
Detecting Tomato Leaf Curl New Delhi Virus Causing Ridge Gourd Yellow Mosaic Disease, and Other Begomoviruses by Antibody-Based Methods

 and
and
Abstract
1. Introduction
2. Results
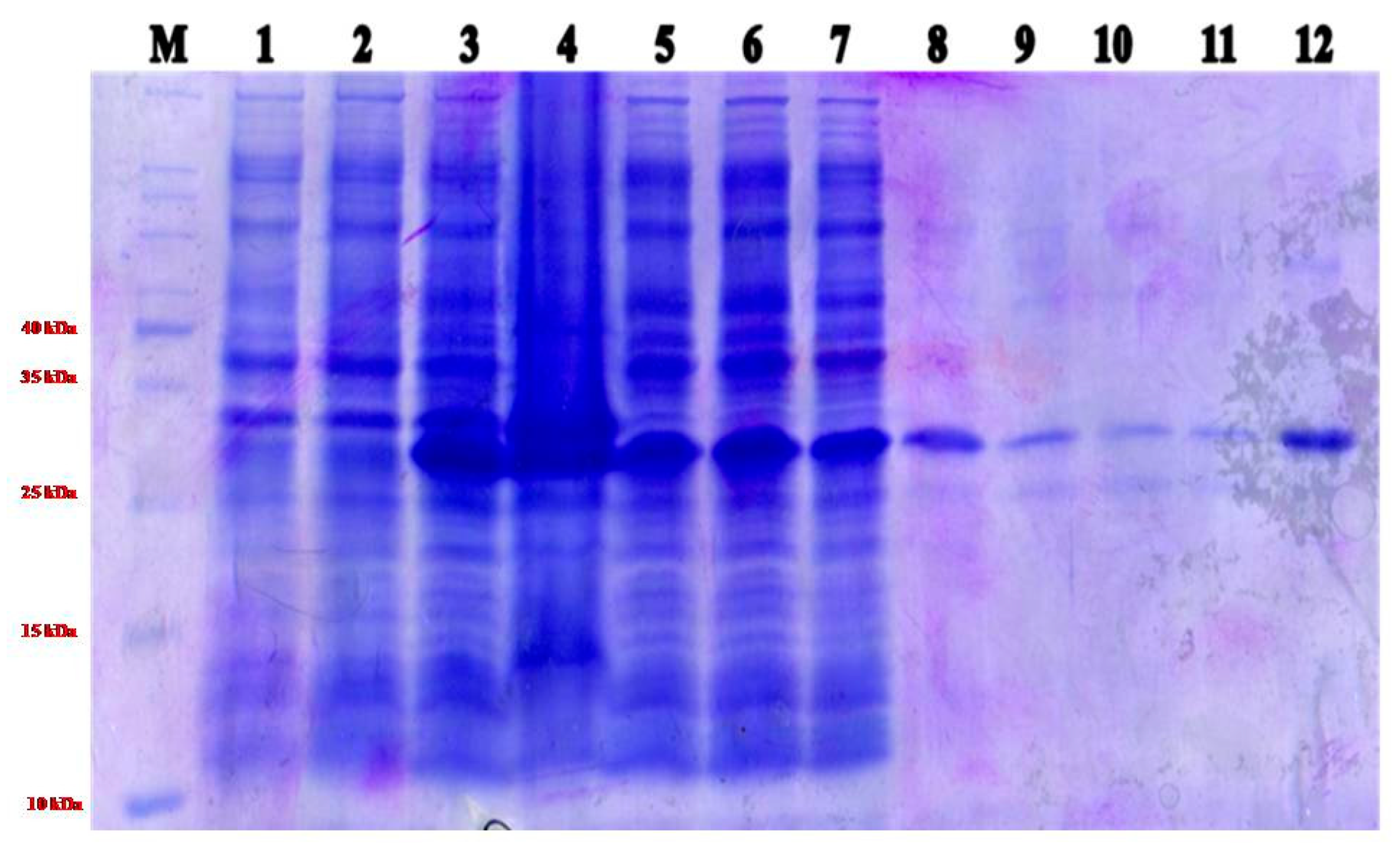
2.1. Expression ToLCNDV- Rg-CP Gene and Purification of Recombinant Protein
2.2. Sensitivity of ToLCNDV-Rg-CP-PAb to Recombinant Protein
2.3. DIBA
2.4. WB
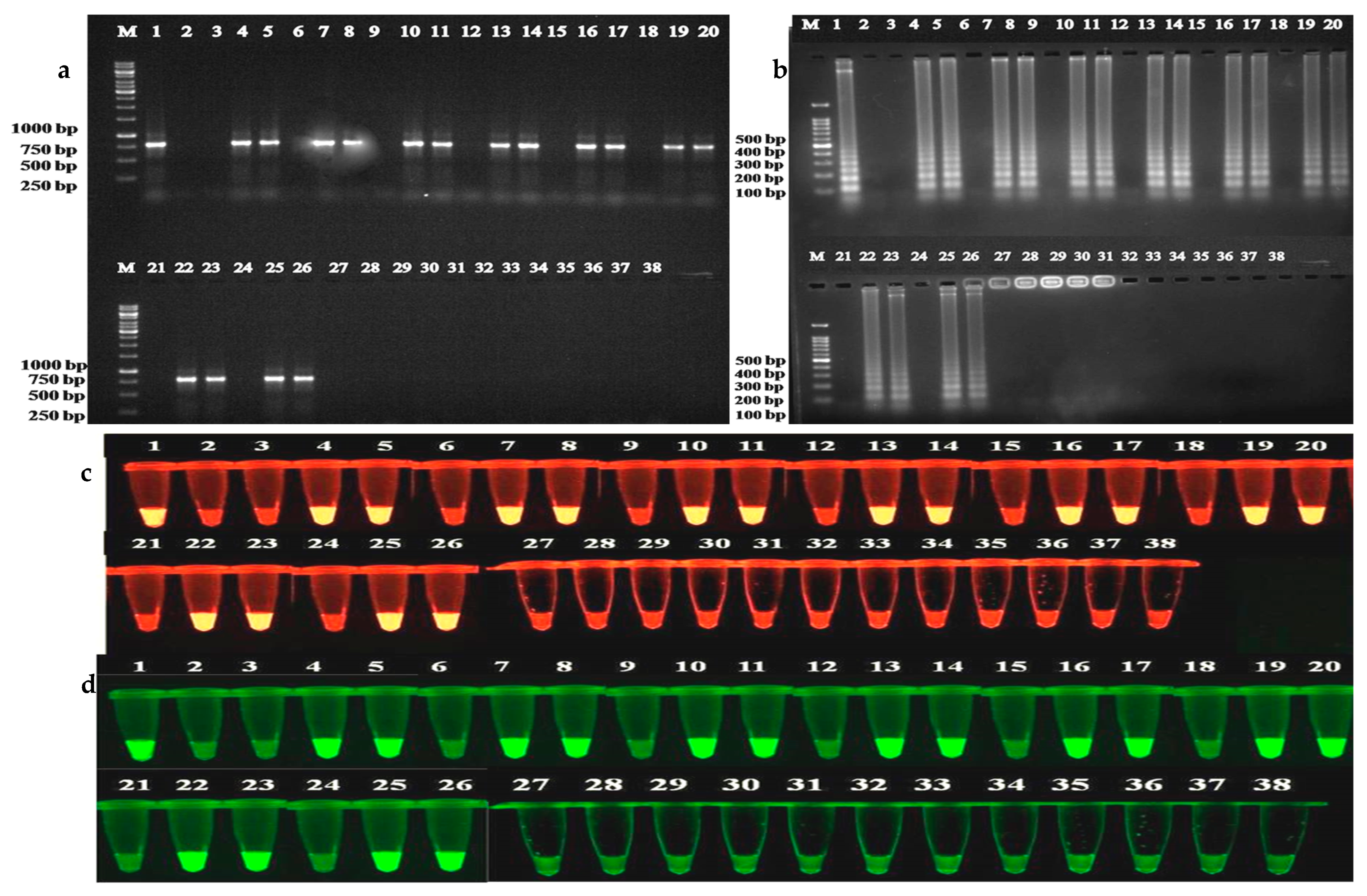
2.5. Immunocapture Assays (IC-PCR and IC-LAMP)
2.6. Determining the Efficacy of ToLCNDV-Rg-CP-PAbs in Immmunocapture Assays
2.7. Broad-Spectrum Detection of Begomoviruses Using ToLCNDV-Rg-CP-PAb
2.8. Sequence Analysis of ToLCNDV-Rg Isolate CP Gene with the Other Begomoviruses
3. Discussion
4. Materials and Methods
4.1. Virus Samples and DNA Isolation
4.2. Construction of ToLCNDV-Rg-CP Plasmid, Expression, and Purification of Protein
4.3. Protein Separation by Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE)
4.4. Sequence Analysis of ToLCNDV-Rg-CP
4.5. Immunization of Rabbit and Serum Collection
4.6. Antibody Tests
4.6.1. Direct Antigen Coating Enzyme-Linked Immuno Sorbent Assay
4.6.2. Dot Immuno Binding Assay
4.6.3. Western Blotting
4.6.4. Immunocapture Polymerase Chain Reaction and Immunocapture Loop Mediated Isothermal Amplification
4.7. Broad-Specificity Analysis of the ToLCNDV-Rg-CP-PAb for the Detection of Begomoviruses Infecting Other Crops
4.7.1. Virus Infected Samples and DNA Isolation
4.7.2. Detection of Begomoviruses in Different Crop Plants
4.7.3. Sequence Analysis of Begomoviruses Associated with Other Crops
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Chakravarthy, M.L. Monograph of Indian cucurbitaceous (Taxonomy and distribution). Rec. Bot. Distrib. India 1959, 17, 6–7. [Google Scholar]
- Gopalakrishnan, T.R. Vegetable Crops; Peter, K.V., Ed.; Horticulture Science Series; New India Publishing Agency: New Delhi, India, 2007; p. 343. [Google Scholar]
- Patil, C.V.; Ramdas, S.V.; Premchand, U.; Shankarappa, K.S. Survey, symptomatology, transmission, host range and characterization of begomovirus associated with yellow mosaic disease of ridge gourd from southern India. Virus Dis. 2017, 28, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Harrison, B.D.; Robinson, D.J. Natural genomic and antigenic variation in whitefly-transmitted geminiviruses (Begomoviruses). Ann. Rev. Phytopathol. 1999, 37, 369–398. [Google Scholar] [CrossRef] [PubMed]
