

Wheat Stem Rust Detection and Race Characterization in Tunisia
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Stem Rust Disease Occurence in Tunisia
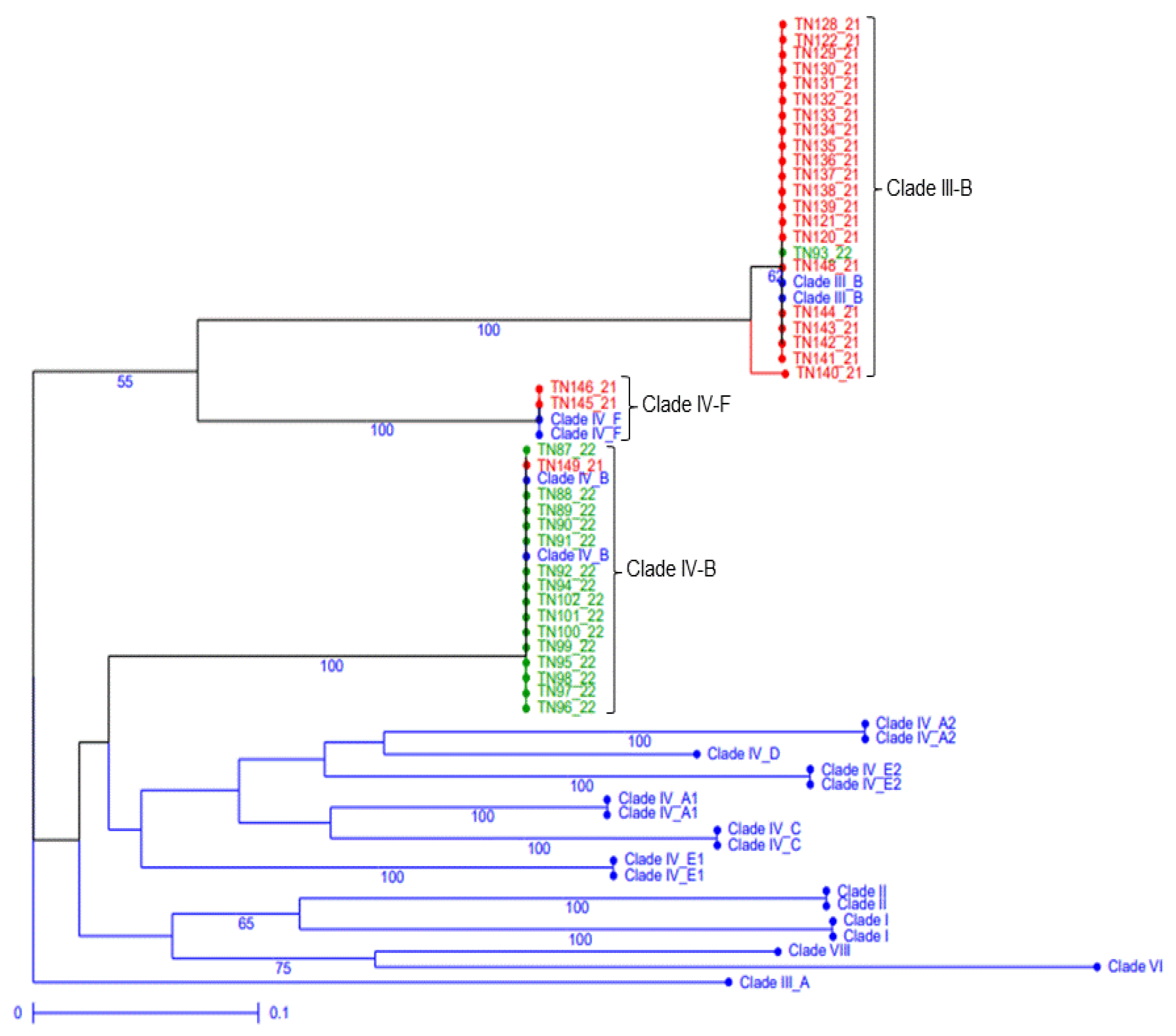
2.2. Genotyping and Race Phenotyping
2.3. Climatic Factors Effect on Pgt Occurrence in Tunisia
3. Discussion
3.1. Pgt Occurrence in the Tunisian Environment
3.2. Race Dominance in Tunisia and Possible Infection Source
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Disease Assessment and Sampling
4.3. Genotyping and Race Phenotyping
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fones, H.N.; Bebber, D.P.; Chaloner, T.M.; Kay, W.T.; Steinberg, G.; Gurr, S.J. Threats to Global Food Security from Emerging Fungal and Oomycete Crop Pathogens. Nat. Food 2020, 1, 332–342. [Google Scholar] [CrossRef]
- Chaloner, T.M.; Gurr, S.J.; Bebber, D.P. Plant Pathogen Infection Risk Tracks Global Crop Yields under Climate Change. Nat. Clim. Chang. 2021, 11, 710–715. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The Global Burden of Pathogens and Pests on Major Food Crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, M.; Alderman, S.; Garvin, D.F.; Pfender, W.F. Infection of Brachypodium Distachyon by Formae Speciales of Puccinia graminis: Early Infection Events and Host-Pathogen Incompatibility. PLoS ONE 2013, 8, e56857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savile: Taxonomy of the Cereal Rust Fungi-Google Scholar. Available online: https://scholar.google.com/scholar_lookup?title=Taxonomy%20of%20the%20cereal%20rust%20fungi&publication_year=1984&author=Savile%2CDBO (accessed on 15 September 2022).
