
Environmental Filtering Drives Fungal Phyllosphere Community in Regional Agricultural Landscapes
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Height of Wheat Plants
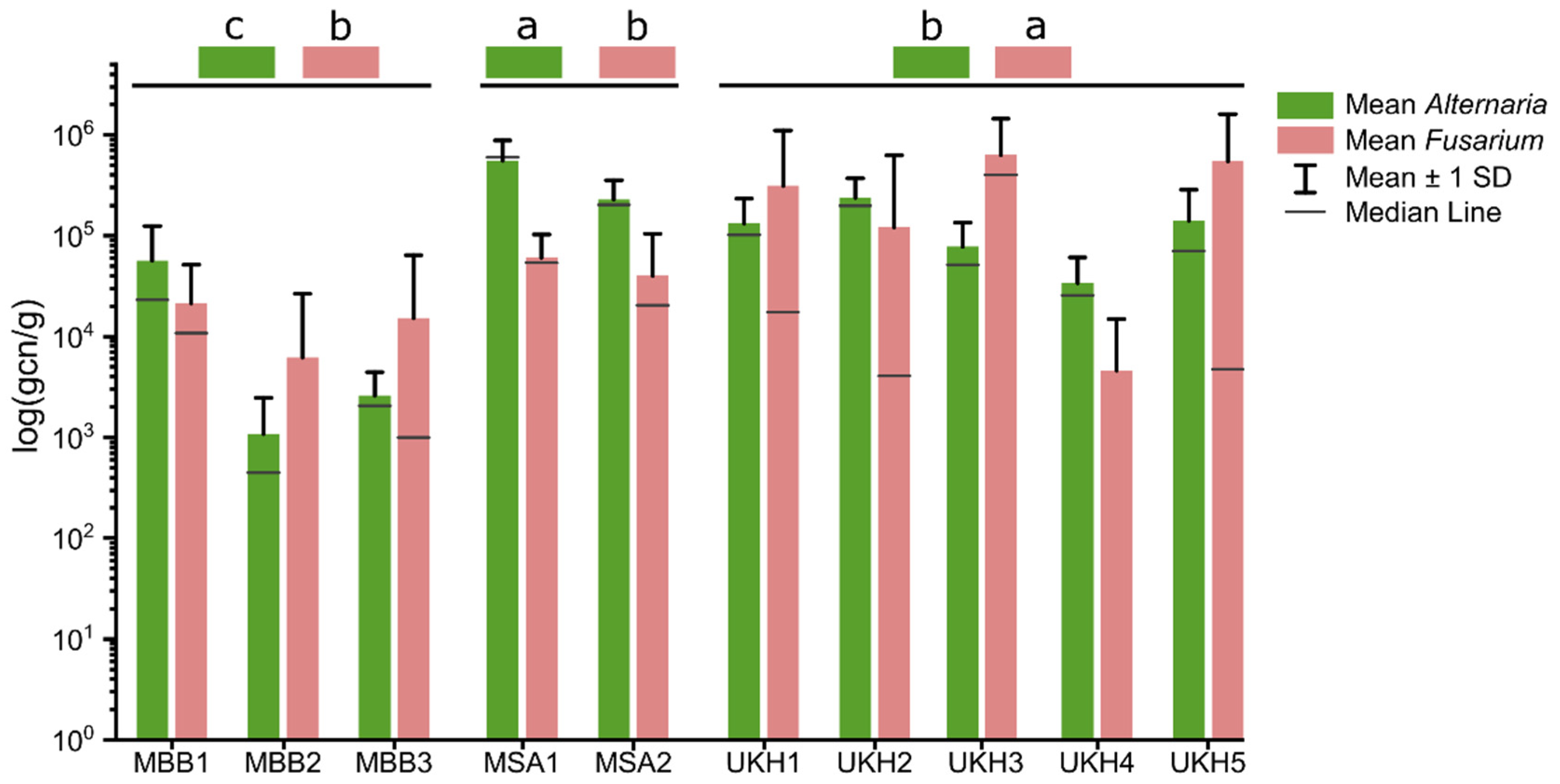
2.2. Quantitative Abundances of Fusaria and Alternaria
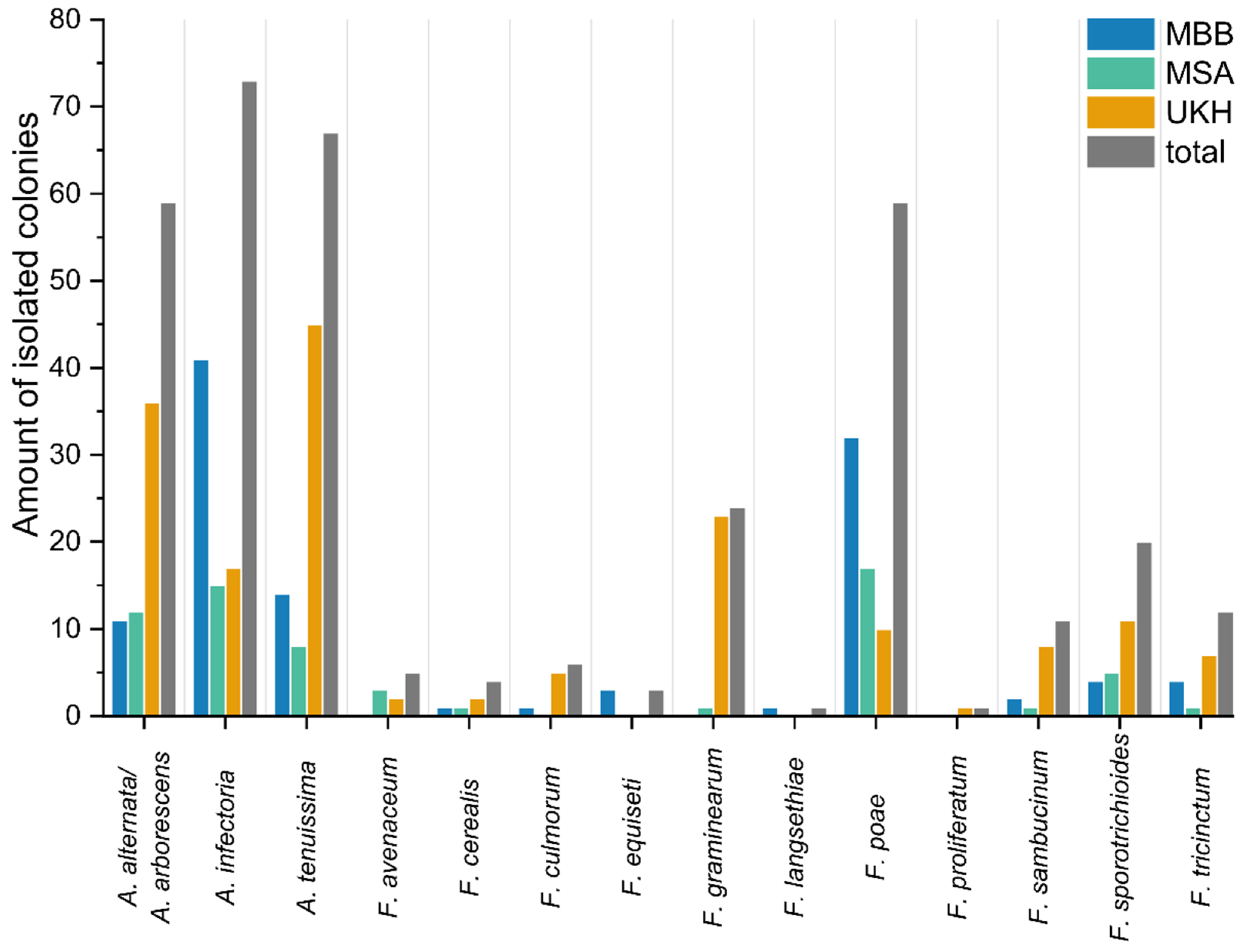
2.3. Culture-Based Identification of Fusaria and Alternaria
2.4. NGS Data Analysis (ITS)
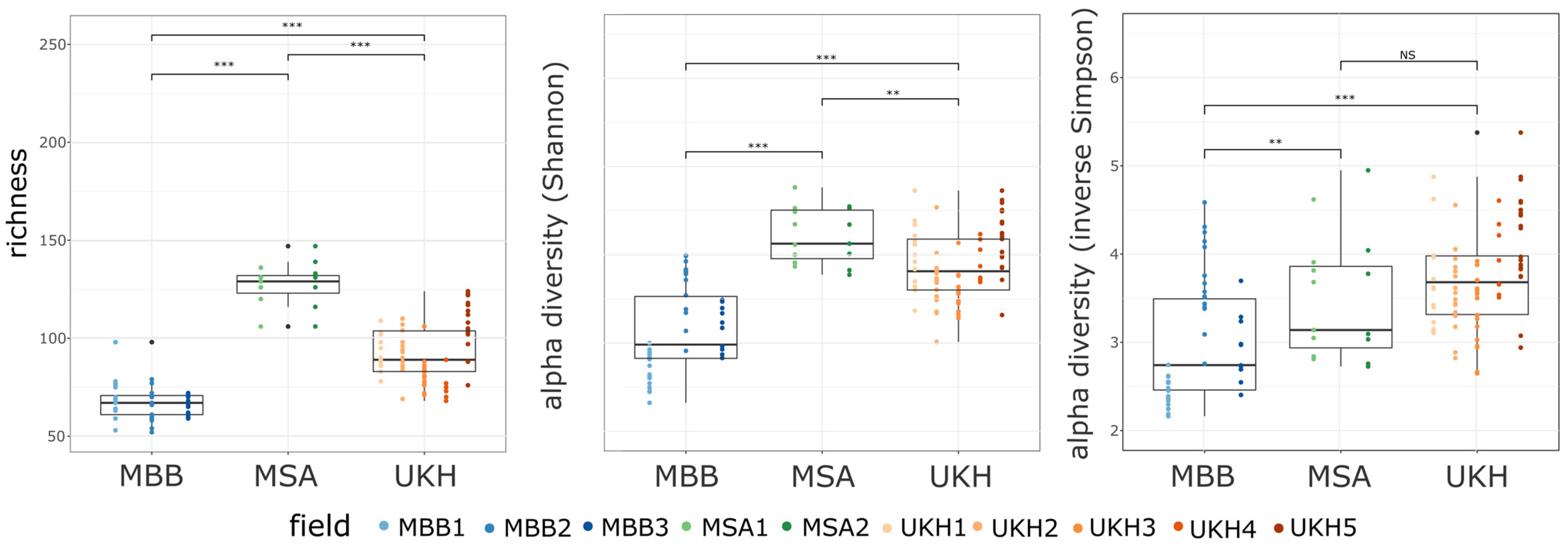
2.4.1. Alpha Diversity
2.4.2. Beta Diversity
2.4.3. Taxonomic Composition
2.4.4. Correlation of Fungal Community Composition with the Quantitative Abundance of Fusarium and Alternaria across and within Regions
3. Discussion
3.1. Differences between and within Regions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Average Characteristics | MBB | MSA | UKH1–UKH3 | UKH4–UKH5 |
|---|---|---|---|---|
| Soil region | Old moraine landscapes | Young moraine landscapes | Young moraine landscapes | Young moraine landscapes |
| Soil type | Chernozem | Albeluvisol | Brown soils– para brown soils | Brown soils–para brown soils |
| SQR | ≥85 | <35–<50 | 50–<70 | 50–<70 |
| Arable yield potential | very high | very low | medium | medium |
| TWI | 8.3 | 7.8 | 7.8 | 7.0 |
| Precipitation sum (mm) (Jan–July 2020) | 277 | 291 | 233 | 262 |
| Mean temperature (°C) (Jan–July 2020) | 10.8 | 10.5 | 10.0 | 9.9 |
| Height of wheat plants (cm) | 85 | 73 | 88 | 91 |
| qPCR fusaria (gcn/g) | 14,220 | 50,256 | 353,505 | 275,063 |
| qPCR alternaria (gcn/g) | 19,997 | 391,773 | 149,091 | 87,041 |
| Dominant Fusarium | F. poae | F. poae | F. graminearum | F. graminearum, F. tricinctum |
| Taxonomic biomarker | Sporidiobolales | Capnodiales, Pleosporales | Hypocreales | Hypocreales |
3.1.1. Magdeburger Börde (MBB)
3.1.2. Müncheberger Sander (MSA)
3.1.3. Uckermärkisches Hügelland (UKH)
3.2. Region-Specific Biomarkers and Their Microbiomes
3.3. Multifactorial Processes in Community Assembly
3.4. Closing Remarks
4. Materials and Methods
4.1. Study Sites and Sample Collection

4.2. Landscape Characteristics
4.2.1. Soil and Yield Potential
4.2.2. Climate
4.3. Sample Collection
4.4. Fungi Isolation and Determination
4.5. Fungal Quantification with qPCR
4.6. NGS
4.7. Data Management
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Appendix A


| Field | Isolated Colonies | Total | |
|---|---|---|---|
| Alternaria | Fusarium | ||
| MBB1 | 19 | 18 | 37 |
| MBB2 | 14 | 18 | 32 |
| MBB3 | 33 | 12 | 45 |
| MSA1 | 18 | 16 | 34 |
| MSA2 | 17 | 13 | 30 |
| UKH1 | 22 | 15 | 37 |
| UKH2 | 15 | 13 | 28 |
| UKH3 | 18 | 22 | 40 |
| UKH4 | 22 | 4 | 26 |
| UKH5 | 21 | 15 | 36 |
| total | 360 | 260 | 620 |
| Df | SumOfSqs | R2 | F | Pr (>F) | ||
|---|---|---|---|---|---|---|
| Soil landscape | 2 | 9.2999 | 0.43751 | 203.64 | 0.001 | *** |
| Field | 7 | 6.5676 | 0.30897 | 41.09 | 0.001 | *** |
| Residual | 236 | 5.3887 | 0.25351 | |||
| Total | 245 | 21.2562 | 1 |
| Df | SumOfSqs | R2 | F | Pr (>F) | ||
