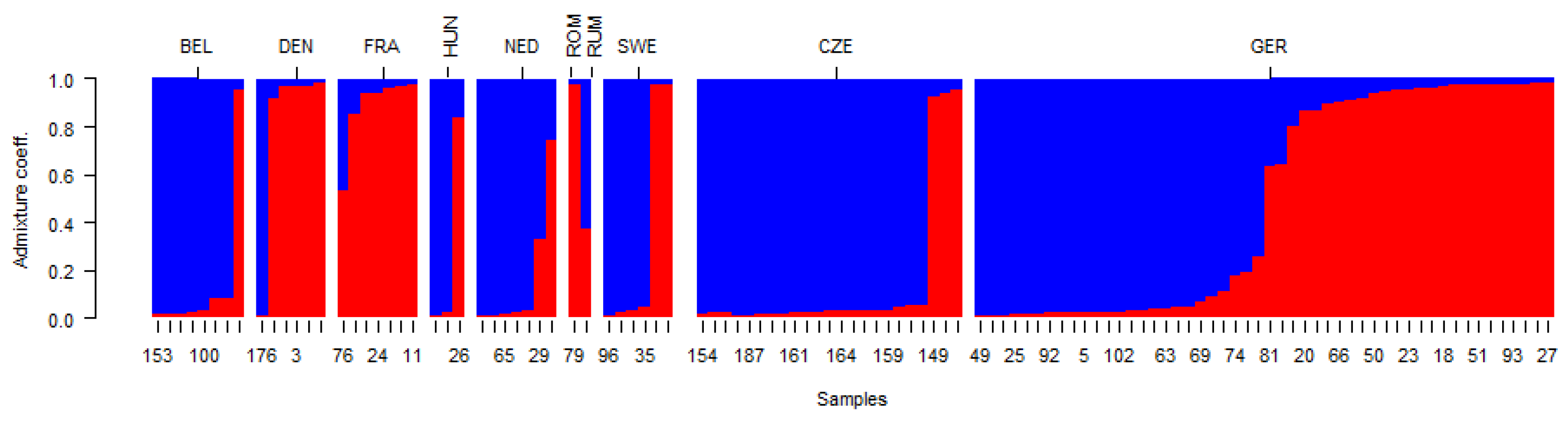
2. Results and Discussion
2.1. Microsatellite Polymorphism and Genetic Diversity of Genetic Resources
2.2. Genetic Structure of Yellow Mustard Varieties
3. Materials and Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hemingway, J.S. The mustard species: Condiment and food ingredient use and potential as oilseed crops. In Brassica Oilseeds Production and Utilization; Kimber, D., McGregor, D.I., Eds.; CAB International: Wallingford, UK, 1995; pp. 373–383. [Google Scholar]
- Ruan, S.F.; Wang, Z.X.; Xiang, S.J.; Chen, H.J.; Shen, Q.; Liu, L.; Wu, W.F.; Cao, S.W.; Wang, Z.W.; Yang, Z.J.; et al. Mechanisms of white mustard seed (Sinapis alba L.) volatile oils as transdermal penetration enhancers. Fitoterapia 2019, 138, 104195. [Google Scholar] [CrossRef] [PubMed]
- Bodnaryk, R.P.; Lamb, R.J. Mechanisms of resistance to the flea beetle, Phyllotreta cruciferae (Goeze), in mustard seedlings, Sinapis alba L. Can. J. Plant Sci. 1991, 71, 13–20. [Google Scholar] [CrossRef]
- Kumari, P.; Singh, K.P.; Rai, P.K. Draft genome of multiple resistance donor plant Sinapis alba: An insight into SSRs, annotations and phylogenetics. PLoS ONE 2020, 15, e0231002. [Google Scholar] [CrossRef] [PubMed]
- Valdes, Y.; Viaene, N.; Moens, M. Effects of yellow mustard amendments on the soil nematode community in a potato field with focus on Globodera rostochiensis. Appl. Soil Ecol. 2012, 59, 39–47. [Google Scholar] [CrossRef]
- Krasnodębska-Ostręga, B.; Sadowska, M.; Biaduń, E.; Mazur, R.; Kowalska, J. Sinapis alba as a useful plant in bioremediation—Studies of defense mechanisms and accumulation of As, Tl and PGEs. Int. J. Phytoremediat. 2022, 24, 1475–1490. [Google Scholar] [CrossRef] [PubMed]
- Masierowska, M. Floral nectaries and nectar production in brown mustard (Brassica juncea) and white mustard (Sinapis alba) (Brassicaceae). Plant Syst. Evol. 2003, 238, 97–107. [Google Scholar] [CrossRef]
- Katepa-Mupondwa, F.; Gugel, R.K.; Raney, J.P. Genetic diversity for agronomic, morphological and seed quality traits in Sinapis alba L. (yellow mustard). Can. J. Plant Sci. 2006, 86, 1015–1025. [Google Scholar] [CrossRef]
- Khoury, C.K.; Brush, S.; Costich, D.E.; Curry, H.A.; de Haan, S.; Engels, J.M.M.; Guarino, L.; Hoban, S.; Mercer, K.L.; Miller, A.J.; et al. Crop genetic erosion: Understanding and responding to loss of crop diversity. New Phytol. 2022, 233, 84–118. [Google Scholar] [CrossRef]
- Yu, J.K.; Chung, Y.S. Plant Variety Protection: Current Practices and Insights. Genes 2021, 12, 1127. [Google Scholar] [CrossRef]
- Govindaraj, M.; Vetriventhan, M.; Srinivasan, M. Importance of genetic diversity assessment in crop plants and its recent advances: An overview of its analytical perspectives. Genet. Res. Int. 2015, 2015, 431487. [Google Scholar] [CrossRef]
- Garland, S.; Curry, H.A. Turning promise into practice: Crop biotechnology for increasing genetic diversity and climate resilience. PLoS Biol. 2022, 20, e3001716. [Google Scholar] [CrossRef] [PubMed]