- Olivera, P.; Newcomb, M.; Szabo, L.J.; Rouse, M.; Johnson, J.; Gale, S.; Luster, D.G.; Hodson, D.; Cox, J.A.; Burgin, L.; et al. Phenotypic and Genotypic Characterization of Race TKTTF of Puccinia graminis f. sp. tritici That Caused a Wheat Stem Rust Epidemic in Southern Ethiopia in 2013–14. Phytopathology 2015, 105, 917–928. [Google Scholar] [CrossRef] [Green Version]
- Meyer, M.; Cox, J.A.; Hitchings, M.D.T.; Burgin, L.; Hort, M.C.; Hodson, D.P.; Gilligan, C.A. Quantifying Airborne Dispersal Routes of Pathogens over Continents to Safeguard Global Wheat Supply. Nat. Plants 2017, 3, 780–786. [Google Scholar] [CrossRef]
- Lidwell-Durnin, J.; Lapthorn, A. The Threat to Global Food Security from Wheat Rust: Ethical and Historical Issues in Fighting Crop Diseases and Preserving Genetic Diversity. Glob. Food Secur. 2020, 26, 100446. [Google Scholar] [CrossRef]
- Golan, J.J.; Pringle, A. Long-Distance Dispersal of Fungi. Microbiol. Spectr. 2017, 24, 309–333. [Google Scholar]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology: Top 10 Fungal Pathogens. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Meyer, M.; Burgin, L.; Hort, M.C.; Hodson, D.P.; Gilligan, C.A. Large-Scale Atmospheric Dispersal Simulations Identify Likely Airborne Incursion Routes of Wheat Stem Rust Into Ethiopia. Phytopathology 2017, 107, 1175–1186. [Google Scholar] [CrossRef] [Green Version]
- Prank, M.; Kenaley, S.C.; Bergstrom, G.C.; Acevedo, M.; Mahowald, N.M. Climate Change Impacts the Spread Potential of Wheat Stem Rust, a Significant Crop Disease. Environ. Res. Lett. 2019, 14, 124053. [Google Scholar] [CrossRef]
- Lewis, C.M.; Persoons, A.; Bebber, D.P.; Kigathi, R.N.; Maintz, J.; Findlay, K.; Bueno-Sancho, V.; Corredor-Moreno, P.; Harrington, S.A.; Kangara, N.; et al. Potential for Re-Emergence of Wheat Stem Rust in the United Kingdom. Commun. Biol. 2018, 1, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodson, D.P. Shifting Boundaries: Challenges for Rust Monitoring. Euphytica 2011, 179, 93–104. [Google Scholar] [CrossRef]
- Singh, R.P.; Hodson, D.P.; Jin, Y.; Lagudah, E.S.; Ayliffe, M.A.; Bhavani, S.; Rouse, M.N.; Pretorius, Z.A.; Szabo, L.J.; Huerta-Espino, J.; et al. Emergence and Spread of New Races of Wheat Stem Rust Fungus: Continued Threat to Food Security and Prospects of Genetic Control. Phytopathology 2015, 105, 872–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, S. AGRICULTURE Wheat Rust Back in Europe. Nature 2017, 542, 145–146. [Google Scholar] [CrossRef]
- Olivera Firpo, P.D.; Newcomb, M.; Flath, K.; Sommerfeldt-Impe, N.; Szabo, L.J.; Carter, M.; Luster, D.G.; Jin, Y. Characterization of Puccinia graminis f. sp. tritici Isolates Derived from an Unusual Wheat Stem Rust Outbreak in Germany in 2013. Plant Pathol. 2017, 66, 1258–1266. [Google Scholar] [CrossRef]
