

An Update of the Cenchrinae (Poaceae, Panicoideae, Paniceae) and a New Genus for the Subtribe to Clarify the Dubious Position of a Species of Panicum L.

 , ,
, ,
Abstract
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
3. Discussion
3.1. Monophyly of the Cenchrinae
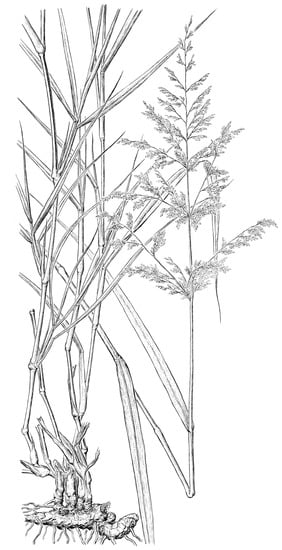
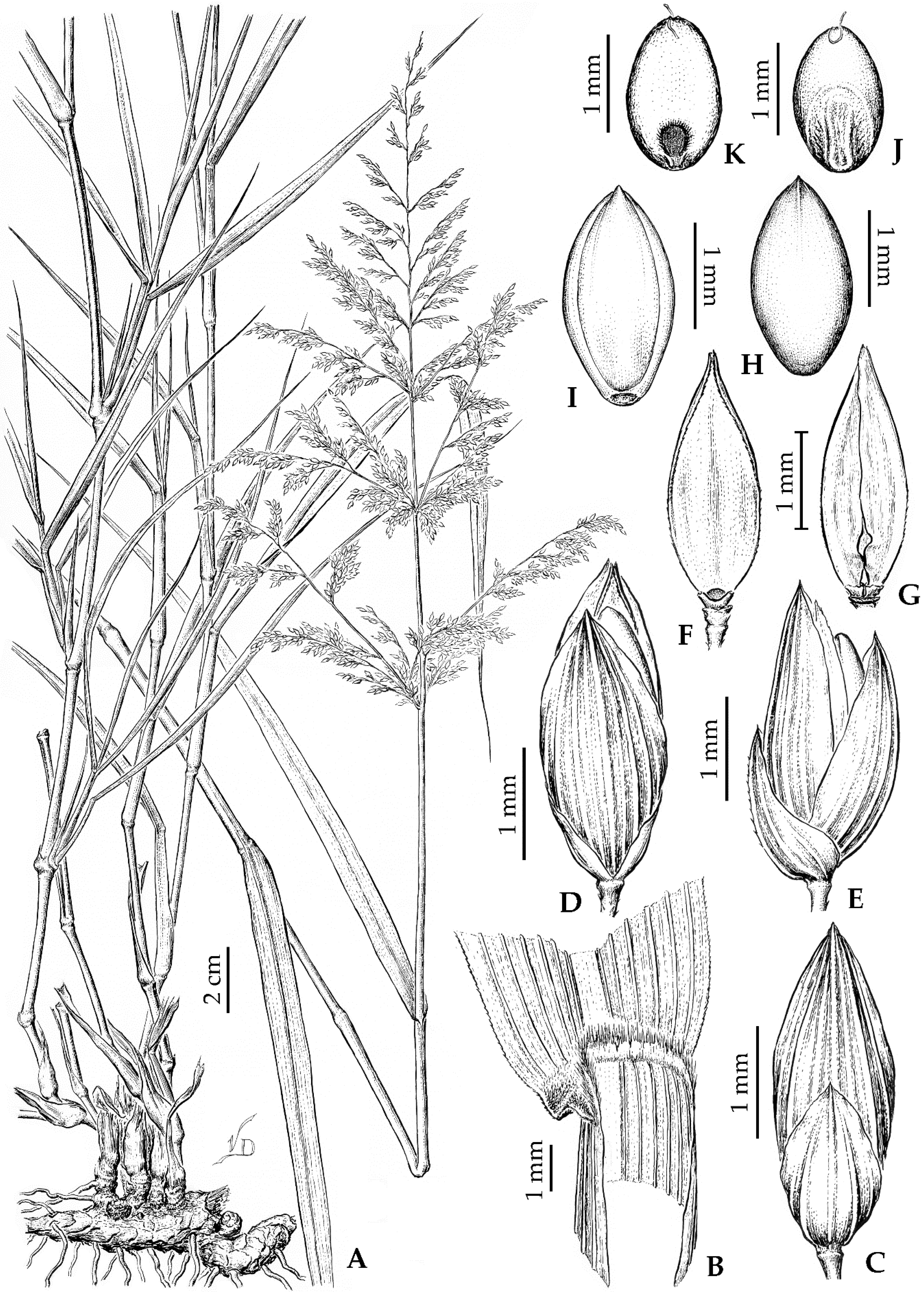
3.2. Panicum antidotale Retz., an Odd Species in the Subtribe Cenchrinae
| Key to Genera of the Subtribe Cenchrinae: |
| 1. Inflorescence subtended by a spatheole…………………………………………………….2 1′. Inflorescence without a spatheole below…………………………………………………..4 2(1). Plants bisexual…………………………………………………………………..Xerochloa 2′. Plants dioecious……………………………………………………………………………….3 3. Male spikelets different from female ones; male inflorescence a single raceme…………………………………………………………………………….……....Spinifex 3′. Male spikelets similar to the female ones; male inflorescence similar to the female one……………………………………………………………………………………...Zygochloa 4(1′). Inflorescence composed of short racemes with spikelets embedded in a thickened central axis…………………………………………………………………………………Stenotaphrum 4′. Inflorescence without spikelets embedded in a thickened central axis…………………5 5(4′). Inflorescence composed of paired or digitate racemes…………………Stereochlaena 5′. Inflorescence open to contracted or spiciform, not composed of paired or digitate racemes……………………………………………………………………………………………6 6(5′). Spikelets with two fertile florets. Endemic of Hawaii…………………Dissochondrus 6′. Spikelets with a male, or neuter, floret below and a fertile floret apically (or male and female spikelets in different inflorescences)…………………………………………………..7 7(6′). Species with sexes segregated on unisexual branches; male spikelets above and female spikelets below………………………………………………………………………………....Hygrochloa 7′. Species without sexes segregated on unisexual branches………………………………...8 8(7′). Caryopsis with a linear hilum……………………………………………..Pseudoraphis 8′. Caryopsis with a punctiform to oblong hilum…………………………………………….9 9(8′). Inflorescence with disarticulation at maturity at the base of the primary branches or at the nodes of the inflorescence axis………………………………………………………………………………………………..10 9′. Inflorescence not disarticulating at the base of the primary branches…………………15 10(9). Lower palea winged, expanded at maturity. Africa…………………...Uranthoecium 10′. Lower palea not expanded at maturity. Australia……………………………………...11 11(10′). Lower glume developed, 3–7-nerved; upper glume ovate, 11–19-nerved (except Pseudochaetochloa, with upper glume 3-nerved and plants perennial); plants annual…...12 11′. Lower glume absent or obscure, nerveless; upper glume 0–7-nerved; plants perennial…………………………………………………………………………………………14 