- McCallum, C.M.; Comai, L.; Greene, E.A.; Henikoff, S. Targeted screening for induced mutations. Nat. Biotechnol. 2000, 18, 455–457. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.; Howell, T.; Nitcher, R.; Missirian, V.; Watson, B.; Ngo, K.J.; Lieberman, M.; Fass, J.; Uauy, C.; Tran, R.K.; et al. Discovery of Rare Mutations in Populations: TILLING by Sequencing. Plant Physiol. 2011, 156, 1257–1268. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A. Genetic diversity and evolution of Brassica genetic resources: From morphology to novel genomic technologies—A review. Plant Genet. Resour. 2017, 15, 388–399. [Google Scholar] [CrossRef]
- Granot, D.; Shabelsky, E.; Schaffermann, D.; Yaniv, Z. Analysis of genetic variability between populations of Sinapis alba and the effect of cultivation on the variability. Acta Hortic. 1996, 407, 67–74. [Google Scholar] [CrossRef]
- Fu, Y.B.; Gugel, R.K.; Katepa-Mupondwa, F. Genetic diversity of Sinapis alba germplasm as revealed by AFLP markers. Plant Genet. Resour. 2006, 4, 87–95. [Google Scholar] [CrossRef]
- Yadav, M.; Rana, J.S. ISSR markers assisted studies on genetic diversity in Brassica juncea. Int. J. Pharma Bio Sci. 2012, 3, 402–411. [Google Scholar]
- Plieske, J.; Struss, D. Microsatellite markers for genome analysis in Brassica. I. Development in Brassica napus and abundance in Brassicaceae species. Theor. Appl. Genet. 2001, 102, 689–694. [Google Scholar] [CrossRef]
- Javidfar, F.; Cheng, B. Construction of a genetic linkage map and QTL analysis of erucic acid content and glucosinolate components in yellow mustard (Sinapis alba L.). BMC Plant Biol. 2013, 13, 142. [Google Scholar] [CrossRef]
- Fu, Y.B.; Cheng, B.; Peterson, G.W. Genetic diversity analysis of yellow mustard (Sinapis alba L.) germplasm based on genotyping by sequencing. Genet. Resour. Crop Evol. 2013, 61, 579–594. [Google Scholar] [CrossRef]
- Karp, A.; Edwards, K.J. Molecular Techniques in the Analysis of the Extent and Distribution of Genetic Diversity; International Plant Genetic Resources Institute: Rome, Italy, 1997; pp. 11–38. [Google Scholar]
- Singh, K.H.; Singh, L.; Parmar, N.; Kumar, S.; Nanjundan, J.; Singh, G.; Thakur, A.K. Molecular characterization and genetic diversity analysis in Indian mustard (Brassica juncea L. Czern & Coss.) varieties using SSR markers. PLoS ONE 2022, 17, e0272914. [Google Scholar] [CrossRef]
- Pradhan, A.; Nelson, M.N.; Plummer, J.A.; Cowling, W.A.; Yan, G. Characterization of Brassica nigra collections using simple sequence repeat markers reveals distinct groups associated with geographical location, and frequent mislabelling of species identity. Genome 2011, 54, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, M.; Feyissa, T.; Hailesilassie, T.; Kanagarajan, S.; Zhu, L.-H. Genetic diversity and population structure in ethiopian mustard (Brassica carinata A. Braun) as revealed by single nucleotide polymorphism markers. Genes 2023, 14, 1757. [Google Scholar] [CrossRef] [PubMed]