- Patpour, M.; Hovmøller, M.S.; Rodriguez-Algaba, J.; Randazzo, B.; Villegas, D.; Shamanin, V.P.; Berlin, A.; Flath, K.; Czembor, P.; Hanzalova, A.; et al. Wheat Stem Rust Back in Europe: Diversity, Prevalence and Impact on Host Resistance. Front. Plant Sci. 2022, 13, 882440. [Google Scholar] [CrossRef]
- Fetch, T.; Mitchell Fetch, J.; Zegeye, T.; Xue, A. Races of Puccinia graminis on Barley, Oat, and Wheat in Canada from 2015 to 2019. Can. J. Plant Pathol. 2021, 43, 463–471. [Google Scholar] [CrossRef]
- Pretorius, Z.A.; Singh, R.P.; Wagoire, W.W.; Payne, T.S. Detection of Virulence to Wheat Stem Rust Resistance Gene Sr31 in Puccinia graminis f. sp. tritici in Uganda. Plant Dis. 2000, 84, 203. [Google Scholar] [CrossRef]
- Singh, R.P.; Hodson, D.P.; Huerta-Espino, J.; Jin, Y.; Bhavani, S.; Njau, P.; Herrera-Foessel, S.; Singh, P.K.; Singh, S.; Govindan, V. The Emergence of Ug99 Races of the Stem Rust Fungus Is a Threat to World Wheat Production. Annu. Rev. Phytopathol. 2011, 49, 465–481. [Google Scholar] [CrossRef] [Green Version]
- Rust Tracker.org: A Global wheat Rust Monitoring System. Pathotype Tracker-Where Is Ug99? Available online: https://rusttracker.cimmyt.org/?page_id=22 (accessed on 12 June 2022).
- Gill, B.K.; Klindworth, D.L.; Rouse, M.N.; Zhang, J.; Zhang, Q.; Sharma, J.S.; Chu, C.; Long, Y.; Chao, S.; Olivera, P.D.; et al. Function and Evolution of Allelic Variations of Sr13 Conferring Resistance to Stem Rust in Tetraploid Wheat (Triticum Turgidum L.). Plant J. 2021, 106, 1674–1691. [Google Scholar] [CrossRef]
- Olivera, P.D.; Bulbula, W.D.; Badebo, A.; Bockelman, H.E.; Edae, E.A.; Jin, Y. Field Resistance to Wheat Stem Rust in Durum Wheat Accessions Deposited at the USDA National Small Grains Collection. Crop. Sci. 2021, 61, 2565–2578. [Google Scholar] [CrossRef]
- Megerssa, S.H.; Ammar, K.; Acevedo, M.; Brown-Guedira, G.; Ward, B.; Degete, A.G.; Randhawa, M.S.; Sorrells, M.E. Multiple-Race Stem Rust Resistance Loci Identified in Durum Wheat Using Genome-Wide Association Mapping. Front. Plant Sci. 2020, 11, 598509. [Google Scholar] [CrossRef]
- Kumar, S.; Fetch, T.G.; Knox, R.E.; Singh, A.K.; Clarke, J.M.; Depauw, R.M.; Cuthbert, R.D.; Campbell, H.L.; Singh, D.; Bhavani, S.; et al. Mapping of Ug99 Stem Rust Resistance in Canadian Durum Wheat. Can. J. Plant Pathol. 2021, 43, 599–611. [Google Scholar] [CrossRef]
- Romdhani, M. La Rouille Noire Du Blé: Résistance Variétale, Races Physiologiques et Dégâts. Ph.D. Thesis, National Agronomic Institute in Tunisia, Tunis, Tunisia, 1976. [Google Scholar]
- Yahyaoui, A. Professor at Borlaug Training Foundation at Colorado State University, Colorado state, United States, CRP Wheat Septoria Precision Phenotyping Platform (CIMMYT/IRESA). Personal communication. 2019. [Google Scholar]
- Hovmøller, M.S.; Patpour, M.; Rodriguez-Algaba, J.; Justesen, A.F.; Hansen, J.G. GRRC Annual Report 2019: Stem- and Yellow Rust Genotyping and Race Analyses. 10. Available online: https://agro.au.dk/forskning/internationale-platforme/wheatrust (accessed on 1 September 2022).
