

Anthocyanin Content and Fusarium Mycotoxins in Pigmented Wheat (Triticum aestivum L. spp. aestivum): An Open Field Evaluation
 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. Chemicals
3.2. Samples
3.3. Sample Preparation for Mycotoxin Analysis
3.4. UHPLC-MS/MS Mycotoxin Analysis
3.5. Extraction of Anthocyanins
3.6. UHPLC-MS/MS Anthocyanins Analysis
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gill, B.S.; Appels, R.; Botha-Oberholster, A.M.; Buell, C.R.; Bennetzen, J.L.; Chalhoub, B.; Chumley, F.; Dvořák, J.; Iwanaga, M.; Keller, B.; et al. A Workshop Report on Wheat Genome Sequencing: International Genome Research on Wheat Consortium. Genetics 2004, 168, 1087–1096. [Google Scholar] [CrossRef]
- Mourad, A.M.I.; Alomari, D.Z.; Alqudah, A.M.; Sallam, A.; Salem, K.F.M. Recent Advances in Wheat (Triticum Spp.) Breeding. In Advances in Plant Breeding Strategies: Cereals; Springer International Publishing: Cham, Switzerland, 2019; Volume 5, pp. 559–593. ISBN 9783030231088. [Google Scholar]
- Gupta, R.; Meghwal, M.; Prabhakar, P.K. Bioactive Compounds of Pigmented Wheat (Triticum Aestivum): Potential Benefits in Human Health. Trends Food Sci. Technol. 2021, 110, 240–252. [Google Scholar] [CrossRef]
- Himi, E.; Maekawa, M.; Miura, H.; Noda, K. Development of PCR Markers for Tamyb10 Related to R-1, Red Grain Color Gene in Wheat. Theor. Appl. Genet. 2011, 122, 1561–1576. [Google Scholar] [CrossRef] [PubMed]
- Tereshchenko, O.; Gordeeva, E.; Arbuzova, V.; Börner, A.; Khlestkina, E. The D Genome Carries a Gene Determining Purple Grain Colour in Wheat. Cereal Res. Commun. 2012, 40, 334–341. [Google Scholar] [CrossRef]
- Gordeeva, E.I.; Shoeva, O.Y.; Khlestkina, E.K. Marker-Assisted Development of Bread Wheat near-Isogenic Lines Carrying Various Combinations of Purple Pericarp (Pp) Alleles. Euphytica 2015, 203, 469–476. [Google Scholar] [CrossRef]
- Shoeva, O.Y.; Gordeeva, E.I.; Khlestkina, E.K. The Regulation of Anthocyanin Synthesis in the Wheat Pericarp. Molecules 2014, 19, 20266–20279. [Google Scholar] [CrossRef]
- Jeewani, D.C.; Zhonghua, W.; Wenhui, J.; Chandana Nishantha, M.D.L.; Wenzhi, N.; Tianxiang, L. Structural Genes Encoding F3′5′h in 4d Chromosome Control the Blue Grain Trait in Wheat. Trop. Agric. Res. Ext. 2021, 24, 35. [Google Scholar] [CrossRef]
- Qualset, C.O.; Soliman, K.M.; Jan, C.C.; Dvorak, J.; Vogt, H.E. Release of Wheat Germplasm: A Blue Aleurone Translocation Stock-UC66049; Agronomy Progress Report No. 139; University of California: Davis, CA, USA, 1983. [Google Scholar]