|---|---|---|---|---|---|---|
| MSA | ||||||
| Field | 2 | 1384 | 0.14592 | 6.5776 | 0.001 | *** |
| Residual | 77 | 8100.8 | 0.85408 | |||
| Total | 79 | 9484.8 | 1 | |||
| MBB | ||||||
| Field | 1 | 206.7 | 0.04366 | 1.3238 | 0.01 | ** |
| Residual | 29 | 4528.3 | 0.95634 | |||
| Total | 30 | 4735 | 1 | |||
| UKH | ||||||
| Field | 4 | 3437.3 | 0.16316 | 6.3368 | 0.001 | *** |
| Residual | 130 | 17,629.3 | 0.83684 | |||
| Total | 134 | 21,066.6 | 1 |
| Df | Variance | F | Pr (>F) | ||
|---|---|---|---|---|---|
| Model | 8 | 37.083 | 83.491 | 0.001 | *** |
| Residual | 236 | 131.025 | |||
| Terms | |||||
| TWI | 1 | 2.903 | 52.281 | 0.001 | *** |
| qPCR Allternaria | 1 | 10.266 | 184.912 | 0.001 | *** |
| qPCR Fusarium | 1 | 3.287 | 59.197 | 0.001 | *** |
| Plant height | 1 | 2.474 | 4.457 | 0.001 | *** |
| SQR.rating | 4 | 18.153 | 81.743 | 0.001 | *** |
| Residual | 236 | 131.025 | |||
| Correlation with axis | |||||
| CAP1 | CAP2 | ||||
| TWI | 0.25268324 | −0.2731339 | |||
| copiesALT | −0.67589029 | −0.2029782 | |||
| copiesFUS | −0.05474777 | 0.3695871 | |||
| Height | 0.31313261 | 0.5519229 |
| Variables | Correlation Method | Correlation Coefficient | p-Value | p-Adjusted | |
|---|---|---|---|---|---|
| Height | Pearson | 0.06262916 | 0.023 | 0.023 | * |
| TWI | Pearson | 0.13793507 | 0.001 | 0.001333 | ** |
| copiesALT | Pearson | 0.13893371 | 0.001 | 0.001333 | ** |
| copiesFUS | Pearson | 0.18767443 | 0.001 | 0.001333 | ** |
References
- Kremen, C.; Iles, A.; Bacon, C. Diversified Farming Systems: An Agroecological, Systems-based Alternative to Modern Industrial Agriculture. Ecol. Soc. 2012, 17, 44. [Google Scholar] [CrossRef]
- Beillouin, D.; Ben-Ari, T.; Makowski, D. Evidence map of crop diversification strategies at the global scale. Environ. Res. Lett. 2019, 14, 123001. [Google Scholar] [CrossRef]
- Tamburini, G.; Bommarco, R.; Wanger, T.C.; Kremen, C.; van der Heijden, M.G.A.; Liebman, M.; Hallin, S. Agricultural diversification promotes multiple ecosystem services without compromising yield. Sci. Adv. 2020, 6, eaba1715. [Google Scholar] [CrossRef]
- Hernández-Ochoa, I.M.; Gaiser, T.; Kersebaum, K.C.; Webber, H.; Seidel, S.J.; Grahmann, K.; Ewert, F. Model-based design of crop diversification through new field arrangements in spatially heterogeneous landscapes. A review. Agron. Sustain. Dev. 2022, 42, 74. [Google Scholar] [CrossRef]
- Schaller, J.; Frei, S.; Rohn, L.; Gilfedder, B.S. Amorphous Silica Controls Water Storage Capacity and Phosphorus Mobility in Soils. Front. Environ. Sci. 2020, 8, 94. [Google Scholar] [CrossRef]
- Pan, R.; Da Silva Martinez, A.; Santos Brito, T.; Pereira Seidel, E. Processes of Soil Infiltration and Water Retention and Strategies to Increase Their Capacity. J. Exp. Agric. Int. 2019, 9, 84–87. [Google Scholar] [CrossRef]