12(11). Upper glume 3-nerved; plants perennial………………………..Pseudochaetochloa 12′. Upper glume 11–19-nerved; plants annual……………………………………………...13 13(12′). Spikelets surrounded by a fan of bristles……………………………….Plagiosetum 13′. Bristle single…………………………………………………………………...Paractaenum 14(11′). Inflorescences of racemes in a bilateral false spike; lower glume linear, almost as long as the lower lemma, 7-nerved. Australia………………………………………………………………………..…Chamaeraphis 14′. Inflorescences of racemes in a multilateral false spike; lower glume reduce; nerveless. Africa and America………………………………………………………………………………..Paratheria 15(9′). Inflorescences without bristles………………………………………………………...16 15′. Inflorescences subtended with 1 to many bristles or spines…………………………...19 16(15). Spikelets laterally compressed………………………………………………………...............................................17 16′. Spikelets dorsiventrally compressed……………………………………………………..18 17(16). Ligule membranous-ciliate; inflorescence composed of unilateral racemes; upper glume and lower lemma 5–7-nerved; upper anthecium stipitate, rugose, and coriaceous…………………………………………………………………………...Whiteochloa 17′. Ligule ciliate; inflorescence with spikelets arranged in open, not racemose branches; upper glume and lower lemma 7–9-nerved; upper anthecium not stipitate, smooth, cartilaginous………………………………………………………………………….Alexfloydia 18(16′). Plants with the lowest internodes developed into a corm-like base, without strong rhizomes; internodes hollow; ligules membranous; upper anthecium rugose…………………………………………………………………………………..Zuloagaea 18′. Plants without corm-like bases, strongly rhizomatous; internodes solid; ligules membranous-ciliate; upper anthecium smooth…………………………………….Janochloa 19(15′). Bristles persistent on the inflorescence………………………………………………20 19′. Bristles or spines falling with the spikelets…………………………………………...…21 20(19). Lower palea expanded at maturity; C3 genus…………………………..Holcolemma 20′. Lower palea not expanded at maturity; C4 genus………………………………...Setaria 21(19′). Spikelet subtended by a single bristle………………………………………………22 21′. Spikelet surrounded by an involucre of bristles………………………………………...23 22(21). Panicle contracted, spiciform, first order branches appressed, short; lower palea not expanded at maturity…………………………………………………………………………….…Setariopsis 22′. Panicle open, first order branches divergent, long; lower palea expanded and winged at maturity………………………………………………………………………………...Ixophorus 23(21′). Inflorescence with distant racemes-like branches; leaf blades pseudopetiolate…………………………………………………………………...Streptolophus 23′. Inflorescence spiciform; leaf blades not pseudopetiolate……………………...Cenchrus |
Taxonomic Treatment
3.3. Setaria P. Beauv.