- Zerbini, F.M.; Briddon, R.W.; Idris, A.; Martin, D.P.; Moriones, E.; Navas-Castillo, J.; Varsani, A. ICTV virus taxonomy profile:Geminiviridae. J. Gen. Virol. 2017, 98, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Kumar, M. Tomato leaf curl New Delhi virus (Geminiviridae). In Encyclopedia of Virology, 4th ed.; Bamford, D.H., Zuckerman, M., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 749–760. [Google Scholar]
- Gemert, J.; Martijn, S.; Candresse, T.; Bottex, B.; Sybren, A.D. Pest survey card on tomato leaf curl New Delhi virus. EFSA Support. Publ. 2020, 1904, 7–17. [Google Scholar]
- Juarez, M.; Rabadan, M.P.; Martínez, L.D.; Tayahi, M.; Grande-Perez, A.; Gomez, P. Natural hosts and genetic diversity of the emerging tomato leaf curl New Delhi virus in Spain. Front. Microbiol. 2019, 10, 140–145. [Google Scholar] [CrossRef]
- Venkatasalam, E.P.; Singh, S.; Sivalingam, P.N.; Malathi, V.G.; Garg, I.D. Polymerase chain reaction and nucleic acid spot hybridization detection of begomovirus (es) associated with apical leaf curl disease of potato. Arch. Phytopathol. Plant Protect. 2011, 44, 987–992. [Google Scholar] [CrossRef]
- Kumar, R.; Tiwari, R.K.; Jeevalatha, A.; Kaundal, P.; Sharma, S.; Chakrabarti, S.K. Potato viruses and their diagnostic techniques: An overview. J. Pharmacogn. Phytochem. 2019, 8, 1932–1944. [Google Scholar]
- Naganur, P.; Premchand, U.; Shankarappa, K.S.; Mesta, R.K.; Manjunatha, C.; Patil, C.V. Development of a loop-mediated isothermal amplification assay for detection of tomato leaf curl New Delhi virus in ridge gourd [Luffa acutangula (L.) Roxb.]. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 2282–2295. [Google Scholar] [CrossRef]
- Venkataravanappa, V.; Ashwathappa, K.V.; Reddy, C.N.L.; Shankarappa, K.S.; Reddy, M.K. Characterization of tomato leaf curl New Delhi virus associated with leaf curl and yellowing disease of watermelon and development of LAMP assay for its detection. 3 Biotech 2020, 10, 282–285. [Google Scholar] [CrossRef]