- Saruchi; Kumar, V.; Mittal, H.; Alhassan, S.M. Biodegradable hydrogels of tragacanth gum polysaccharide to improve water retention capacity of soil and environment-friendly controlled release of agrochemicals. Int. J. Biol. Macromol. 2019, 132, 1252–1261. [Google Scholar] [CrossRef] [PubMed]
- Olesen, J.E.; Trnka, M.; Kersebaum, K.C.; Skjelvåg, A.O.; Seguin, B.; Peltonen-Sainio, P.; Rossi, F.; Kozyra, J.; Micale, F. Impacts and adaptation of European crop production systems to climate change. Eur. J. Agron. 2011, 34, 96–112. [Google Scholar] [CrossRef]
- Gambella, F.; Bianchini, L.; Cecchini, M.; Egidi, G.; Ferrara, A.; Salvati, L.; Colantoni, A.; Morea, D. Moving toward the north? The spatial shift of olive groves in Italy. Agric. Econ. 2021, 67, 129–135. [Google Scholar] [CrossRef]
- King, M.; Altdorff, D.; Li, P.; Galagedara, L.; Holden, J.; Unc, A. Northward shift of the agricultural climate zone under 21st-century global climate change. Sci. Rep. 2018, 8, 7904. [Google Scholar] [CrossRef] [PubMed]
- Webber, H.; Ewert, F.; Olesen, J.E.; Müller, C.; Fronzek, S.; Ruane, A.C.; Bourgault, M.; Martre, P.; Ababaei, B.; Bindi, M.; et al. Diverging importance of drought stress for maize and winter wheat in Europe. Nat. Commun. 2018, 9, 4249. [Google Scholar] [CrossRef] [Green Version]
- Le Gouis, J.; Oury, F.X.; Charmet, G. How changes in climate and agricultural practices influenced wheat production in Western Europe. J. Cereal Sci. 2020, 93, 102960. [Google Scholar] [CrossRef]
- Parikka, P.; Hakala, K.; Tiilikkala, K. Expected shifts in Fusarium species’ composition on cereal grain in Northern Europe due to climatic change. Food Addit. Contam.—Part A 2012, 29, 1543–1555. [Google Scholar] [CrossRef]
- Desai, S.; Dubey, S.C.; Prasad, R.D. Impacts of climate change on Fusarium species vis-à-vis adaptation strategies. Indian Phytopathol. 2020, 73, 593–603. [Google Scholar] [CrossRef]
- Lawrence, C.B.; Mitchell, T.K.; Craven, K.D.; Cho, Y.; Cramer, R.A.; Kim, K.H. At death’s door: Alternaria pathogenicity mechanisms. Plant Pathol. J. 2008, 24, 101–111. [Google Scholar] [CrossRef]
- Zwickel, T.; Kahl, S.; Klaffke, H.; Rychlik, M.; Müller, M. Spotlight on the Underdogs—An Analysis of Underrepresented Alternaria Mycotoxins Formed Depending on Varying Substrate, Time and Temperature Conditions. Toxins 2016, 8, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomma, B.P.H.J. Alternaria spp.: From general saprophyte to specific parasite. Mol. Plant Pathol. 2003, 4, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Schiro, G.; Verch, G.; Grimm, V.; Müller, M. Alternaria and Fusarium Fungi: Differences in Distribution and Spore Deposition in a Topographically Heterogeneous Wheat Field. J. Fungi 2018, 4, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, F.; Cazzato, S.; Walder, F.; Vogelgsang, S.; Bender, S.F.; van der Heijden, M.G.A. Humidity and high temperature are important for predicting fungal disease outbreaks worldwide. New Phytol. 2022, 234, 1553–1556. [Google Scholar] [CrossRef] [PubMed]