- Guichoux, E.; Lagashe, L.; Wagner, S.; Chaumeil, P.; Léger, P.; Lepais, O.; Lepoitteiven, C.; Malausa, T.; Revardel, E.; Salin, F.; et al. Current trends in microsatellite genotyping. Mol. Ecol. Resour. 2011, 11, 591–611. [Google Scholar] [CrossRef] [PubMed]
- Mondini, L.; Noorani, A.; Pagnotta, M. Assessing plant genetic diversity by molecular tools. Diversity 2009, 1, 19–35. [Google Scholar] [CrossRef]
- Varshney, R.K. Designing future crops: Genomics-assisted breeding comes of age. Trends Plant. Sci. 2021, 26, 631–649. [Google Scholar] [CrossRef]
- Olsson, G. Self-incompatibility and outcrossing in rape and white mustard. Hereditas 1960, 46, 241–252. [Google Scholar] [CrossRef]
- Treccarichi, S.; Ben Ammar, H.; Amari, M.; Cali, R.; Tribulato, A.; Branca, F. Molecular markers for detecting inflorescence size of Brassica oleracea L. crops and B. oleracea complex species (n = 9) useful for breeding of broccoli (B. oleracea var. italica) and cauliflower (B. oleracea var. botrytis). Plants 2023, 12, 407. [Google Scholar] [CrossRef]
- Cui, X.; Dong, Y.; Hou, X.; Cheng, Y.; Zhang, J.; Jin, M. Development and characterization of microsatellite markers in Brassica rapa ssp. chinensis and transferabillity among related species. Agric. Sci. China 2008, 7, 19–31. [Google Scholar] [CrossRef]
- Havlickova, L.; Jozova, E.; Rychla, A.; Klima, M.; Kucera, V.; Curn, V. Genetic diversity assessment in winter oilseed rape (Brassica napus L.) collection using AFLP, ISSR and SSR markers. Czech J. Genet. Plant Breed. 2014, 50, 216–225. [Google Scholar] [CrossRef]
- Vazhakkannadi, V.; Singh, N.; Vasudev, S.; Yadava, D.; Kumar, S.; Naresh, S.; Bhat, S.; Prabhu, K. Assessment of genetic diversity in Brassica juncea (Brassicaceae) genotypes using phenotypic differences and SSR markers. Rev. Biol. Trop. 2013, 61, 1919–1934. [Google Scholar]
- Khedikar, Y.; Clarke, W.; Chen, L.; Higgins, E.; Kagale, S.; Koh, C.; Bennett, R.; Parkin, I. Narrow genetic base shapes population structure and linkage disequilibrium in an industrial oilseed crop, Brassica carinata A. Braun. Sci. Rep. 2020, 10, 12629. [Google Scholar] [CrossRef] [PubMed]
- Sobotka, R.; Dolanská, L.; Čurn, V.; Ovesná, J. Fluorescence-based AFLPs occur as the most suitable marker system for oilseed rape cultivar identification. J. Appl. Genet. 2004, 45, 161–173. [Google Scholar] [PubMed]
- Cowling, W.A. Genetic diversity in Australian canola and implications for crop breeding for changing future environments. Field Crops Res. 2007, 104, 103–111. [Google Scholar] [CrossRef]
- Chikkaputtaiah, C.; Debbarma, J.; Baruah, I.; Havlickova, L.; Deka Boruah, H.P.; Curn, V. Molecular genetics and functional genomics of abiotic stress-responsive genes in oilseed rape (Brassica napus L.): A review of recent advances and future. Plant Biotechnol. Rep. 2017, 11, 365–384. [Google Scholar] [CrossRef]