3.4. Cenchrus and Related Taxa
3.5. Stenotaphrum Trin.
4. Materials and Methods
4.1. Molecular Studies
4.1.1. Taxon Sampling
4.1.2. DNA Amplification and Sequencing
4.1.3. Phylogenetic Analyses
4.2. Taxonomic Treatment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morrone, O.; Aagesen, L.; Scataglini, M.A.; Salariato, D.L.; Denham, S.S.; Chemisquy, M.A.; Sede, S.M.; Giussani, L.M.; Zuloaga, F.O. Phylogeny of the Paniceae (Poaceae: Panicoideae): Integrating plastid DNA sequences and morphology into a new classification. Cladistics 2012, 28, 333–356. [Google Scholar] [CrossRef]
- Burke, S.V.; Wysocki, W.P.; Zuloaga, F.O.; Craine, J.M.; Pires, J.C.; Edger, P.P.; Mayfield-Jones, D.; Clark, L.G.; Kelchner, S.A.; Duvall, M.R. Evolutionary relationships in Panicoid grasses based on plastome phylogenomics (Panicoideae; Poaceae). BMC Plant Biol. 2016, 16, 140. [Google Scholar] [CrossRef] [PubMed]
- Hackel, J.; Vorontsova, M.S.; Nanjarisoa, O.P.; Hall, R.C.; Razanatsoa, J.; Malakasi, P.; Besnard, G. Grass diversification in Madagascar: In situ radiation of two large C3 shade clades and support for a Miocene to Pliocene origin of C4 grassy biomes. J. Biogeogr. 2018, 45, 750–761. [Google Scholar] [CrossRef]
- Saarela, J.M.; Burke, S.V.; Wysocki, W.P.; Barrett, M.D.; Clark, L.G.; Craine, J.M.; Peterson, P.M.; Soreng, R.J.; Vorontsova, M.S.; Durvall, M.R. A 250 plastome phylogeny of the grass family (Poaceae): Topological support under different data partitions. PeerJ 2018, 6, e4299. [Google Scholar] [CrossRef]
- Gallaher, T.J.; Peterson, P.M.; Soreng, R.J.; Zuloaga, F.O.; Li, D.-Z.; Clark, L.G.; Tyrrell, C.D.; Welker, C.A.D.; Kellogg, E.A.; Teisher, J.K. Grasses through space and time: An overview of the biogeographical and macroevolutionary history of Poaceae. J. Syst. Evol. 2022, 60, 522–569. [Google Scholar] [CrossRef]
- Gutierrez, M.; Gracen, V.E.; Edwards, G.E. Biochemical and cytological relationships in C4 plants. Planta 1974, 119, 279–300. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; Tai, Y.S.; Liu, D.J.; Ku, M.S.B. Photosynthetic Mechanisms of Weeds in Taiwan. Aust. J. Plant Physiol. 1993, 20, 757–769. [Google Scholar] [CrossRef]
- Prendergast, H.D.V.; Hattersley, P.W.; Stone, N.E. New Structural/Biochemical Associations in Leaf Blades of C4 Grasses (Poaceae). Aust. J. Plant Physiol. 1987, 14, 403–420. [Google Scholar] [CrossRef]
- GPWG II (Grass Phylogeny Working Group II). New grass phylogeny resolves deep evolutionary relationships and discovers C4 origins. New Phytol. 2012, 193, 304–312. [Google Scholar] [CrossRef]
- Zuloaga, F.O.; Salariato, D.L.; Scataglini, A. Molecular phylogeny of Panicum s. str. (Poaceae, Panicoideae, Paniceae) and insights into its biogeography and evolution. PLoS ONE 2018, 13, e0191529. [Google Scholar] [CrossRef]