- Kivlin, S.N.; Winston, G.C.; Goulden, M.L.; Treseder, K.K. Environmental filtering affects soil fungal community composition more than dispersal limitation at regional scales. Fungal Ecol. 2014, 12, 14–25. [Google Scholar] [CrossRef]
- Kelly, A.C.; Clear, R.M.; O’Donnell, K.; McCormick, S.; Turkington, T.K.; Tekauz, A.; Gilbert, J.; Kistler, H.C.; Busman, M.; Ward, T.J. Diversity of Fusarium head blight populations and trichothecene toxin types reveals regional differences in pathogen composition and temporal dynamics. Fungal Genet. Biol. 2015, 82, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Pasquali, M.; Beyer, M.; Logrieco, A.; Audenaert, K.; Balmas, V.; Basler, R.; Boutigny, A.L.; Chrpová, J.; Czembor, E.; Gagkaeva, T.; et al. A European Database of Fusarium graminearum and F. culmorum Trichothecene Genotypes. Front. Microbiol. 2016, 7, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goertz, A.; Zuehlke, S.; Spiteller, M.; Steiner, U.; Dehne, H.W.; Waalwijk, C.; de Vries, I.; Oerke, E.C. Fusarium species and mycotoxin profiles on commercial maize hybrids in Germany. Eur. J. Plant Pathol. 2010, 128, 101–111. [Google Scholar] [CrossRef]
- Müller, M.E.H.; Brenning, A.; Verch, G.; Koszinski, S.; Sommer, M. Multifactorial spatial analysis of mycotoxin contamination of winter wheat at the field and landscape scale. Agric. Ecosyst. Environ. 2010, 139, 245–254. [Google Scholar] [CrossRef]
- Shakir, S.; Zaidi, S.S.e.A.; de Vries, F.T.; Mansoor, S. Plant Genetic Networks Shaping Phyllosphere Microbial Community. Trends Genet. 2021, 37, 306–316. [Google Scholar] [CrossRef]
- Bashir, I.; War, A.F.; Rafiq, I.; Reshi, Z.A.; Rashid, I.; Shouche, Y.S. Phyllosphere microbiome: Diversity and functions. Microbiol. Res. 2022, 254, 126888. [Google Scholar] [CrossRef]
- Schiro, G.; Colangeli, P.; Müller, M.E.H. A Metabarcoding Analysis of the Mycobiome of Wheat Ears Across a Topographically Heterogeneous Field. Front. Microbiol. 2019, 10, 2095. [Google Scholar] [CrossRef] [Green Version]
- Sapkota, R.; Jørgensen, L.N.; Nicolaisen, M. Spatiotemporal variation and networks in the mycobiome of the wheat canopy. Front. Plant Sci. 2017, 8, 1357. [Google Scholar] [CrossRef] [Green Version]
- Munkvold, G.P. Fusarium Species and Their Associated Mycotoxins. Methods Mol. Biol. 2017, 1542, 51–106. [Google Scholar] [CrossRef]
- Mirocha, C.J.; Xie, W.; Xu, Y.; Wilcoxson, R.D.; Woodward, R.P.; Etebarian, R.H.; Behele, G. Production of trichothecene mycotoxins by Fusarium graminearum and Fusarium culmorum on barley and wheat. Mycopathologia 1994, 128, 19–23. [Google Scholar] [CrossRef]
- Fahrig, L.; Girard, J.; Duro, D.; Pasher, J.; Smith, A.; Javorek, S.; King, D.; Lindsay, K.F.; Mitchell, S.; Tischendorf, L. Farmlands with smaller crop fields have higher within-field biodiversity. Agric. Ecosyst. Environ. 2015, 200, 219–234. [Google Scholar] [CrossRef]