- Kolář, F.; Fér, T.; Štěch, M.; Trávníček, P.; Dušková, E.; Schönswetter, P.; Suda, J. Bringing together evolution on serpentine and polyploidy: Spatiotemporal history of the diploid-tetraploid complex of Knautia arvensis (Dipsacaceae). PLoS ONE 2012, 7, e39988. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stehens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Bird, C.; Karl, S.A.; Smouse, P.E.; Toonen, R.J. Detecting and measuring genetic differentiation. In Phylogeography and Population Genetics in Crustacea; Koenemann, S., Held, C., Schubart, C., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 31–55. [Google Scholar] [CrossRef]
- Yao, Q.L.; Chen, F.B.; Fang, P.; Zhou, G.F.; Fan, Y.H.; Zhang, Z.R. Genetic diversity of Chinese vegetable mustard (Brassica juncea Coss) landraces based on SSR data. Biochem. Syst. Ecol. 2012, 45, 41–48. [Google Scholar] [CrossRef]
- Ofori, A.; Becker, H.C.; Kopisch-Obuch, F.J. Effect of crop improvement on genetic diversity in oilseed Brassica rapa (turnip-rape) cultivars, detected by SSR markers. J. Appl. Genet. 2008, 49, 207–212. [Google Scholar] [CrossRef]
- Abdel-haleem, H.; Luo, Z.; Szczepanek, A. Genetic diversity and population structure of the USDA collection of Brassica juncea L. Ind. Crops Prod. 2022, 187, 115379. [Google Scholar] [CrossRef]
- Sanchez, D.; Sadoun, S.B.; Mary-Huard, T.; Allier, A.; Moreau, L.; Charcosset, A. Improving the use of plant genetic resources to sustain breeding programs’ efficiency. Proc. Natl. Acad. Sci. USA 2023, 120, e2205780119. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, H.; Bhanu, A.; Srivastava, K.; Singh, M.; Shreya; Hemantaranjan, A. Assessment of genetic diversity in crop plants: An overview. Adv. Plants Agric. Res. 2017, 7, 279–286. [Google Scholar] [CrossRef]
- Begna, T. Role and economic importance of crop genetic diversity in food security. Int. J. Agric. Sci. Food Technol. 2021, 7, 164–169. [Google Scholar] [CrossRef]
- Kurowska, M.; Daszkowska-Golec, A.; Gruszka, D.; Marzec, M.; Szurman, M.; Szarejko, I.; Maluszynski, M. TILLING: A shortcut in functional genomics. J. Appl. Genet. 2011, 52, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Mba, C. Induced mutations unleash the potentials of plant genetic resources for food and agriculture. Agronomy 2013, 3, 200–231. [Google Scholar] [CrossRef]
- Doyle, J. DNA Protocols for Plants. In Molecular Techniques in Taxonomy; Hewitt, G.M., Johnston, A.W.B., Young, J.P.W., Eds.; Springer: Berlin/Heidelberg, Germany, 1991; pp. 283–293. [Google Scholar] [CrossRef]
- Louarn, S.; Torp, A.; Holme, I.; Andersen, S.B.; Jensen, B. Database derived microsatellite markers (SSRs) for cultivar differentiation in Brassica oleracea. Genet. Resour. Crop Evol. 2007, 54, 1717–1725. [Google Scholar] [CrossRef]
- Jozová, E.; Čurn, V. Microsatellite genotyping with 4-color fluorescent detection using multiple-tailed primers in oilseed rape. Úroda 2014, 12, 191–194. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 30 October 2022).