- Zuloaga, F.O. Systematics of the New World species of Panicum (Poaceae: Paniceae). In Grass Systematics and Evolution; Soderstrom, T.R., Hilu, K.W., Campbell, C.S., Barkworth, M.E., Eds.; Smithsonian Inst. Press: Washington, DC, USA, 1987; Chapter 26; pp. 287–306. [Google Scholar]
- Donadio, S.; Giussani, L.M.; Kellogg, E.A.; Zuloaga, F.O.; Morrone, O. A preliminary molecular phylogeny of Pennisetum and Cenchrus (Poaceae-Paniceae) based on the trnL-F, rpl16 chloroplast markers. Taxon 2009, 58, 392–404. [Google Scholar] [CrossRef]
- Chemisquy, M.A.; Giussani, L.M.; Scataglini, M.A.; Kellogg, E.A.; Morrone, O. Phylogenetic studies favour the unification of Pennisetum, Cenchrus and Odontelytrum (Poaceae): A combined nuclear, plastid and morphological analysis, and nomenclatural combinations in Cenchrus. Ann. Bot. 2010, 106, 107–130. [Google Scholar] [CrossRef]
- Soreng, R.J.; Peterson, P.M.; Zuloaga, F.O.; Romaschenko, K.; Clark, L.G.; Teisher, J.K.; Gillespie, L.J.; Barberá, P.; Welker, C.A.D.; Kellogg, E.A. A worldwide phylogenetic classification of the Poaceae (Gramineae) III: An update. J. Syst. Evol. 2022, 60, 476–521. [Google Scholar] [CrossRef]
- Kellogg, E.A. Evolution of Setaria. In Genetics and Genomics of Setaria, Plant Genetics and Genomics: Crops and Models; Doust, A., Diao, X., Eds.; Springer: Cham, Switzerland, 2017; Volume 19, pp. 3–27. [Google Scholar] [CrossRef]
- Webster, R.D. Nomenclature of Setaria (Poaceae: Paniceae). Sida 1993, 15, 447–489. [Google Scholar]
- Morrone, O.; Aliscioni, S.S.; Veldkamp, J.F.; Pensiero, J.F.; Zuloaga, F.O.; Kellogg, E.A. Revision of the Old World Species of Setaria (Poaceae: Panicoideae: Paniceae). Syst. Bot. Monogr. 2014, 96, 1–161. [Google Scholar]
- Pensiero, J.F. Las especies sudamericanas del género Setaria (Poaceae, Paniceae). Darwiniana 1999, 37, 37–151. [Google Scholar] [CrossRef]
- Stapf, O.; Hubbard, O.E. Setaria P. Beauv. In Flora of tropical Africa, Gramineae (Maydeae-Paniceae); Oliver, D., Ed.; Reeve & Co: Ashford, UK, 1930; Volume 9, pp. 768–866. [Google Scholar]
- Rominger, J.M.D. Taxonomy of Setaria (Gramineae) in North America; University of Illinois at Urbana-Champaign: Champaign, IL, USA, 1959; Volume 29, pp. 1–132. [Google Scholar] [CrossRef]
- BFG (The Brazil Flora Group). Brazilian Flora 2020: Leveraging the power of a collaborative scientific network. Taxon 2021, 71, 178–198. [Google Scholar] [CrossRef]
- Callmander, M.W. Catalogue of the Vascular Plants of Madagascar (Madagascar Catalogue), Missouri Botanical Garden and Antananarivo. Available online: http://legacy.tropicos.org/Project/Madagascar (accessed on 10 June 2022).