- Watanabe, N.; Martinek, P. Pyramiding Ba (Blue aleurone) genes to develop darker blue grain wheat: Prospect of nine-dose of Ba gene. Breed. Res. 2022, 24, 152. [Google Scholar]
- Lachman, J.; Martinek, P.; Kotíková, Z.; Orsák, M.; Šulc, M. Genetics and Chemistry of Pigments in Wheat Grain—A Review. J. Cereal Sci. 2017, 74, 145–154. [Google Scholar] [CrossRef]
- Saini, P.; Kumar, N.; Kumar, S.; Mwaurah, P.W.; Panghal, A.; Attkan, A.K.; Singh, V.K.; Garg, M.K.; Singh, V. Bioactive Compounds, Nutritional Benefits and Food Applications of Colored Wheat: A Comprehensive Review. Crit. Rev. Food Sci. Nutr. 2020, 61, 3197–3210. [Google Scholar] [CrossRef]
- Jia, Y.; Selva, C.; Zhang, Y.; Li, B.; McFawn, L.A.; Broughton, S.; Zhang, X.; Westcott, S.; Wang, P.; Tan, C.; et al. Uncovering the Evolutionary Origin of Blue Anthocyanins in Cereal Grains. Plant J. 2020, 101, 1057–1074. [Google Scholar] [CrossRef]
- Santos-Buelga, C.; Mateus, N.; de Freitas, V. Anthocyanins. Plant Pigments and Beyond. J. Agric. Food Chem. 2014, 62, 6879–6884. [Google Scholar] [CrossRef]
- Lin, Y.; Fan, L.; He, J.; Wang, Z.; Yin, Y.; Cheng, Y.; Li, Z. Anthocyanins Contribute to Fruit Defense against Postharvest Green Mold. Postharvest Biol. Technol. 2021, 181, 111661. [Google Scholar] [CrossRef]
- Landoni, M.; Puglisi, D.; Cassani, E.; Borlini, G.; Brunoldi, G.; Comaschi, C.; Pilu, R. Phlobaphenes Modify Pericarp Thickness in Maize and Accumulation of the Fumonisin Mycotoxins. Sci. Rep. 2020, 10, 1417. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, A.; Satyanarayana, K.V. Disease and Pest Resistance in Grains of Sorghum and Millets. J. Cereal Sci. 2006, 44, 287–304. [Google Scholar] [CrossRef]
- Spaggiari, M.; Righetti, L.; Galaverna, G.; Giordano, D.; Scarpino, V.; Blandino, M.; Dall’Asta, C. HR-MS Profiling and Distribution of Native and Modified Fusarium Mycotoxins in Tritordeum, Wheat and Barley Whole Grains and Corresponding Pearled Fractions. J. Cereal Sci. 2019, 87, 178–184. [Google Scholar] [CrossRef]
- Simsek, S.; Burgess, K.; Whitney, K.L.; Gu, Y.; Qian, S.Y. Analysis of Deoxynivalenol and Deoxynivalenol-3-Glucoside in Wheat. Food Control 2012, 26, 287–292. [Google Scholar] [CrossRef]
- McCormick, S.P.; Kato, T.; Maragos, C.M.; Busman, M.; Lattanzio, V.M.T.; Galaverna, G.; Dall-Asta, C.; Crich, D.; Price, N.P.J.; Kurtzman, C.P. Anomericity of T-2 Toxin-Glucoside: Masked Mycotoxin in Cereal Crops. J. Agric. Food Chem. 2015, 63, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; de Saeger, S.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked Mycotoxins: A Review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef]