- Jombart, T.; Ahmed, I. Adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Paradis, E. pegas: An R package for population genetics with an integrated–modular approach. Bioinformatics 2010, 26, 419–420. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microsatellite Marker | Number of Individuals | PIC | Na (Number of Alleles) | Number of Alleles in Population | Length of Alleles in Bp | Median and Max. Frequency of Alleles | Ho (Observed Heterozygosity) | He (Nei, 1973; Expected Heterozygosity) | Fis (Inbreeding Coefficient) | 1-D (Simpson Index) | Hexp (Nei. 1978; Gene Diversity) | Evenness |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BoREM1b _A | 198 | 0.19 | 2 | 209 | 173–175 | 104.5; 187 | 0.12 | 0.11 | −0.06 | 0.11 | 0.11 | 0.50 |
| BoREM1b _B | 198 | 0.00 | 1 | 3 | 181 | 3; 3 | 0 | 0 | - | - | - | |
| BoREM1b _C | 198 | 0.00 | 1 | 3 | 187 | 3; 3 | 0 | 0 | - | - | - | |
| BolAB19TF | 198 | 0.00 | 1 | 1 | 314 | 1; 1 | 0 | 0 | - | - | - | |
| BoPC34 | 198 | 0.51 | 3 | 323 | 149–151 | 148; 172 | 0.73 | 0.50 | −0.45 | 0.50 | 0.50 | 0.94 |
| P381 | 198 | 0.50 | 2 | 21 | 213–229 | 105; 11 | 0.05 | 0.50 | 0.90 | 0.50 | 0.51 | 1.00 |
| D3 _A | 198 | 0.65 | 6 | 342 | 155–163 | 33.5; 149 | 0.83 | 0.67 | −0.24 | 0.67 | 0.67 | 0.85 |
| D3 _B | 198 | 0.44 | 5 | 101 | 167–171 | 10; 73 | 0.20 | 0.39 | 0.50 | 0.38 | 0.68 | 0.54 |
| P7 | 198 | 0.27 | 6 | 119 | 141–156 | 3.5; 101 | 0.11 | 0.21 | 0.49 | 0.21 | 0.21 | 0.39 |
| P9 _B | 198 | 0.38 | 3 | 184 | 125–138 | 32; 141 | 0.28 | 0.29 | 0.05 | 0.27 | 0.27 | 0.54 |
| P9 _C | 198 | 0.35 | 4 | 231 | 145–160 | 23; 184 | 0.25 | 0.23 | −0.06 | 0.23 | 0.24 | 0.49 |
| P30 _B | 198 | 0.32 | 2 | 5 | 166–174 | 2.5; 4 | 0 | 0.32 | 1.00 | 0.32 | 0.36 | 0.72 |
| P30 _C | 198 | 0.00 | 1 | 2 | 192 | 2; 2 | 0 | 0 | - | - | - | |
| P35 _A | 198 | 0.52 | 6 | 109 | 149–161 | 3.5; 68 | 0.53 | 0.47 | −0.12 | 0.46 | 0.47 | 0.63 |
| P35 _B | 198 | 0.00 | 1 | 1 | 173 | 1; 1 | 0 | 0 | - | - | - |
| df | SS | MS | Genetic Variance Estimates (Sigma) | Proportion of Variance (%) | Phi (Φ) | p- Values | |
|---|---|---|---|---|---|---|---|
| Between types of genetic resources | 2 | 19.41 | 9.705 | 0.143 | 4.602 | ΦCT = 0.0460 | <0.0001 |
| Between genotypes within types of genetic resources | 31 | 123.07 | 3.970 | 0.257 | 8.272 | ΦSC = 0.0867 | <0.0001 |
| Within genetic resources | 164 | 414.12 | 2.707 | 2.707 | 87.126 | ΦST = 0.1287 | 0.0028 |
| Total | 197 | 556.60 | 2.992 | 3.107 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jozová, E.; Rost, M.; Rychlá, A.; Stehlíková, D.; Pudhuvai, B.; Hejna, O.; Beran, P.; Čurn, V.; Klíma, M. Microsatellite Markers: A Tool to Assess the Genetic Diversity of Yellow Mustard (Sinapis alba L.). Plants 2023, 12, 4026. https://doi.org/10.3390/plants12234026
Jozová E, Rost M, Rychlá A, Stehlíková D, Pudhuvai B, Hejna O, Beran P, Čurn V, Klíma M. Microsatellite Markers: A Tool to Assess the Genetic Diversity of Yellow Mustard (Sinapis alba L.). Plants. 2023; 12(23):4026. https://doi.org/10.3390/plants12234026
Chicago/Turabian StyleJozová, Eva, Michael Rost, Andrea Rychlá, Dagmar Stehlíková, Baveesh Pudhuvai, Ondřej Hejna, Pavel Beran, Vladislav Čurn, and Miroslav Klíma. 2023. "Microsatellite Markers: A Tool to Assess the Genetic Diversity of Yellow Mustard (Sinapis alba L.)" Plants 12, no. 23: 4026. https://doi.org/10.3390/plants12234026