- Patpour, M.; Hovmøller, M.S.; Hansen, J.G.; Justesen, A.F.; Thach, T.; Rodriguez-Algab, J.; Hodson, D.; Randazzo, B. Epidemics of Yellow and Stem Rust in Southern Italy 2016–2017 | GlobalRust.Org. Available online: https://globalrust.org/content/epidemics-yellow-and-stem-rust-southern-Italy-2016-2017 (accessed on 3 September 2022).
- Nazari, K.; Kurtulus, E.; Kavaz, H.; Ozturk, O.M.; Egerci, Y.; Cer, C.; Jarrahi, T.; Gasmi, C.; Kadiroglu, A. First Report of Races TKTTP and TKKTP of Puccinia graminis f. sp. tritici with Virulence to Wheat Stem Rust Resistance Gene Sr24 in Turkey and Tunisia. Plant Dis. 2022, 106, 757. [Google Scholar] [CrossRef]
- Park, R.; Fetch, T.; Hodson, D.; Jin, Y.; Nazari, K.; Prashar, M.; Pretorius, Z. International Surveillance of Wheat Rust Pathogens: Progress and Challenges. Euphytica 2011, 179, 109–117. [Google Scholar] [CrossRef]
- Chakraborty, S.; Luck, J.; Hollaway, G.; Fitzgerald, G.; White, N. Rust-Proofing Wheat for a Changing Climate. Euphytica 2011, 179, 19–32. [Google Scholar] [CrossRef]
- Ali, S.; Gladieux, P.; Leconte, M.; Gautier, A.; Justesen, A.F.; Hovmøller, M.S.; Enjalbert, J.; de Vallavieille-Pope, C. Origin, Migration Routes and Worldwide Population Genetic Structure of the Wheat Yellow Rust Pathogen Puccinia striiformis f.sp. tritici. PLoS Pathog. 2014, 10, e1003903. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.K.M.; Hovmøller, M.S. Aerial Dispersal of Pathogens on the Global and Continental Scales and Its Impact on Plant Disease. Science 2002, 297, 537–541. [Google Scholar] [CrossRef] [Green Version]
- Baiocco, S.; Cavina, F.; Pradolesi, G. A Weather-Based Simulation Model for the Development of Wheat Stem Rust Epidemics. In Proceedings of the 2021 29th Conference of Open Innovations Association (FRUCT), Tampere, Finland, 12–14 May 2021; IEEE: Piscataway, NJ, USA, 2021; pp. 22–29. Available online: https://ieeexplore.ieee.org/document/9435559 (accessed on 10 November 2022). [CrossRef]
- Ben M’Barek, S.; Karisto, P.; Abdedayem, W.; Laribi, M.; Fakhfakh, M.; Kouki, H.; Mikaberidze, A.; Yahyaoui, A. Improved Control of Septoria Tritici Blotch in Durum Wheat Using Cultivar Mixtures. Plant Pathol. 2020, 69, 1655–1665. [Google Scholar] [CrossRef]
- Pfender, W.F. Effect of Autumn Planting Date and Stand Age on Severity of Stem Rust in Seed Crops of Perennial Ryegrass. Plant Dis. 2004, 88, 1017–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naseri, B.; Sabeti, P. Analysis of the Effects of Climate, Host Resistance, Maturity and Sowing Date on Wheat Stem Rust Epidemics. J. Plant Pathol. 2021, 103, 197–205. [Google Scholar] [CrossRef]
- Southern, J.W.; Wilcoxson, R.D. Effect of Planting Date on Slow Rusting of Wheat by Puccinia graminis f. sp. tritici. Int. J. Trop. Plant Dis. 1983, 1, 21–24. [Google Scholar]
- Stubbs, R.W.; Prescott, J.M.; Saari, E.E.; Dubin, H.J. Cereal Disease Methodology Manual; CIMMYT: Mexico City, Mexico, 1986; 46p. [Google Scholar]
- Roelfs, A.P.; Martens, J.W. An International System of Nomenclature for Puccinia graminis f. sp. tritici. Phytopathology 1988, 78, 526–533. [Google Scholar] [CrossRef] [Green Version]
- Olivera, P.D.; Sikharulidze, Z.; Dumbadze, R.; Szabo, L.J.; Newcomb, M.; Natsarishvili, K.; Rouse, M.N.; Luster, D.G.; Jin, Y. Presence of a Sexual Population of Puccinia graminis f. sp. tritici in Georgia Provides a Hotspot for Genotypic and Phenotypic Diversity. Phytopathology 2019, 109, 2152–2160. [Google Scholar] [CrossRef] [Green Version]
- WheatRust. Available online: https://agro.au.dk/forskning/internationale-platforme/wheatrust (accessed on 26 October 2022).