- Atanasova-Penichon, V.; Barreau, C.; Richard-Forget, F. Antioxidant Secondary Metabolites in Cereals: Potential Involvement in Resistance to Fusarium and Mycotoxin Accumulation. Front. Microbiol. 2016, 7, 566. [Google Scholar] [CrossRef]
- Etzerodt, T.; Gislum, R.; Laursen, B.B.; Heinrichson, K.; Gregersen, P.L.; Jørgensen, L.N.; Fomsgaard, I.S. Correlation of Deoxynivalenol Accumulation in Fusarium-Infected Winter and Spring Wheat Cultivars with Secondary Metabolites at Different Growth Stages. J. Agric. Food Chem. 2016, 64, 4545–4555. [Google Scholar] [CrossRef]
- Bollina, V.; Kushalappa, A.C.; Choo, T.M.; Dion, Y.; Rioux, S. Identification of Metabolites Related to Mechanisms of Resistance in Barley against Fusarium Graminearum, Based on Mass Spectrometry. Plant Mol. Biol. 2011, 77, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Gunnaiah, R.; Kushalappa, A.C. Metabolomics Deciphers the Host Resistance Mechanisms in Wheat Cultivar Sumai-3, against Trichothecene Producing and Non-Producing Isolates of Fusarium Graminearum. Plant Physiol. Biochem. 2014, 83, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Choo, T.M.; Vigier, B.; Savard, M.E.; Blackwell, B.; Martin, R.; Wang, J.; Yang, J.; Abdel-Aal, E.S.M. Black Barley as a Means of Mitigating Deoxynivalenol Contamination. Crop. Sci. 2015, 55, 1096–1103. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef]
- Watanabe, N. Wheat Near-Isogenic Lines; Sankeisha: Nagoya, Japan, 2003; p. 156. [Google Scholar]
- Gordeeva, E.; Shamanin, V.; Shoeva, O.; Kukoeva, T.; Morgounov, A.; Khlestkina, E. The Strategy for Marker-Assisted Breeding of Anthocyanin-Rich Spring Bread Wheat (Triticum Aestivum l.) Cultivars in Western Siberia. Agronomy 2020, 10, 1603. [Google Scholar] [CrossRef]
- Chrpová, J.; Orsák, M.; Martinek, P.; Lachman, J.; Trávníčková, M. Potential Role and Involvement of Antioxidants and Other Secondary Metabolites of Wheat in the Infection Process and Resistance to Fusarium spp. Agronomy 2021, 11, 2235. [Google Scholar] [CrossRef]
- Ovando-Martínez, M.; Ozsisli, B.; Anderson, J.; Whitney, K.; Ohm, J.B.; Simsek, S. Analysis of Deoxynivalenol and Deoxynivalenol-3-Glucoside in Hard Red Spring Wheat Inoculated with Fusarium Graminearum. Toxins 2013, 5, 2522–2532. [Google Scholar] [CrossRef]
- Amarasinghe, C.C.; Simsek, S.; Brûlé-Babel, A.; Fernando, W.G.D. Analysis of Deoxynivalenol and Deoxynivalenol-3-Glucosides Content in Canadian Spring Wheat Cultivars Inoculated with Fusarium Graminearum. Food Addit. Contam. Part A 2016, 33, 1254–1264. [Google Scholar] [CrossRef]