- Cancino, M.; Abouzid, A.M.; Morales, F.J.; Purcifull, D.E.; Polston, J.E.; Hiebert, E. Generation and characterization of three monoclonal antibodies useful in detecting and distinguishing bean golden mosaic virus isolates. Phytopathology 1995, 85, 484–490. [Google Scholar] [CrossRef]
- Abouzid, A.M.; Freitas-Astua, J.; Purcifull, D.E.; Polston, J.E.; Beckham, K.A.; Crawford, W.E.; Petersen, M.A.; Peyser, B.; Patte, C.; Hiebert, E. Serological studies using polyclonal antisera prepared against the viral coat protein of four begomoviruses expressed in Escherichia coli. Plant Dis. 2002, 86, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Colariccio, A.; Lovisolo, O.; Boccardo, G.; Chagas, C.M.; d’Aquilio, M.; Rossetti, V. Preliminary purification and double stranded RNA analysis of citrus leprosis virus. Int. Organ. Citrus Virol. Conf. Proc. 1957-2010 2000, 14, 159–163. [Google Scholar]
- Ling, K.S.; Zhu, H.Y.; Jiang, Z.Y.; Gonsalves, D. Effective application of DAS-ELISA for detection of grapevine leafroll associated closterovirus-3 using a polyclonal antiserum developed from recombinant coat protein. Eur. J. Plant Pathol. 2000, 106, 301–309. [Google Scholar] [CrossRef]
- Ling, K.S.; Zhu, H.Y.; Petrovic, N.; Gonsalves, D. Serological detection of grapevine leafroll virus 2 using an antiserum developed against the recombinant coat protein. J. Phytopathol. 2007, 155, 65–69. [Google Scholar] [CrossRef]
- Hull, R. Plant Virology, 5th ed.; Academic Press: Cambridge, MA, USA, 2013; p. 1118. [Google Scholar]
- Alves-Junior, M.; Marraccini, F.M.; MeloFilho, P.A.; Dusi, A.N.; Pio-Ribeiro, G.; Ribeiro, B.M. Recombinant expression of garlic virus C (GARV-C) capsid protein in insect cells and its potential for the production of specific antibodies. Microbiol. Res. 2008, 163, 354–361. [Google Scholar] [CrossRef]
- Canto, T.; Prior, D.A.M.; Hellward, K.H.; Oparka, K.J.; Palukaitis, P. Characterization of cucumber mosaic virus IV. Movement protein and coat protein are both essential for cell-to-cell movement of cucumber mosaic virus. Virology 1997, 237, 237–248. [Google Scholar] [CrossRef]
- Cerovska, N.; Filigarova, M.; Pecenkova, T. Production of polyclonal antibodies to a recombinant potato moptop virus non-structural triple gene block protein 1. J. Phytopathol. 2006, 154, 422–427. [Google Scholar] [CrossRef]