- Šálek, M.; Hula, V.; Kipson, M.; Daňková, R.; Niedobová, J.; Gamero, A. Bringing diversity back to agriculture: Smaller fields and non-crop elements enhance biodiversity in intensively managed arable farmlands. Ecol. Indic. 2018, 90, 65–73. [Google Scholar] [CrossRef]
- Martiny, J.B.H.; Bohannan, B.J.M.; Brown, J.H.; Colwell, R.K.; Fuhrman, J.A.; Green, J.L.; Horner-Devine, M.C.; Kane, M.; Krumins, J.A.; Kuske, C.R.; et al. Microbial biogeography: Putting microorganisms on the map. Nat. Rev. Microbiol. 2006, 4, 102–112. [Google Scholar] [CrossRef]
- Hanson, C.A.; Fuhrman, J.A.; Horner-Devine, M.C.; Martiny, J.B.H. Beyond biogeographic patterns: Processes shaping the microbial landscape. Nat. Rev. Microbiol. 2012, 10, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Schiro, G.; Müller, T.; Verch, G.; Sommerfeld, T.; Mauch, T.; Koch, M.; Grimm, V.; Müller, M.E.H. The distribution of mycotoxins in a heterogeneous wheat field in relation to microclimate, fungal and bacterial abundance. J. Appl. Microbiol. 2019, 126, 177–190. [Google Scholar] [CrossRef]
- Kokkonen, M.; Jestoi, M.; Laitila, A. Mycotoxin production of Fusarium langsethiae and Fusarium sporotrichioides on cereal-based substrates. Mycotoxin Res. 2012, 28, 25–35. [Google Scholar] [CrossRef]
- Strayer, D.L.; Power, M.E.; Fagan, W.F.; Pickett, S.T.A.; Belnap, J. A classification of ecological boundaries. Bioscience 2003, 53, 723–729. [Google Scholar] [CrossRef] [Green Version]
- Cadenasso, M.L.; Pickett, S.T.A.; Weathers, K.C.; Jones, C.G. A framework for a theory of ecological boundaries. Bioscience 2003, 53, 750–758. [Google Scholar] [CrossRef] [Green Version]
- Marburger, D.A.; Venkateshwaran, M.; Conley, S.P.; Esker, P.D.; Lauer, J.G.; Ané, J.M. Crop Rotation and Management Effect on Fusarium spp. Populations. Crop Sci. 2015, 55, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Marburger, D.A.; Conley, S.P.; Esker, P.D.; Lauer, J.G.; Ané, J.-M.; Marburger, D.A.; Conley, S.P.; Lauer, J.G.; Ané, J.-M. Yield Response to Crop/Genotype Rotations and Fungicide Use to Manage Fusarium-related Diseases. Crop Sci. 2015, 55, 889–898. [Google Scholar] [CrossRef]
- Drakopoulos, D.; Sulyok, M.; Jenny, E.; Kägi, A.; Bänziger, I.; Logrieco, A.F.; Krska, R.; Vogelgsang, S.; Jedryczka, M. Fusarium Head Blight and Associated Mycotoxins in Grains and Straw of Barley: Influence of Agricultural Practices. Agronomy 2021, 11, 801. [Google Scholar] [CrossRef]
- Stenglein, S.A. Fusarium poae: A pathogen that needs more attention. J. Plant Pathol. 2009, 91, 25–36. [Google Scholar]
- Vogelgsang, S.; Sulyok, M.; Hecker, A.; Jenny, E.; Krska, R.; Schuhmacher, R.; Forrer, H.R. Toxigenicity and pathogenicity of Fusarium poae and Fusarium avenaceum on wheat. Eur. J. Plant Pathol. 2008, 122, 265–276. [Google Scholar] [CrossRef]
- Tillmann, M.; von Tiedemann, A.; Winter, M. Crop rotation effects on incidence and diversity of Fusarium species colonizing stem bases and grains of winter wheat. J. Plant Dis. Prot. 2017, 124, 121–130. [Google Scholar] [CrossRef]
