
Melatonin as a Possible Natural Anti-Viral Compound in Plant Biocontrol
 ,
,  , , and
, , and
Abstract
:1. Introduction
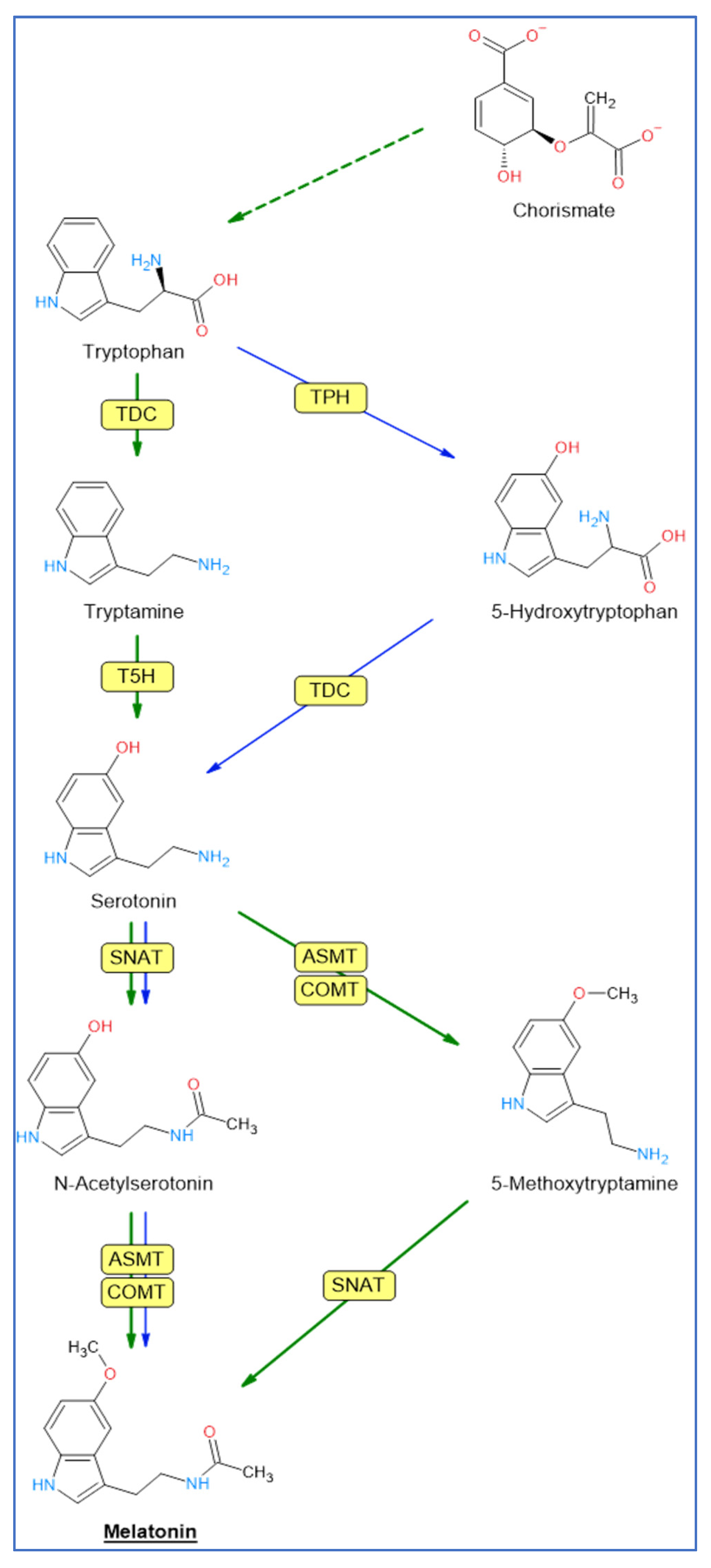
2. Biosynthetic Pathway of Melatonin
3. Physiological Roles of Phytomelatonin
4. Melatonin in Plant Disease Biocontrol
5. Melatonin-Integrated Plant Biocontrol Model
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wright, M.L. Melatonin, Diel Rhythms, and Metamorphosis in Anuran Amphibians. Gen. Comp. Endocrinol. 2002, 126, 251–254. [Google Scholar] [CrossRef]
- Zeman, M.; Herichová, I. Circadian Melatonin Production Develops Faster in Birds than in Mammals. Gen. Comp. Endocrinol. 2011, 172, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Claustrat, B.; Leston, J. Melatonin: Physiological Effects in Humans. Neurochirurgie 2015, 61, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Yu, Y.; Shen, Y.; Liu, Q.; Zhao, Z.; Sharma, R.; Reiter, R.J. Melatonin Synthesis and Function: Evolutionary History in Animals and Plants. Front. Endocrinol. 2019, 10, 249. [Google Scholar] [CrossRef]
- Saha, S.; Singh, K.M.; Gupta, B.B.P. Melatonin Synthesis and Clock Gene Regulation in the Pineal Organ of Teleost Fish Compared to Mammals: Similarities and Differences. Gen. Comp. Endocrinol. 2019, 279, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singh, R.; Tripathi, M.K. Photoperiodic Manipulation Modulates the Innate and Cell Mediated Immune Functions in the Fresh Water Snake, Natrix Piscator. Sci. Rep. 2020, 10, 14722. [Google Scholar] [CrossRef]
- Cheng, G.; Ma, T.; Deng, Z.; Gutiérrez-Gamboa, G.; Ge, Q.; Xu, P.; Zhang, Q.; Zhang, J.; Meng, J.; Reiter, R.J.; et al. Plant-Derived Melatonin from Food: A Gift of Nature. Food Funct. 2021, 12, 2829–2849. [Google Scholar] [CrossRef]
- Tilden, A.R.; Becker, M.A.; Amma, L.L.; Arciniega, J.; McGaw, A.K. Melatonin Production in an Aerobic Photosynthetic Bacterium: An Evolutionarily Early Association with Darkness. J. Pineal Res. 1997, 22, 102–106. [Google Scholar] [CrossRef]
- Rodriguez-Naranjo, M.I.; Torija, M.J.; Mas, A.; Cantos-Villar, E.; Garcia-Parrilla, M.d.C. Production of Melatonin by Saccharomyces Strains under Growth and Fermentation Conditions. J. Pineal Res. 2012, 53, 219–224. [Google Scholar] [CrossRef]
- Hardeland, R. Melatonin in the Evolution of Plants and Other Phototrophs. Melatonin Res. 2019, 2, 10–36. [Google Scholar] [CrossRef]
- Que, Z.; Ma, T.; Shang, Y.; Ge, Q.; Zhang, Q.; Xu, P.; Zhang, J.; Francoise, U.; Liu, X.; Sun, X. Microorganisms: Producers of Melatonin in Fermented Foods and Beverages. J. Agric. Food Chem. 2020, 68, 4799–4811. [Google Scholar] [CrossRef]