- Singh, S.; Nirmalkar, V.K.; Awasthi, L.P. Recent advances in begomovirus research in India. In Applied Plant Virology; Awasthi, L.P., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 493–513. [Google Scholar]
- Almasi, M.A.; Dehabadi, S.H.; Eftekhari, Z. Immunocapture loop mediated isothermal amplification for rapid detection of tomato yellow leaf curl virus (TYLCV) without DNA extraction. J. Plant Pathol. Microbiol. 2013, 4, 185. [Google Scholar]
- Martin, S.; Alioto, D.; Milne, R.G. Detection of citrus psorosis virus in field trees by direct tissue blot immunoassay in comparison with ELISA, symptomatology, biological indexing and cross-protection tests. Plant Pathol. 2002, 51, 134–141. [Google Scholar] [CrossRef]
- Agarwal, G.; Kavalappara, S.R.; Gautam, S.; Silva, A.D.; Simmons, A.; Srinivasan, R.; Dutta, B. Field screen and genotyping of Phaseolus vulgaris against two begomoviruses in Georgia, USA. Insects 2021, 12, 49. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Chang, Y.C. Performances and application of antisera produced by recombinant capsid proteins of cymbidium mosaic virus and odontoglossum ringspot virus. Eur. J. Plant Pathol. 2008, 122, 297–306. [Google Scholar] [CrossRef]
- Carson, M.; Johnson, D.H.; McDonald, H.; Brouillette, C.; Delucas, L.J. His-tag impact on structure. Acta Crystallogr. Sect. D Biol. Crystallogr. 2007, 63, 295–301. [Google Scholar] [CrossRef]
- Rajasulochana, P.; Dhamotharan, R.; Srinivasulu, P. Comparison of DAC-ELISA for the detection of cucumber mosaic and banana streak viruses infecting banana. J. Am. Sci. 2008, 4, 1003–1545. [Google Scholar]
- Sharma, K.; Misra, R.S. Molecular approaches towards analysing the viruses infecting maize (Zea mays L.). J. Gen. Mol. Virol. 2011, 3, 69–85. [Google Scholar]
- Fukuta, S.; Iida, T.; Mizukami, Y.; Ishida, A.; Ueda, J.; Kanbe, M.; Ishimoto, Y. Detection of Japanese yam mosaic virus by RT-LAMP. Arch. Virol. 2003, 148, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Marchant, W.G.; Gautam, S.; Hutton, S.F.; Srinivasan, R. Tomato yellow leaf curl virus-resistant and -susceptible tomato genotypes similarly impact the virus population genetics. Front. Plant Sci. 2020, 11, 599697. [Google Scholar] [CrossRef]