- Sousa, V.F.; Santos, C.A.G.; Boldrini, I.I. Setaria in Flora e Funga do Brasil, Jardim Botânico do Rio de Janeiro. Available online: http://floradobrasil.jbrj.gov.br/reflora/floradobrasil/FB13581 (accessed on 31 March 2022).
- Clayton, W.D.; Renvoize, S.A. Genera Graminum. Grasses of the World. Kew Bull. Addit. Ser. 1986, 13, 389. [Google Scholar]
- Stapf, O. Paspalidium Stapf. In Flora of Tropical Africa, Gramineae (Maydeae-Paniceae); Prain, D., Ed.; Reeve & Co.: Ashford, UK, 1920; pp. 582–586. [Google Scholar]
- Fox, W.E.; Hatch, S.L. New combinations in Setaria (Poaceae: Paniceae). Sida 1999, 18, 1037–1047. [Google Scholar]
- Webster, R.D. Nomenclatural changes in Setaria and Paspalidium (Poaceae: Paniceae). Sida 1995, 16, 439–446. [Google Scholar]
- Veldkamp, J.F. Miscellaneous notes on Southeast Asian Gramineae. IX. Setaria and Paspalidium. Blumea 1994, 39, 373–384. [Google Scholar]
- Kellogg, E.A.; Aliscioni, S.S.; Morrone, O.; Pensiero, J.; Zuloaga, F.O. A phylogeny of Setaria (Poaceae, Panicoideae, Paniceae) and related genera based on the chloroplast gene ndhF. Int. J. Plant Sci. 2009, 170, 117–131. [Google Scholar] [CrossRef]
- Soreng, R.J.; Peterson, P.M.; Romaschenko, K.; Davidse, G.; Zuloaga, F.O.; Judziewicz, E.J.; Filgueiras, T.S.; Davis, J.I.; Morrone, O. A worldwide phylogenetic classification of the Poaceae (Gramineae). J. Syst. Evol. 2015, 53, 117–137. [Google Scholar] [CrossRef]
- Aliscioni, S.S.; Ospina, J.C.; Gomiz, N.E. Morphology and leaf anatomy of Setaria s.l. (Poaceae: Panicoideae: Paniceae) and its taxonomic significance. Plant Syst. Evol. 2016, 302, 173–185. [Google Scholar] [CrossRef]
- Pilger, R. Acritochaete, eine neue Gramineen-Gattung aus Afrika. Bot. Jahrb. Syst. 1902, 32, 53–55. [Google Scholar]
- POWO (Plants of the World Online). Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 8 March 2022).
- Giussani, L.M.; Cota-Sánchez, J.H.; Zuloaga, F.O.; Kellogg, E.A. A molecular phylogeny of the grass subfamily Panicoideae (Poaceae) shows multiple origins of C4 photosynthesis. Amer. J. Bot. 2001, 88, 1993–2012. [Google Scholar] [CrossRef]
- Gómez-Martínez, R.; Culham, A. Phylogeny of the subfamily Panicoideae with emphasis on the tribe Paniceae: Evidence from the trnL-F cpDNA region. In Grasses: Systematics and Evolution; Jacobs, S.W.L., Everett, J.E., Eds.; Commonwealth Scientific and Industrial Research Organization (CSIRO) Publishing: Collingwood, Australia, 2000; pp. 136–140. [Google Scholar]
- Zuloaga, F.O.; Morrone, O.; Giussani, L.M. A cladistic analysis of the Paniceae: A preliminary approach. In Grasses: Systematics and Evolution; Jacobs, S.W.L., Everett, J.E., Eds.; Commonwealth Scientific and Industrial Research Organization (CSIRO) Publishing: Collingwood, Australia, 2000; pp. 123–135. [Google Scholar]
- Duvall, M.R.; Noll, J.D.; Minn, A.H. Phylogenetics of Paniceae (Poaceae). Amer. J. Bot. 2001, 88, 1988–1992. [Google Scholar] [CrossRef]
- Duvall, M.R.; Saar, D.E.; Grayburn, W.S.; Holbrook, G.P. Complex transitions between C3 and C4 photosynthesis during the evolution of Paniceae: A phylogenetic case study emphasizing the position of Steinchisma hians (Poaceae), a C3–C4 intermediate. Int. J. Plant Sci. 2003, 164, 949–958. [Google Scholar] [CrossRef]