- Urbina, H.; Aime, M.C. A closer look at Sporidiobolales: Ubiquitous microbial community members of plant and food biospheres View supplementary material A closer look at Sporidiobolales: Ubiquitous microbial community members of plant and food biospheres. Mycologia 2018, 110, 79–92. [Google Scholar] [CrossRef]
- Schisler, D.A.; Slininger, P.J.; Boehm, M.J.; Paul, P.A. Co-culture of Yeast Antagonists of Fusarium Head Blight and their Effect on Disease Development in Wheat. Plant Pathol. J. 2011, 10, 128–137. [Google Scholar] [CrossRef] [Green Version]
- Schisler, D.A.; Core, A.B.; Boehm, M.J.; Horst, L.; Krause, C.; Dunlap, C.A.; Rooney, A.P. Population dynamics of the Fusarium head blight biocontrol agent Cryptococcus flavescens OH 182.9 on wheat anthers and heads. Biol. Control 2014, 70, 17–27. [Google Scholar] [CrossRef]
- Podgórska-Kryszczuk, I.; Solarska, E.; Kordowska-Wiater, M. Biological Control of Fusarium culmorum, Fusarium graminearum and Fusarium poae by Antagonistic Yeasts. Pathogens 2022, 11, 86. [Google Scholar] [CrossRef]
- Rojas, E.C.; Sapkota, R.; Jensen, B.; Jørgensen, H.J.L.; Henriksson, T.; Jørgensen, L.N.; Nicolaisen, M.; Collinge, D.B. Fusarium Head Blight Modifies Fungal Endophytic Communities During Infection of Wheat Spikes. Microb. Ecol. 2020, 79, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, I.; Persson, P.; Friberg, H. Fusarium Head Blight From a Microbiome Perspective. Front. Microbiol. 2021, 12, 371. [Google Scholar] [CrossRef]
- Fakhrunnisa; Hashmi, M.H.; Ghaffar, A. In vitro interaction of Fusarium spp., with other fungi. Pak. J. Bot. 2006, 38, 1317–1322. [Google Scholar]
- Hoffmann, A.; Lischeid, G.; Koch, M.; Lentzsch, P.; Sommerfeld, T.; Müller, M.E.H. Co-cultivation of Fusarium, Alternaria, and Pseudomonas on wheat-ears affects microbial growth and mycotoxin production. Microorganisms 2021, 9, 443. [Google Scholar] [CrossRef] [PubMed]
- Matarese, F.; Sarrocco, S.; Gruber, S.; Seidl-Seiboth, V.; Vannacci, G. Biocontrol of Fusarium head blight: Interactions between Trichoderma and mycotoxigenic Fusarium. Microbiology 2012, 158, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J. Trichoderma as biocontrol agent against pests: New uses for a mycoparasite. Biol. Control 2021, 159, 104634. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Adler, P.B.; Godoy, O.; James, E.C.; Fuller, S.; Levine, J.M. Community assembly, coexistence and the environmental filtering metaphor. Funct. Ecol. 2015, 29, 592–599. [Google Scholar] [CrossRef]
- Schlägel, U.E.; Grimm, V.; Blaum, N.; Colangeli, P.; Dammhahn, M.; Eccard, J.A.; Hausmann, S.L.; Herde, A.; Hofer, H.; Joshi, J.; et al. Movement-mediated community assembly and coexistence. Biol. Rev. 2020, 95, 1073–1096. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, A.; Funk, R.; Müller, M.E.H. Blowin’ in the Wind: Wind Dispersal Ability of Phytopathogenic Fusarium in a Wind Tunnel Experiment. Atmosphere 2021, 12, 1653. [Google Scholar] [CrossRef]