- Marchant, W.; Gautam, S.; Dutta, B.; Srinivasan, R. Whitefly-mediated transmission and subsequent acquisition of highly similar and naturally occurring tomato yellow leaf curl virus variants. Phytopathology 2022, 112, 720–728. [Google Scholar] [CrossRef]
- Harrison, B.D.; Zhou, X.; Otim-Nape, G.M.; Liu, Y.; Robinson, D.J. Role of a novel type of double infection in the geminivirus-induced epidemic of severe cassava mosaic in Uganda. Ann. Appl. Biol. 1997, 131, 437–448. [Google Scholar] [CrossRef]
- Lodhi, M.A.; Ye, G.N.; Weeden, N.F.; Reisch, B. A simple and efficient method for DNA extraction from grapevine cultivars and Vitis species. Plant Mol. Biol. Rep. 1994, 12, 6–13. [Google Scholar] [CrossRef]
- Maruthi, M.N.; Colvin, J.; Seal, S.E.; Gibson, G.; Cooper, J. Coadaptation between cassava mosaic geminiviruses and their local vector populations. Virus Res. 2002, 86, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Jagannath, M.R.; Kesavulu, M.M.; Deepa, R.; Sastri, P.N.; Kumar, S.S.; Suguna, K.; Rao, C.D. N-and C-terminal cooperation in rotavirus enterotoxin: Novel mechanism of modulation of the properties of a multifunctional protein by a structurally and functionally overlapping conformational domain. J. Virol. 2006, 80, 412–425. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Ann. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the bacteriophage. Nature 1970, 277, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Clark, M.F.; Bar-Joseph, M. Enzyme immunosorbent assays in plant virology. In Methods in Virology; Maramorosch, K., Koprowski, H., Eds.; Academic Press: New York, NY, USA, 1984; pp. 51–85. [Google Scholar]
- Hawkes, R.; Niday, E.; Gordon, J. A dot immuno binding assay for monoclonal and other antibodies. Ann. Biochem. 1982, 119, 142–147. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of protein from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef]
- Wyatt, S.D.; Brown, J.K. Detection of subgroup III geminivirus isolates in leaf extracts by degenerate primers and polymerase chain reaction. Phytopathology 1996, 86, 1288–1293. [Google Scholar] [CrossRef]
- Venkataravanappa, V.; Lakshminarayanareddy, C.N.; Salil, J.; Krishnareddy, M. Molecular characterization of distinct bipartite begomovirus infecting bhendi (Abelomoschus esculentus L.) in India. Virus Genes 2012, 44, 522–535. [Google Scholar] [CrossRef]