- Jestoi, M. Emerging Fusarium-Mycotoxins Fusaproliferin, Beauvericin, Enniatins, and Moniliformin—A Review. Crit. Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar] [CrossRef]
- Martinek, P.; Škorpík, M.; Chrpová, J.; Fučík, P. Schweiger, J. Development of the New Winter Wheat Variety Skorpion with Blue Grain. Czech J. Genet. Plant Breed. 2013, 49, 90–94. [Google Scholar] [CrossRef] [Green Version]
- Morgounov, A.; Karaduman, Y.; Akin, B.; Aydogan, S.; Baenziger, P.S.; Bhatta, M.; Chudinov, V.; Dreisigacker, S.; Govindan, V.; Güler, S.; et al. Yield and Quality in Purple-Grained Wheat Isogenic Lines. Agronomy 2020, 10, 86. [Google Scholar] [CrossRef]
- Khlestkina, E.K. Genes Determining the Coloration of Different Organs in Wheat. Russ. J. Genet. Appl. Res. 2013, 3, 54–65. [Google Scholar] [CrossRef]
- Scarpino, V.; Blandino, M. Effects of Durum Wheat Cultivars with Different Degrees of Fhb Susceptibility Grown under Different Meteorological Conditions on the Contamination of Regulated, Modified and Emerging Mycotoxins. Microorganisms 2021, 9, 408. [Google Scholar] [CrossRef]
- Malachová, A.; Sulyok, M.; Beltrán, E.; Berthiller, F.; Krska, R. Optimization and Validation of a Quantitative Liquid Chromatography-Tandem Mass Spectrometric Method Covering 295 Bacterial and Fungal Metabolites Including All Regulated Mycotoxins in Four Model Food Matrices. J. Chromatogr. A 2014, 1362, 145–156. [Google Scholar] [CrossRef]
- Zaupa, M.; Calani, L.; del Rio, D.; Brighenti, F.; Pellegrini, N. Characterization of Total Antioxidant Capacity and (Poly)Phenolic Compounds of Differently Pigmented Rice Varieties and Their Changes during Domestic Cooking. Food Chem. 2015, 187, 338–347. [Google Scholar] [CrossRef]
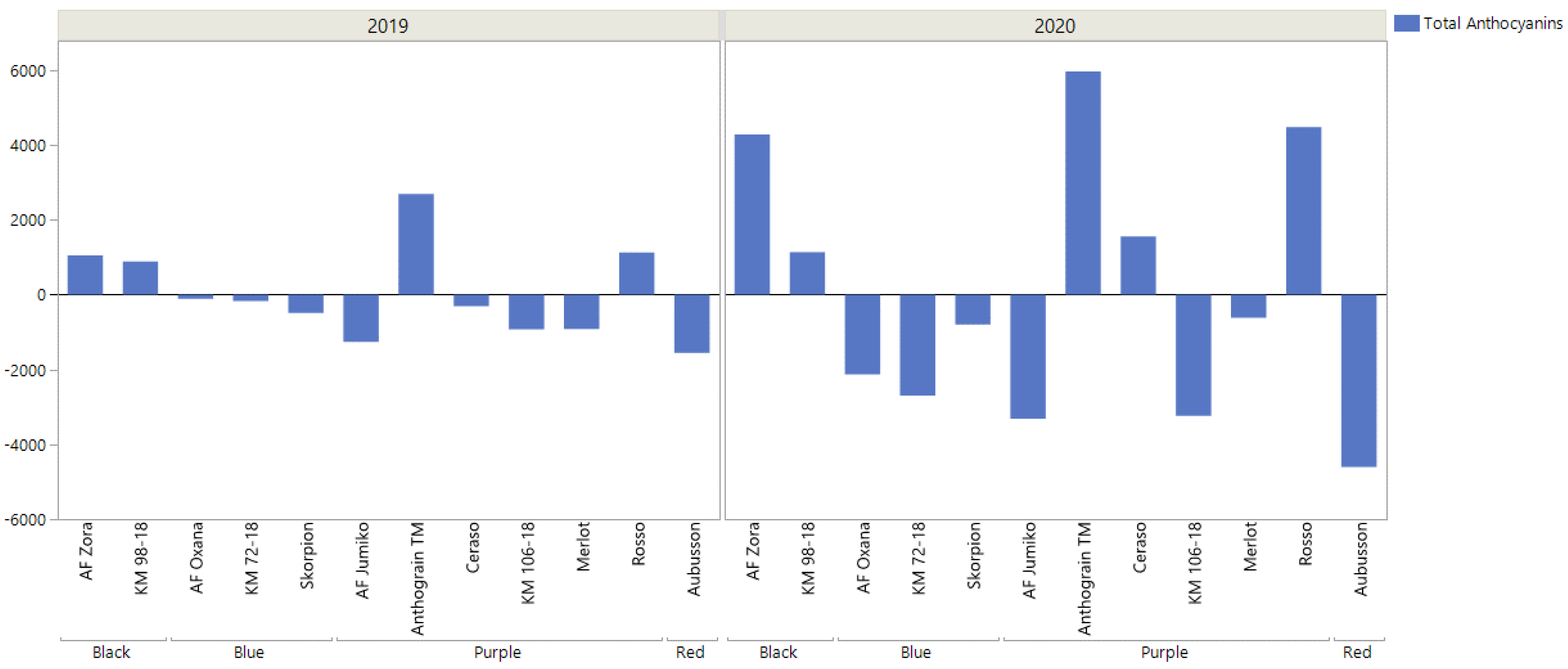


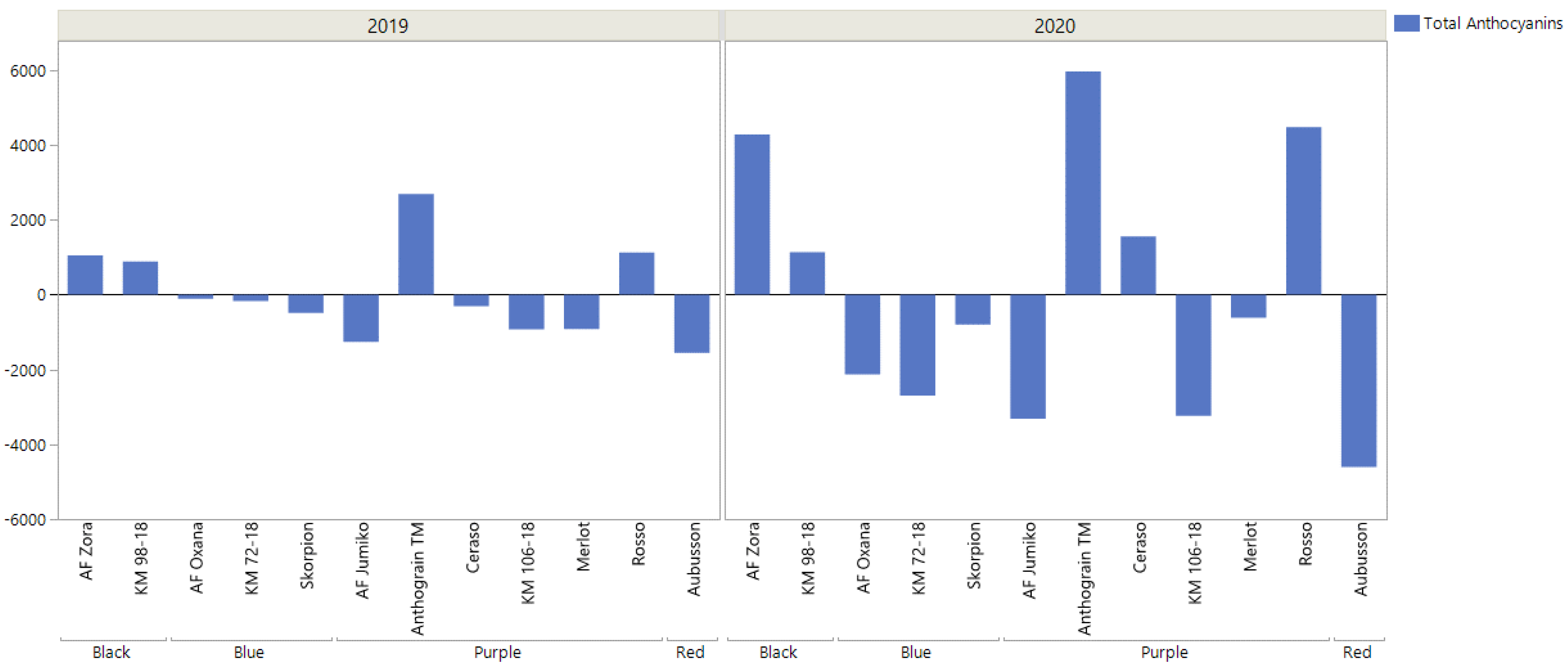{kind=link}
{kind=link}
| Growing Season | Month | Rainfall | Rainy Days | GDD 1 |
|---|---|---|---|---|
| (mm) | (days) | (Σ °C d−1) | ||
| 2018–2019 | November | 124 | 5 | 292 |
| December | 11 | 1 | 151 | |
| January | 6 | 1 | 141 | |
| February | 43 | 7 | 195 | |
| March | 17 | 4 | 314 | |
| April | 116 | 7 | 393 | |
| May | 178 | 9 | 478 | |
| June | 40 | 4 | 667 | |
| November–June | 535 | 39 | 2632 | |
| November–March | 200 | 19 | 1093 | |
| April–June | 335 | 21 | 1539 | |
| 2019–2020 | November | 314 | 12 | 249 |
| December | 132 | 8 | 193 | |
| January | 5 | 1 | 168 | |
| February | 1 | 0 | 229 | |
| March | 62 | 7 | 285 | |
| April | 81 | 10 | 414 | |
| May | 122 | 7 | 579 | |
| June | 113 | 7 | 624 | |
| November–June | 830 | 52 | 2741 | |
| November–March | 514 | 28 | 1124 | |
| April–June | 317 | 24 | 1617 |