- Thapa, S.; Prasanna, R. Prospecting the characteristics and significance of the phyllosphere microbiome. Ann. Microbiol. 2018, 68, 229–245. [Google Scholar] [CrossRef]
- Xu, N.; Zhao, Q.; Zhang, Z.; Zhang, Q.; Wang, Y.; Qin, G.; Ke, M.; Qiu, D.; Peijnenburg, W.J.G.M.; Lu, T.; et al. Phyllosphere Microorganisms: Sources, Drivers, and Their Interactions with Plant Hosts. J. Agric. Food Chem. 2022, 70, 4860–4870. [Google Scholar] [CrossRef]
- Trivellone, V.; Hoberg, E.P.; Boeger, W.A.; Brooks, D.R. Food security and emerging infectious disease: Risk assessment and risk management. R. Soc. Open Sci. 2022, 9, 211687. [Google Scholar] [CrossRef]
- Kopecký, M.; Macek, M.; Wild, J. Topographic Wetness Index calculation guidelines based on measured soil moisture and plant species composition. Sci. Total Environ. 2021, 757, 143785. [Google Scholar] [CrossRef]
- Beven, K.J.; Kirkby, M.J.; Kirkby, A.J. A physically based, variable contributing area model of basin hydrology. Hydrol. Sci. J. 1979, 24, 43–69. [Google Scholar] [CrossRef]
- Nierenberg, H. Untersuchungen über die morphologische und biologische Differenzierung in der Fusarium-Sektion Liseola. In Mitteilungen aus der Biologischen Bundesanstalt für Land- und Forstwirtschaft Berlin-Dahlem—169; Kommissionsverlag Paul Parey: Berlin, Germany, 1976; ISBN 3-489-16900-X. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Hoboken, NJ, USA, 2007; pp. 1–388. [Google Scholar] [CrossRef]
- Noman, E.; Al-Gheethi, A.; Rahman, N.K.; Talip, B.; Mohamed, R.; Kadir, O.A. Single Spore Isolation as a Simple and Efficient Technique to obtain fungal pure culture. IOP Conf. Ser. Earth Environ. Sci. 2018, 140, 012055. [Google Scholar] [CrossRef]
- Geiser, D.M.; del Mar Jiménez-Gasco, M.; Kang, S.; Makalowska, I.; Veeraraghavan, N.; Ward, T.J.; Zhang, N.; Kuldau, G.A.; O’donnell, K. FUSARIUM-ID v. 1.0: A DNA Sequence Database for Identifying Fusarium. Eur. J. Plant Pathol. 2004, 110, 473–479. [Google Scholar] [CrossRef]
- Müller, T.; Ruppel, S.; Behrendt, U.; Lentzsch, P.; Müller, M.E.H. Antagonistic potential of fluorescent pseudomonads colonizing wheat heads against mycotoxin producing alternaria and fusaria. Front. Microbiol. 2018, 9, 2124. [Google Scholar] [CrossRef]
- Gerling, M.; Pätzig, M.; Hempel, L.; Büttner, C.; Müller, M.E.H. Arable Weeds at the Edges of Kettle Holes as Overwintering Habitat for Phytopathogenic Fungi. Agronomy 2022, 12, 823. [Google Scholar] [CrossRef]
- Li, S.; Deng, Y.; Wang, Z.; Zhang, Z.; Kong, X.; Zhou, W.; Yi, Y.; Qu, Y. Exploring the accuracy of amplicon-based internal transcribed spacer markers for a fungal community. Mol. Ecol. Resour. 2020, 20, 170–184. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. vegan: Community Ecology Package 2022.