- Aliscioni, S.S.; Giussani, L.M.; Zuloaga, F.O.; Kellogg, E.A. A molecular phylogeny of Panicum (Poaceae: Paniceae): Tests of monophyly and phylogenetic placement within the Panicoideae. Am. J. Bot. 2003, 90, 796–821. [Google Scholar] [CrossRef]
- Christin, P.A.; Salamin, N.; Kellogg, E.A.; Vicentini, A.; Besnard, G. Integrating phylogeny into studies of C4 variation in the grasses. Plant Physiol. 2009, 149, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, T.; Bocksberger, G.; Brüggemann, W.; Berberich, T. Phylogenetic relationship and molecular taxonomy of African grasses of the genus Panicum inferred from four chloroplast DNA-barcodes and nuclear gene sequences. J. Plant Res. 2013, 126, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Grande Allende, J.R. Novitates Agrostologicae, IV. Additional segregates from Panicum incertae sedis. Phytoneuron 2014, 22, 1–6. [Google Scholar]
- Zuloaga, F.O.; Salomón, L.; Scataglini, M.A. Phylogeny of sections Clavelligerae and Pectinatae of Panicum (Poaceae, Panicoideae, Paniceae): Establishment of the new subtribe Dichantheliinae and the genus Adenochloa. Plant Syst. Evol. 2014, 301, 1693–1711. [Google Scholar] [CrossRef]
- Sede, S.M.; Morrone, O.; Giussani, L.M.; Zuloaga, F.O. Phylogenetic studies in the Paniceae (Poaceae): A realignment of section Lorea of Panicum. Syst. Bot. 2008, 33, 284–300. [Google Scholar] [CrossRef]
- Morrone, O.; Zuloaga, F.O.; Davidse, G.; Filgueiras, T.S. Canastra, a new genus Paniceae (Poaceae, Panicoideae) segregated from Arthropogon. Novon 2001, 11, 429–436. [Google Scholar] [CrossRef]
- Zuloaga, F.O.; Aliscioni, S.S.; Scataglini, M.A. Cnidochloa: A new genus of grasses from Brazil (Poaceae: Panicoideae: Paniceae). Ann. Missouri Bot. Gard. 2020, 15, 183–194. [Google Scholar] [CrossRef]
- Soreng, R.J. Coleataenia Griseb. (1879): The correct name for Sorengia Zuloaga & Morrone (2010) (Poaceae: Paniceae). J. Bot. Res. Inst. Texas 2010, 4, 691–692. [Google Scholar]
- Morrone, O.; Scataglini, M.A.; Zuloaga, F.O. Cyphonanthus, a new genus segregated from Panicum (Poaceae: Panicoideae: Paniceae) based on morphological, anatomical, and molecular data. Taxon 2007, 56, 521–532. [Google Scholar] [CrossRef]
- Zuloaga, F.O.; Giussani, L.M.; Morrone, O. Hopia, a new monotypic genus segregated from Panicum (Poaceae). Taxon 2007, 56, 145–156. [Google Scholar]
- Scataglini, M.A.; Zuloaga, F.O. Morronea, A new genus segregated from Panicum (Paniceae, Poaceae) based on morphological and molecular data. Syst. Bot. 2013, 38, 1076–1086. [Google Scholar] [CrossRef]
- Sede, S.M.; Zuloaga, F.O.; Morrone, O. Phylogenetic studies in the Paniceae (Poaceae-Panicoideae): Ocellochloa, a new genus from the New World. Syst. Bot. 2009, 34, 684–692. [Google Scholar] [CrossRef]
- Morrone, O.; Denham, S.S.; Aliscioni, S.S.; Zuloaga, F.O. Parodiophyllochloa, a new genus segregated from Panicum (Paniceae, Poaceae) based on morphological and molecular data. Syst. Bot. 2008, 33, 66–76. [Google Scholar] [CrossRef]
- Acosta, J.M.; Scataglini, M.A.; Reinheimer, R.; Zuloaga, F.O. A phylogenetic study of subtribe Otachyriinae (Poaceae, Panicoideae, Paspaleae). Plant Syst. Evol. 2014, 300, 2155–2167. [Google Scholar] [CrossRef]
- Zuloaga, F.O.; Scataglini, M.A.; Morrone, O. A phylogenetic evaluation of Panicum sects. Agrostoidea, Megista, Prionita and Tenera (Panicoideae, Poaceae): Two new genera, Stephostachys and Sorengia. Taxon 2010, 59, 1535–1546. [Google Scholar] [CrossRef]