| Year | Grain Colour | Variety | Cyanidin | Dephinidin | Peonidin | Petunidin | Malvidin | Pelargonidin | Total ACNs | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| µg kg−1 | |||||||||||||||||
| 3-O-Glc | 3-O-Rut | O-MalHex | 3-O-Glc | 3-O-Rut | 3-O-Glc | 3-O-Rut | O-MalHex | 3-O-Glc | 3-O-Rut | 3-O-Glc | 3-O-Glc | 3-O-Rut | O-MalHex | ||||
| 2020 | Black | AF Zora | 735 ± 36 b | 1867 ± 89 a | 398 ± 92 c | 622 ± 53 a | 1041 ± 97 a | 644 ± 149 b | 1583 ± 345 a | 1660 ± 409 b | 127 ± 6 a | 353 ± 27 a | 122 ± 0 | 86 ± 11 b | 99 ± 14 b | 34 ± 4 c | 9488 ± 930 |
| KM 98-18 | 415 ± 84 c | 1983 ± 399 ab | 225 ± 26 c | 419 ± 51 b | 1185 ± 18 9 a | 241 ± 35 c | 576 ± 78 c | 597 ± 90 d | 106 ± 5 a | 387 ± 51 a | 118 ± 2 | 27 ± 2 c | 30 ± 2 de | <LOD | 6345 ± 1012 | ||
| Blue | Skorpion | 278 ± 48 d | 1449 ± 240 b | 93 ± 21 d | 323 ± 28 c | 838 ± 93 b | 129 ± 20 d | 420 ± 66 c | 251 ± 55 e | 99 ± 4 ab | 293 ± 27 b | 113 ± 1 | 32 ± 4 c | 41 ± 7 d | <LOQ | 4372 ± 630 | |
| AF Oxana | 146 ± 29 d | 967 ± 213 c | 29 ± 3 e | 289 ± 37 c | 817 ± 152 b | 58 ± 4 d | 236 ± 48 d | 61 ± 6 f | 90 ± 4 b | 253 ± 42 b | 111 ± 4 | <LOQ | 25 ± 1 e | 25 ± 0 c | 3074 ± 551 | ||
| KM 72-18 | 164 ± 45 d | 752 ± 273 c | 32 ± 8 e | 255 ± 40 c | 508 ± 123 c | 66 ± 11 d | 217 ± 50 d | 74 ± 30 f | 90 ± 5 b | 197 ± 45 bc | 109 ± 3 | <LOD | 27 ± 0 e | 150 ± 27 b | 2453 ± 591 | ||
| Purple | AnthograinTM CDC | 1362 ± 180 a | 1459 ± 154 b | 2194 ± 239 a | <LOD | <LOD | 1354 ± 185 a | 729 ± 81 b | 3517 ± 341 a | <LOD | 72 ± 0.2 c | 111 ± 3 | 97 ± 9 b | 64 ± 5 c | 37 ± 2 c | 11110 ± 1158 | |
| Ceraso | 451 ± 31 c | 2077 ± 282 a | 281 ± 19 c | 412 ± 33 b | 1211 ± 172 a | 287 ± 24 c | 593 ± 83 c | 758 ± 90 c | 104 ± 5 a | 379 ± 51 a | 118 ± 3 | 29 ± 1 c | 31 ± 2 d | 35 ± 2 c | 6766 ± 720 | ||
| Jumiko | 87 ± 9 e | 124 ± 6 e | 142 ± 16 d | <LOD | 118 ± 5 d | 273 ± 27 c | 262 ± 19 d | 735 ± 77 c | <LOD | 74 ± 1 c | <LOD | 26 ± 0 c | 25 ± 0 d | 50 ± 4 c | 1891 ± 135 | ||
| KM 106-18 | 94 ± 14 e | 206 ± 28 e | 169 ± 12 d | <LOD | 122 ± 6 d | 224 ± 24 c | 321 ± 41 d | 650 ± 54 cd | <LOD | 75 ± 2 c | <LOD | 30 ± 2 c | 30 ± 3 d | 174 ± 42 b | 1970 ± 185 | ||
| Merlot | 244 ± 41 d | 464 ± 74 d | 277 ± 50 c | <LOD | 111 ± 0 d | 587 ± 122 b | 842 ± 157 b | 1665 ± 368 b | <LOD | 75 ± 1 c | <LOD | 77 ± 14 b | 79 ± 14 bc | 365 ± 16 a | 4471 ± 823 | ||
| Rosso | 571 ± 14 c | 1443 ± 94 b | 794 ± 34 b | <LOD | <LOD | 1071 ± 20 a | 1699 ± 82 a | 3367 ± 104 a | <LOD | 77 ± 1 c | <LOD | 135 ± 3 a | 167 ± 10 a | 49 ± 9 c | 9664 ± 348 | ||
| Red | Aubusson | 28 ± 5 f | 59 ± 32 f | 25 ± 1 e | 118 ± 0 d | 125 ± 22 d | 52 ± 1 d | 57 ± 3 e | 66 ± 4 f | <LOQ | 80 ± 1 c | <LOD | <LOQ | <LOQ | 212 ± 14 ab | 437 ± 206 | |