- Patpour, M.; Justesen, A.F.; Tecle, A.W.; Yazdani, M.; Yasaie, M.; Hovmøller, M.S. First Report of Race TTRTF of Wheat Stem Rust (Puccinia graminis f. sp. tritici) in Eritrea. Plant Dis. 2020, 104, 973. [Google Scholar] [CrossRef]
- Esmail, M.; Szabo, L.J. Wheat Stem Rust Pathogen (Pgt) Identification and Characterization in Egypt Using Single Nucleotide Polymorphism (SNP) Markers | GlobalRust.Org. Available online: https://globalrust.org/content/wheat-stem-rust-pathogen-pgt-identification-and-characterization-egypt-using-single (accessed on 3 September 2022).
- Tesfaye, T.; Chala, A.; Shikur, E.; Hodson, D.; Szabo, L.J. First Report of TTRTF Race of Wheat Stem Rust, Puccinia graminis f. sp. tritici, in Ethiopia. Plant Dis. 2020, 104, 293. [Google Scholar] [CrossRef]
- Omrani, A.; Roohparvar, R. First Report of TTRTF Race of the Wheat Stem Rust Pathogen, Puccinia graminis f. sp. tritici from Iran (Northwest, Cold Zone). J. Appl. Plant Prot. 2021, 9, 101–103. [Google Scholar]
- Hundie, B.; Girma, B.; Tadesse, Z.; Edae, E.; Olivera, P.; Abera, E.H.; Bulbula, W.D.; Abeyo, B.; Badebo, A.; Cisar, G.; et al. Characterization of Ethiopian Wheat Germplasm for Resistance to Four Puccinia graminis f. sp. tritici Races Facilitated by Single-Race Nurseries. Plant Dis. 2019, 103, 2359–2366. [Google Scholar] [CrossRef] [Green Version]
- Hovmøller, M.S.; Patpour, M.; Rodriguez-Algaba, J.; Thach, T.; Justesen, F.; Hansen, J.G. GRRC Report of Yellow and Stem Rust Genotyping and Race Analyses 2020, Flakkebjerg, DK-4200 Slagelse, Denmark. 2020, 12. Available online: https://agro.au.dk/forskning/internationale-platforme/wheatrust (accessed on 1 September 2022).
- Ghrabi, G.Z. Professor at the National Agronomic Institute of Tunisia, Tunis, Tunisia. Personal communication. 2021. [Google Scholar]
- RustTracker.org. A Global Wheat Rust Monitoring System. Stem Rust Pathotype Frequency Map. Available online: https://rusttracker.cimmyt.org/?page_id=26 (accessed on 1 September 2022).
- Meliani, N.; Dib, M.E.A.; Allali, H.; Tabti, B. Hypoglycaemic Effect of Berberis Vulgaris L. in Normal and Streptozotocin-Induced Diabetic Rats. Asian Pac. J. Trop. Biomed. 2011, 1, 468–471. [Google Scholar] [CrossRef] [Green Version]
- Mezouar, D. Recherche de l’activités Biologiques de Berberis Vulgaris. Master’s Thesis, University of Abou Bekr Belkaïd, Tlemcen, Algeria, 2012. [Google Scholar]
- BenBelkacem, A. Professor at the Institute of Agronomic Research of Algeria, Alger, Algeria. Personal communication. 2022. [Google Scholar]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A Decimal Code for the Growth Stages of Cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Peterson, R.F.; Campbell, A.B.; Hannah, A.E. A Diagrammatic Scale for Estimating Rust Intensity on Leaves and Stems of Cereals. Can. J. Res. 1948, 26, 496–500. [Google Scholar] [CrossRef]
- Park, R.F.; Hodson, D.P.; Nazari, K.; Hovmøller, M.; Fetch, T.; Jin, Y.; Pretorius, Z.; Szabo, L. BGRI. Wheat Rust Survey Protocols. 2013, 29. Available online: https://rusttracker.cimmyt.org/wp-content/uploads/2011/11/2013-Updated-BGRI-protocols-v2-web.pdf (accessed on 18 January 2023).