- Zuloaga, F.O.; Morrone, O.; Scataglini, M.A. Monograph of Trichanthecium (Poaceae, Paniceae). Syst. Bot. Monogr. 2011, 94, 87–90. [Google Scholar]
- Bess, E.C.; Doust, A.N.; Davidse, G.; Kellogg, E.A. Zuloagaea, a New Genus of Neotropical Grass within the “Bristle Clade” (Poaceae: Paniceae). Syst. Bot. 2006, 31, 656–670. [Google Scholar] [CrossRef]
- Brown, W.V. The Kranz syndrome and its subtypes in grass systematics. Mem. Torrey Bot. Club 1977, 23, 1–97. [Google Scholar]
- Osborne, C.P.; Salomaa, A.; Kluyver, T.A.; Visser, V.; Kellogg, E.A.; Morrone, O.; Vorontsova, M.S.; Clayton, W.D.; Simpson, D.A. A global database of C4 photosynthesis in grasses. New Phytol. 2014, 204, 441–446. [Google Scholar] [CrossRef]
- Simon, B.K. Studies in Australian grasses 6*. Alexfloydia, Cliffordiochloa and Dallwatsonia, three new Panicoid grass genera. Austrobaileya 1992, 3, 669–681. [Google Scholar]
- Veldkamp, J.F. Revision of Panicum and Whiteochloa in Malesia (Gramineae-Paniceae). Blumea 1996, 41, 181–216. [Google Scholar]
- Rana, T.S.; Ranade, S.A. The enigma of monotypic taxa and their taxonomic implications. Curr. Sci. 2009, 96, 219–229. [Google Scholar]
- Rach, J.; Desalle, R.; Sarkar, I.N.; Schierwater, B.; Hadrys, H. Character-based DNA barcoding allows discrimination of genera, species and populations in Odonata. Proc. Biol. Sci. 2008, 275, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Dawkins, K.L.; Furse, J.M.; Wild, C.H.; Hughes, J.M. A novel genus and cryptic species harboured within the monotypic freshwater crayfish genus Tenuibranchiurus Riek, 1951 (Decapoda: Parastacidae). PeerJ 2017, 5, e3310. [Google Scholar] [CrossRef]
- Davidse, G.; Pohl, R.W. Chromosome numbers and notes on some Central American grasses. Can. J. Bot. 1972, 50, 273–283. [Google Scholar] [CrossRef]
- Sharma, M.L.; Sharma, K. Cytological studies in the North Indian grasses. Cytologia 1979, 44, 861–872. [Google Scholar] [CrossRef]
- Bir, S.S.; Sahni, M. SOCGI plant chromosome number reports—IV. J. Cytol. Genet. 1986, 21, 152–154. [Google Scholar]
- Sinha, R.R.P.; Bhardwaj, A.K.; Singh, R.K. SOCGI plant chromosome number reports—IX. J. Cytol. Genet. 1990, 25, 140–143. [Google Scholar]
- Ahsan, S.M.N.; Vahidy, A.A.; Ali, S.I. Chromosome Numbers and Incidence of Polyploidy in Panicoideae (Poaceae) from Pakistan. Ann. Mo. Bot. Gard. 1994, 81, 775–783. [Google Scholar] [CrossRef]
- Bir, S.S.; Sahni, M. SOCGI plant chromosome number reports—I. J. Cytol. Genet. 1983, 18, 58–59. [Google Scholar]
- Burton, G.W. A cytological study of some species in tribe Paniceae. Amer. J. Bot. 1942, 29, 355–361. [Google Scholar] [CrossRef]
- Gould, F.W. Chromosome numbers of Texas grasses. Can. J. Bot. 1968, 46, 1315–1325. [Google Scholar] [CrossRef]
- Hamoud, M.A.; Haroun, S.A.; Macleod, R.D.; Richards, A.J. Cytological relationships of selected species of Panicum L. Biol. Plant 1994, 35, 37–45. [Google Scholar] [CrossRef]
- Bor, N.L. Grasses of Burma, Ceylon, India and Pakistan; Pergamon Press Ltd.: Oxford, UK, 1960; 767p. [Google Scholar]
- Davidse, G.; Pohl, R.W. New Taxa and Nomenclatural Combinations of Mesoamerican Grasses (Poaceae). Novon 1992, 2, 81–110. [Google Scholar] [CrossRef]
- Hitchcock, A.S.; Chase, A. The North American Species of Panicum; U.S. Government Printing Office: Washington, DC, USA, 1910; Volume 15, pp. 1–396.