| Mean ± SD | 381 ± 376 | 1071 ± 742 | 388 ± 680 | 348 ± 158 | 608 ± 465 | 416 ± 422 | 628 ± 526 | 1117 ± 1215 | 103 ± 14 | 193 ± 133 | 115 ± 5 | 60 ± 40 | 56 ± 44 | 113 ± 12 | 5170 ± 3495 | ||
| 3-O-Glc | 3-O-Rut | O-MalHex | 3-O-Glc | 3-O-Rut | 3-O-Glc | 3-O-Rut | O-MalHex | 3-O-Glc | 3-O-Rut | 3-O-Glc | 3-O-Glc | 3-O-Rut | O-MalHex | Total ACNs | |||
| 2019 | Black | AF Zora | 151 ± 14 b | 413 ± 54 a | 97 ± 25 c | 214 ± 28 a | 324 ± 67 ab | 209 ± 51 b | 509 ± 76 a | 509 ± 145 b | 78 ± 5 | 163 ± 35 | 95 ± 3 | 13 ± 4 | 13 ± 2 | 4 ± 2 | 2818 ± 161 |
| KM 98-18 | 129 ± 7 b | 596 ± 52 a | 111 ± 34 b | 194 ± 18 ab | 474 ± 43 a | 125 ± 26 c | 296 ± 14 bc | 321 ± 97 c | 75 ± 3 | 228 ± 14 | 96 ± 0 | <LOQ | <LOQ | <LOQ | 2650 ± 86 | ||
| Blue | Skorpion | 38 ± 6 d | 275 ± 57 b | 7 ± 2 e | 136 ± 5 b | 252 ± 34 b | 44 ± 4 d | 166 ± 38 c | 57 ± 14 e | 70 ± 1 | 137 ± 19 | 92 ± 3 | <LOQ | <LOQ | <LOQ | 1275 ± 149 | |
| AF Oxana | 47 ± 3 d | 404 ± 26 a | 3 ± 1 e | 175 ± 9 b | 423 ± 20 a | 37 ± 1 d | 175 ± 8 c | 38 ± 6 e | 72 ± 1 | 185 ± 9 | 92 ± 1 | <LOQ | <LOQ | <LOD | 1639 ± 51 | ||
| KM 72-18 | 73 ± 36 c | 393 ± 156 ab | 3 ± 2 e | 186 ± 45 ab | 324 ± 96 ab | 42 ± 5 d | 187 ± 55 c | 36 ± 3 e | 78 ± 8 | 176 ± 49 | 96 ± 6 | <LOQ | <LOQ | 31 ± 8 | 1593 ± 461 | ||
| Purple | AnthograinTM CDC | 436 ± 56 a | 381 ± 73 b | 711 ± 64 a | <LOD | <LOQ | 655 ± 70 a | 329 ± 58 b | 1789 ± 140 a | 63 ± 2 | <LOD | <LOD | 22 ± 3 | 6 ± 2 | 65 ± 9 | 4451 ± 457 | |
| Ceraso | 65 ± 13 c | 131 ± 21 c | 118 ± 24 b | <LOD | 88 ± 2 c | 195 ± 34 bc | 196 ± 39 c | 572 ± 84 b | <LOQ | <LOD | <LOD | 19 ± 3 | 11 ± 3 | <LOQ | 1458 ± 229 | ||
| Jumiko | 14 ± 6 e | 53 ± 3 d | 28 ± 13 d | <LOD | 89 ± 2 c | 79 ± 25 d | 56 ± 11 e | 186 ± 77 d | <LOQ | <LOD | <LOD | <LOQ | <LOD | 9 ± 1 | 504 ± 133 | ||
| KM 106-18 | 33 ± 5 d | 73 ± 4 d | 72 ± 9 c | <LOD | 86 ± 0 c | 123 ± 9 c | 97 ± 9 d | 345 ± 30 c | <LOQ | <LOD | <LOD | <LOQ | <LOQ | 18 ± 2 | 780 ± 72 | ||
| Merlot | 33 ± 3 d | 77 ± 4 d | 37 ± 3 d | <LOD | 88 ± 2 c | 147 ± 5 c | 107 ± 7 d | 333 ± 17 c | <LOQ | <LOD | <LOD | 5 ± 0.4 | <LOQ | 83 ± 45 | 844 ± 41 | ||
| Rosso | 167 ± 85 b | 258 ± 122 bc | 222 ± 109 b | <LOD | 86 ± 0 c | 381 ± 164 b | 419 ± 206 ab | 1238 ± 565ab | <LOQ | <LOD | <LOD | 23 ± 13 | 17 ± 12 | <LOQ | 2866 ± 1276 | ||
| Red | Aubusson | <LOQ | 49 ± 0 d | <LOQ | <LOD | 86 ± 0 c | <LOQ | 35 ± 0 e | 39 ± 2 e | <LOQ | <LOD | <LOD | <LOQ | <LOD | 59 ± 5 | 82 ± 112 | |
| Mean ± SD | 108 ± 120 | 258 ± 182 | 128 ± 204 | 181 ± 29 | 211 ± 153 | 185 ± 185 | 215 ± 146 | 455 ± 540 | 73 ± 6 | 178 ± 33 | 94 ± 2 | 16 ± 8 | 12 ± 51 | 39 ± 31 | 1747 ± 1240 | ||