- Stoxen, S. Population Structure of Puccinia graminis f. sp. tritici in the United States. Master’s Thesis, University of Minnesota, Minnesota, MI, USA, 2012. [Google Scholar]
- Jin, Y.; Szabo, L.J.; Rouse, M.N.; Fetch, T.; Pretorius, Z.A.; Wanyera, R.; Njau, P. Detection of Virulence to Resistance Gene Sr36 Within the TTKS Race Lineage of Puccinia graminis f. sp. tritici. Plant Dis. 2009, 93, 367–370. [Google Scholar] [CrossRef] [Green Version]
- Zhong, S.; Leng, Y.; Friesen, T.L.; Faris, J.D.; Szabo, L.J. Development and Characterization of Expressed Sequence Tag-Derived Microsatellite Markers for the Wheat Stem Rust Fungus Puccinia graminis f. Sp. tritici. Phytopathology 2009, 99, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Rust-Stem: Differential Sets. Available online: https://www.fao.org/agriculture/crops/rust/stem/stem-pathotypetracker/stem-differentialsets/en/ (accessed on 16 September 2022).
- Stakman, E.C.; Stewart, D.M.; Loegering, W.Q. Identification of Physiologic Races of Puccinia graminis Var. tritici; USDA: Washington, DC, USA, 1962. [Google Scholar]
- Ripley, B.; Venables, B.; Bates, D.M.; Hornik, K.; Gebhardt, A.; Firth, D.; Ripley, M.B. Package ‘Mass’. Cran r 2013, 538, 113–120. [Google Scholar]
- Lüdecke, D.; Ben-Shachar, M.; Patil, I.; Waggoner, P.; Makowski, D. Performance: An R Package for Assessment, Comparison and Testing of Statistical Models. JOSS 2021, 6, 3139. [Google Scholar] [CrossRef]
- Perrier, X.; Jacquemoud-Collet, J.P. DARwin Software; Version 6; CIRAD: Paris, France, 2006; Available online: https://darwin.cirad.fr/index.php (accessed on 10 November 2022).




{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Df | Square Sum | F Value | p-Value |
|---|---|---|---|---|
| Sites | 4 | 2319.405 | 16.002 | <0.0001 *** |
| Accessions | 62 | 2864.556 | 1.057 | 0.3775 |
| Sites × Accessions | 205 | 6917.802 | 843.634 | 0.0274 * |
| Sowing Areas | ||||||
|---|---|---|---|---|---|---|
| Beja | Jendouba | Zaghouan | Mateur | Bizerte | ||
| Septoria Platform Experimental Station | Kodia-Septoria Platform Experimental Station | Farmer Field | Farmer Field | Farmer Field | ||
| 2020–2021 cropping season | ||||||
| Sowing date | 4-Nov | 6-Nov | 13-Nov | 15-Nov | 16-Nov | |
| N° of replicates/site | 1 | 1 | 1 | 1 | 1 | |
| 2021–2022 cropping season | ||||||
| Sowing date | Early | 26-Nov | 18-Nov | 23-Nov | 20-Nov | - |
| Late | 24-Dec | 23-Dec | - | - | - | |
| N° of replicates/site | 2 | 2 | 2 | 2 | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdedayem, W.; Patpour, M.; Laribi, M.; Justesen, A.F.; Kouki, H.; Fakhfakh, M.; Hovmøller, M.S.; Yahyaoui, A.H.; Hamza, S.; Ben M’Barek, S. Wheat Stem Rust Detection and Race Characterization in Tunisia. Plants 2023, 12, 552. https://doi.org/10.3390/plants12030552
Abdedayem W, Patpour M, Laribi M, Justesen AF, Kouki H, Fakhfakh M, Hovmøller MS, Yahyaoui AH, Hamza S, Ben M’Barek S. Wheat Stem Rust Detection and Race Characterization in Tunisia. Plants. 2023; 12(3):552. https://doi.org/10.3390/plants12030552
Chicago/Turabian StyleAbdedayem, Wided, Mehran Patpour, Marwa Laribi, Annemarie F. Justesen, Hajer Kouki, Moez Fakhfakh, Mogens S. Hovmøller, Amor H. Yahyaoui, Sonia Hamza, and Sarrah Ben M’Barek. 2023. "Wheat Stem Rust Detection and Race Characterization in Tunisia" Plants 12, no. 3: 552. https://doi.org/10.3390/plants12030552