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Liu, C.; Cui, Y.; Li, X.; Yao, M. microeco: An R package for data mining in microbial community ecology. FEMS Microbiol. Ecol. 2021, 97, 255. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]












| Field | MBB1 | MBB2 | MBB3 | MSA1 | MSA2 | UKH1 | UKH2 | UKH3 | UKH4 | UKH5 |
|---|---|---|---|---|---|---|---|---|---|---|
| MBB1 | 0 | |||||||||
| MBB2 | 1.4 | 0 | ||||||||
| MBB3 | 2.6 | 0.2 | 0 | |||||||
| MSA1 | 179.7 | 182.2 | 183.2 | 0 | ||||||
| MSA2 | 179.7 | 182.2 | 183.3 | 0.02 | 0 | |||||
| UKH1 | 196.3 | 198.4 | 199.8 | 91.3 | 91.3 | 0 | ||||
| UKH2 | 196.3 | 198.4 | 199.9 | 92.0 | 92.0 | 0 | 0 | |||
| UKH3 | 193.6 | 195.7 | 197.1 | 92.1 | 92.1 | 2.4 | 21.8 | 0 | ||
| UKH4 | 187.0 | 189.0 | 190.5 | 95.2 | 95.2 | 11.0 | 10.7 | 7.8 | 0 | |
| UKH5 | 188.4 | 190.4 | 191.8 | 98.7 | 98.7 | 12.8 | 12.4 | 9.7 | 2.5 | 0 |
| MBB | January | February | March | April | May | June | July |
|---|---|---|---|---|---|---|---|
| Ø air temperature (°C) | 4.5 | 6.2 | 5.8 | 10.7 | 12 | 18.4 | 18.1 |
| Ø rel. humidity (%) | 83.1 | 76.2 | 66.5 | 53.4 | 61.7 | 65.2 | 64.9 |
| precipitation sum (mm) | 23 | 81 | 27 | 7 | 27 | 72 | 40 |
| MSA | |||||||
| Ø air temperature (°C) | 4.0 | 5.7 | 4.9 | 9.9 | 12.6 | 18.5 | 18.1 |
| Ø rel. humidity (%) | 84.0 | 78.1 | 69.3 | 55.9 | - | 67.9 | 68.2 |
| precipitation sum (mm) | 42 | 77 | 24 | 17 | 29 | 59 | 43 |
| UKH1–UKH3 | |||||||
| Ø air temperature (°C) | 4.1 | 5.3 | 4.5 | 8.9 | 12.6 | 17.4 | 17.4 |
| Ø rel. humidity (%) | 89.2 | 81.2 | 74.8 | 63.3 | 70.5 | 72.0 | 68.7 |
| precipitation sum (mm) | 36 | 53 | 36 | 14 | 27 | 37 | 30 |
| UKH4–UKH5 | |||||||
| Ø air temperature (°C) | 4.0 | 5.2 | 4.4 | 8.9 | 12.4 | 17.3 | 17.0 |
| Ø rel. humidity (%) | 89.2 | 81.2 | 74.8 | 63.3 | 70.5 | 72.0 | 68.7 |
| precipitation sum (mm) | 40 | 56 | 38 | 13 | 26 | 52 | 37 |
| Region | Long-Term Average Temperature (°C) | Long-Term Average Precipitation (mm) |
|---|---|---|
| MBB | 9.9 | 534 |
| MSA | 9.5 | 553 |
| UKH1–UKH3 | 9.1 | 553 |
| UKH4–UKH5 | 9.0 | 580 |
| S SUF pl3 | Probe | 5′-ACCCTTACCGAGCTCAGCGGCTTCCTATT-3′ |
|---|---|---|
| Fa PL3 f | forward | 5′-TACCCCGCCACTCGAGCG-3′ |
| Fus pl rev | reverse | 5′-TTGAGCTTGTCAAGAACCCAGGCG-3′ |
| Alt-Prt | probe | 5′-TGGGTTCGCCCACCACTAGGACA-3′ |
| Alt-F | forward | 5′-TCTTTTGCGTACTTCTTGTTTCCTT-3′ |
| Alt-R | reverse | 5′-TTACTGACGCTGATTGCAATTACA-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoffmann, A.; Posirca, A.-R.; Lewin, S.; Verch, G.; Büttner, C.; Müller, M.E.H. Environmental Filtering Drives Fungal Phyllosphere Community in Regional Agricultural Landscapes. Plants 2023, 12, 507. https://doi.org/10.3390/plants12030507
Hoffmann A, Posirca A-R, Lewin S, Verch G, Büttner C, Müller MEH. Environmental Filtering Drives Fungal Phyllosphere Community in Regional Agricultural Landscapes. Plants. 2023; 12(3):507. https://doi.org/10.3390/plants12030507
Chicago/Turabian StyleHoffmann, Annika, Alexandra-Raluca Posirca, Simon Lewin, Gernot Verch, Carmen Büttner, and Marina E. H. Müller. 2023. "Environmental Filtering Drives Fungal Phyllosphere Community in Regional Agricultural Landscapes" Plants 12, no. 3: 507. https://doi.org/10.3390/plants12030507
APA StyleHoffmann, A., Posirca, A.-R., Lewin, S., Verch, G., Büttner, C., & Müller, M. E. H. (2023). Environmental Filtering Drives Fungal Phyllosphere Community in Regional Agricultural Landscapes. Plants, 12(3), 507. https://doi.org/10.3390/plants12030507