| Year | Grain Colour | Variety | DON | DON3Glc | 3/15Ac-DON | ZEN | Enniatin B | T-2 | HT-2 |
|---|---|---|---|---|---|---|---|---|---|
| mg kg−1 | |||||||||
| 2020 | Black | AF Zora | 2.327 ± 1.021 d | 0.101 ± 0.034 d | <LOD | 0.012 ± 0.000 d | 0.247 ± 0.138 d | <LOQ | <LOD |
| KM 98-18 | 3.280 ± 2.093 c | 0.141 ± 0.060 c | <LOD | 0.045 ± 0.014 b | 0.518 ± 0.388 b | <LOQ | <LOQ | ||
| Blue | Skorpion | 7.235 ± 2.594 b | 0.289 ± 0.060 b | <LOD | 0.098 ± 0.047 a | 0.418 ± 0.317 c | <LOQ | <LOQ | |
| AF Oxana | 9.603 ± 2.767 a | 0.357 ± 0.092 a | <LOD | 0.088 ± 0.037 a | 0.681 ± 0.314 b | <LOQ | <LOQ | ||
| KM 72-18 | 6.769 ± 3.163 b | 0.329 ± 0.103 a | <LOD | 0.057 ± 0.020 b | 0.951 ± 0.366 a | <LOQ | <LOQ | ||
| Purple | AnthograinTM CDC | 1.466 ± 0.561 d | 0.084 ± 0.010 d | <LOD | 0.028 ± 0.013 c | 0.281 ± 0.227 d | <LOD | <LOD | |
| Ceraso | 2.333 ± 1.406 d | 0.100 ± 0.054 d | <LOD | 0.034 ± 0.007 c | 0.554 ± 0.509 b | <LOD | <LOQ | ||
| Jumiko | 2.562 ± 0.271 d | 0.177 ± 0.017 c | <LOD | 0.023 ± 0.021 c | 0.317 ± 0.192 c | <LOD | <LOD | ||
| KM 106-18 | 3.889 ± 1.008 c | 0.174 ± 0.033 c | <LOD | 0.040 ± 0.032 b | 0.416 ± 0.103 c | <LOQ | <LOQ | ||
| Merlot | 3.460 ± 0.494 c | 0.152 ± 0.030 c | <LOD | 0.032 ± 0.015 c | 0.194 ± 0.071 de | <LOD | <LOD | ||
| Rosso | 1.555 ± 0.247 d | 0.107 ± 0.028 d | <LOD | 0.027 ± 0.014 c | 0.129 ± 0.066 e | <LOD | <LOD | ||
| Red | Aubusson | 0.471 ± 0.128 e | 0.033 ± 0.007 e | <LOD | 0.01 ± 0.000 d | 0.125 ± 0.057 e | <LOD | <LOD | |
| Mean ± SD | 3.746 ± 2.991 | 0.170 ± 0.109 | n.d. | 0.046 ± 0.033 | 0.403 ± 0.326 | n.d. | n.d | ||
| 2019 | Black | AF Zora | 1.058 ± 0.176 cd | 0.054 ± 0.013 b | <LOD | <LOD | 0.083 ± 0.011 c | <LOD | <LOD |
| KM 98-18 | 1.046 ± 0.299 cd | 0.046 ± 0.013 c | <LOD | <LOQ | 0.121 ± 0.068 b | <LOD | <LOQ | ||
| Blue | Skorpion | 3.212 ± 0.772 a | 0.127 ± 0.026 a | <LOD | 0.081 ± 0.000 | 0.130 ± 0.047 ab | <LOD | <LOD | |
| AF Oxana | 3.922 ± 0.266 a | 0.105 ± 0.018 a | <LOD | <LOD | 0.103 ± 0.012 b | <LOD | <LOQ | ||
| KM 72-18 | 1.377 ± 0.352 c | 0.059 ± 0.021 b | <LOD | <LOQ | 0.171 ± 0.043 a | <LOD | 0.021 ± 0.000 | ||
| Purple | AnthograinTM CDC | 0.539 ± 0.121 f | 0.030 ± 0.000 d | <LOD | <LOD | 0.034 ± 0.012 d | <LOQ | <LOQ | |
| Ceraso | 0.653 ± 0.199 e | <LOQ | <LOD | 0.010 ± 0.000 | 0.026 ± 0.000 e | <LOD | <LOD | ||
| Jumiko | 0.752 ± 0.060 e | 0.029 ± 0.006 d | <LOD | <LOD | 0.044 ± 0.014 d | <LOD | <LOD | ||
| KM 106-18 | 0.960 ± 0.361 d | 0.033 ± 0.008 d | <LOD | <LOD | 0.039 ± 0.005 d | <LOD | <LOD | ||