- Pilger, R. Gramineae III. Unterfamilie Panicoideae. In Die Natürlichen Pflanzenfamilien; Engler, A., Prantl, K., Eds.; W Engelmann: Leipzig, Germany, 1940; Volume 14e, pp. 1–208. [Google Scholar]
- Vasey, G. New species of grasses. Bull. Torrey. Bot. Club. 1884, 11, 61–72. [Google Scholar] [CrossRef]
- Hitchcock, A.S. Pseudochaetochloa, a new genus of grass from Australia. J. Wash. Acad. Sci. 1924, 14, 491–492. [Google Scholar]
- Sauer, J.D. Revision of Stenotaphrum (Gramineae: Paniceae) with attention to its historical geography. Brittonia 1972, 24, 202–222. [Google Scholar] [CrossRef]
- Webster, R.D. The Australian Paniceae (Poaceae); J. Cramer: Berlin, Germany, 1987; 322p. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. Bot. Soc. Amer. 1987, 19, 11–15. [Google Scholar]
- Olmstead, R.G.; Sweere, J.A. Combining data in phylogenetic systematics: An empirical approach using three molecular data sets in the Solanaceae. Syst. Biol. 1994, 43, 467–481. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Molec. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Fitch, W.M. Toward defining the course of evolution: Minimal change for a specific tree topology. Syst. Zool. 1971, 20, 406–416. [Google Scholar] [CrossRef]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Molec. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Crandall, K.A. Phylogeny estimation and hypothesis testing using maximum likelihood. Annu. Rev. Ecol. Syst. 1997, 28, 437–466. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Larget, B.; Miller, R.E.; Ronquist, F. Potential applications and pitfalls of Bayesian inference of phylogeny. Syst. Biol. 2002, 51, 673–688. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Farris, J.S.; Nixon, K.C. TNT, a free program for phylogenetic analysis. Cladistics 2008, 24, 774–786. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 23 December 2010. [Google Scholar]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web-servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff, New York Botanical Garden’s Virtual Herbarium, Continuously Updated. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 8 March 2022).


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delfini, C.; Aliscioni, S.S.; Acosta, J.M.; Pensiero, J.F.; Zuloaga, F.O. An Update of the Cenchrinae (Poaceae, Panicoideae, Paniceae) and a New Genus for the Subtribe to Clarify the Dubious Position of a Species of Panicum L. Plants 2023, 12, 749. https://doi.org/10.3390/plants12040749
Delfini C, Aliscioni SS, Acosta JM, Pensiero JF, Zuloaga FO. An Update of the Cenchrinae (Poaceae, Panicoideae, Paniceae) and a New Genus for the Subtribe to Clarify the Dubious Position of a Species of Panicum L. Plants. 2023; 12(4):749. https://doi.org/10.3390/plants12040749
Chicago/Turabian StyleDelfini, Carolina, Sandra S. Aliscioni, Juan M. Acosta, José F. Pensiero, and Fernando O. Zuloaga. 2023. "An Update of the Cenchrinae (Poaceae, Panicoideae, Paniceae) and a New Genus for the Subtribe to Clarify the Dubious Position of a Species of Panicum L." Plants 12, no. 4: 749. https://doi.org/10.3390/plants12040749
APA StyleDelfini, C., Aliscioni, S. S., Acosta, J. M., Pensiero, J. F., & Zuloaga, F. O. (2023). An Update of the Cenchrinae (Poaceae, Panicoideae, Paniceae) and a New Genus for the Subtribe to Clarify the Dubious Position of a Species of Panicum L. Plants, 12(4), 749. https://doi.org/10.3390/plants12040749