- Brown, J.K.; Zerbini, F.M.; Navas-Castillo, J.; Moriones, E.; Ramos-Sobrinho, R.; Silva, J.C.F.; Fiallo-Olivé, E.; Briddon, R.W.; Hernandez-Zepeda, C.; Idris, A.; et al. Revision of begomovirus taxonomy based on pairwise sequence comparisons. Arch. Virol. 2015, 160, 1593–1619. [Google Scholar] [CrossRef]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl No | Begomovirus | Hosts |
|---|---|---|
| 1 | Tomato leaf curl New Delhi virus (ToLCNDV) | Bitter gourd, bottle gourd, cucumber, ridge gourd, snake gourd, and pumpkin |
| 2 | Tomato leaf curl Palampur virus (ToLCPalV) | Watermelon |
| 3 | Squash leaf curl China virus (SLCCNV) | Pumpkin |
| 4 | Chilli leaf curl virus (ChiLCV) | Chilli and capsicum |
| 5 | Mungbean yellow mosaic India virus (MYMIV) | Cowpea and French bean |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naganur, P.; Shankarappa, K.S.; Mesta, R.K.; Rao, C.D.; Venkataravanappa, V.; Maruthi, M.N.; Reddy, L.R.C.N. Detecting Tomato Leaf Curl New Delhi Virus Causing Ridge Gourd Yellow Mosaic Disease, and Other Begomoviruses by Antibody-Based Methods. Plants 2023, 12, 490. https://doi.org/10.3390/plants12030490
Naganur P, Shankarappa KS, Mesta RK, Rao CD, Venkataravanappa V, Maruthi MN, Reddy LRCN. Detecting Tomato Leaf Curl New Delhi Virus Causing Ridge Gourd Yellow Mosaic Disease, and Other Begomoviruses by Antibody-Based Methods. Plants. 2023; 12(3):490. https://doi.org/10.3390/plants12030490
Chicago/Turabian StyleNaganur, Priya, Kodegandlu Subbanna Shankarappa, Raghavendra K. Mesta, Chilakalapudi Durga Rao, Venkataravanappa Venkataravanappa, Midatharahally Narasegowda Maruthi, and Lakshminarayana Reddy C. Narasimha Reddy. 2023. "Detecting Tomato Leaf Curl New Delhi Virus Causing Ridge Gourd Yellow Mosaic Disease, and Other Begomoviruses by Antibody-Based Methods" Plants 12, no. 3: 490. https://doi.org/10.3390/plants12030490
APA StyleNaganur, P., Shankarappa, K. S., Mesta, R. K., Rao, C. D., Venkataravanappa, V., Maruthi, M. N., & Reddy, L. R. C. N. (2023). Detecting Tomato Leaf Curl New Delhi Virus Causing Ridge Gourd Yellow Mosaic Disease, and Other Begomoviruses by Antibody-Based Methods. Plants, 12(3), 490. https://doi.org/10.3390/plants12030490