| Merlot | 2.039 ± 0.986 bc | 0.053 ± 0.015 b | <LOD | <LOD | 0.022 ± 0.000 e | <LOD | <LOD | ||
| Rosso | 1.040 ± 0.449 | 0.057 ± 0.013 b | <LOD | <LOD | 0.038 ± 0.027 d | <LOD | <LOD | ||
| Red | Aubusson | 0.961 ± 0.114 d | 0.054 ± 0.015 b | <LOD | <LOQ | 0.058 ± 0.037 | <LOQ | <LOQ | |
| Mean ± SD | 1.463 ± 1.096 | 0.053 ± 0.034 | n.d. | 0.052 ± 0.021 | 0.079 ± 0.011 | n.d. | n.d. | ||
| Grain Color | Variety | Status | Registration | Responsible of Selection |
|---|---|---|---|---|
| Red (control) | Aubusson | Cultivar | 2003 | Limagrain Italia S.p.A., Fidenza (PR), Italy |
| Purple | AnthograinTM CDC | Cultivar | 2018 | Hetland Seeds Ltd., Naicam, SK, Canada |
| Ceraso | Cultivar | 2014 | Saatzucht Donau GesmbH. and CoKG, Austria | |
| AF Jumiko | Cultivar | 2018 | Agrotest Fyto, Ltd., Kroměříž, Czech Republic | |
| KM 106-18 | Breeding line | Agrotest Fyto, Ltd., Kroměříž, Czech Republic | ||
| Merlot | Cultivar | 2015 | Saatzucht Donau GesmbH. and CoKG, Austria | |
| Rosso | Cultivar | 2011 | Saatzucht Donau GesmbH. and CoKG, Austria | |
| Blue | Skorpion | Cultivar | 2011 1 | Agrotest Fyto, Ltd., Kroměříž, Czech Republic |
| AF Oxana | Cultivar | 2019 | Agrotest Fyto, Ltd., Kroměříž, Czech Republic | |
| KM 72-18 | Breeding line | Agrotest Fyto, Ltd., Kroměříž, Czech Republic | ||
| Black | AF Zora | Cultivar | 2021 | Agrotest Fyto, Ltd., Kroměříž, Czech Republic |
| KM 98-18 | Breeding line | Agrotest Fyto, Ltd., Kroměříž, Czech Republic |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gozzi, M.; Blandino, M.; Dall’Asta, C.; Martinek, P.; Bruni, R.; Righetti, L. Anthocyanin Content and Fusarium Mycotoxins in Pigmented Wheat (Triticum aestivum L. spp. aestivum): An Open Field Evaluation. Plants 2023, 12, 693. https://doi.org/10.3390/plants12040693
Gozzi M, Blandino M, Dall’Asta C, Martinek P, Bruni R, Righetti L. Anthocyanin Content and Fusarium Mycotoxins in Pigmented Wheat (Triticum aestivum L. spp. aestivum): An Open Field Evaluation. Plants. 2023; 12(4):693. https://doi.org/10.3390/plants12040693
Chicago/Turabian StyleGozzi, Marco, Massimo Blandino, Chiara Dall’Asta, Petr Martinek, Renato Bruni, and Laura Righetti. 2023. "Anthocyanin Content and Fusarium Mycotoxins in Pigmented Wheat (Triticum aestivum L. spp. aestivum): An Open Field Evaluation" Plants 12, no. 4: 693. https://doi.org/10.3390/plants12040693
APA StyleGozzi, M., Blandino, M., Dall’Asta, C., Martinek, P., Bruni, R., & Righetti, L. (2023). Anthocyanin Content and Fusarium Mycotoxins in Pigmented Wheat (Triticum aestivum L. spp. aestivum): An Open Field Evaluation. Plants, 12(4), 693. https://doi.org/10.3390/plants12040693