- Rehman, R.S.; Hussain, M.; Ali, M.; Zafar, S.A.; Pasha, A.N.; Bashir, H.; Ashraf, N.A.; Javed, A.; Shah, W.A. A Comprehensive Review on Melatonin Compound and Its Functions in Different Fungi and Plants. Int. J. Pathog. Res. 2022, 10, 9–21. [Google Scholar] [CrossRef]
- Arnao, M.B. Phytomelatonin: Discovery, Content, and Role in Plants. Adv. Bot. 2014, 2014, e815769. [Google Scholar] [CrossRef]
- Lerner, A.B.; Case, J.D.; Takahashi, Y.; Lee, T.H.; Mori, W. Isolation of Melatonin, the Pineal Gland Factor That Lightens Melanocytes 1. J. Am. Chem. Soc. 1958, 80, 2587. [Google Scholar] [CrossRef]
- Lerner, A.B.; Case, J.D.; Heinzelman, R.V. Structure of Melatonin. J. Am. Chem. Soc. 1959, 81, 6084–6085. [Google Scholar] [CrossRef]
- Kolar, J.; Machackova, I.; Illnerova, H.; Prinsen, E.; van Dongen, W.; van Onckelen, H. Melatonin in Higher Plant Determined by Radioimmunoassay and Liquid Chromatography-Mass Spectrometry. Biol. Rhythm Res. 1995, 26, 406–409. [Google Scholar]
- Dubbels, R.; Reiter, R.J.; Klenke, E.; Goebel, A.; Schnakenberg, E.; Ehlers, C.; Schiwara, H.W.; Schloot, W. Melatonin in Edible Plants Identified by Radioimmunoassay and by High Performance Liquid Chromatography-Mass Spectrometry. J. Pineal Res. 1995, 18, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Hattori, A.; Migitaka, H.; Iigo, M.; Itoh, M.; Yamamoto, K.; Ohtani-Kaneko, R.; Hara, M.; Suzuki, T.; Reiter, R.J. Identification of Melatonin in Plants and Its Effects on Plasma Melatonin Levels and Binding to Melatonin Receptors in Vertebrates. Biochem. Mol. Biol. Int. 1995, 35, 627–634. [Google Scholar]
- Xie, Z.; Chen, F.; Li, W.A.; Geng, X.; Li, C.; Meng, X.; Feng, Y.; Liu, W.; Yu, F. A Review of Sleep Disorders and Melatonin. Neurol. Res. 2017, 39, 559–565. [Google Scholar] [CrossRef]
- Vadnie, C.A.; McClung, C.A. Circadian Rhythm Disturbances in Mood Disorders: Insights into the Role of the Suprachiasmatic Nucleus. Neural Plast. 2017, 2017, e1504507. [Google Scholar] [CrossRef]
- Majidinia, M.; Reiter, R.J.; Shakouri, S.K.; Yousefi, B. The Role of Melatonin, a Multitasking Molecule, in Retarding the Processes of Ageing. Ageing Res. Rev. 2018, 47, 198–213. [Google Scholar] [CrossRef]
- Blume, C.; Angerer, M.; Raml, M.; del Giudice, R.; Santhi, N.; Pichler, G.; Kunz, A.B.; Scarpatetti, M.; Trinka, E.; Schabus, M. Healthier Rhythm, Healthier Brain? Integrity of Circadian Melatonin and Temperature Rhythms Relates to the Clinical State of Brain-Injured Patients. Eur. J. Neurol. 2019, 26, 1051–1059. [Google Scholar] [CrossRef] [Green Version]
- Socaciu, A.I.; Ionut, R.; Socaciu, M.A.; Ungur, A.P.; Bârsan, M.; Chiorean, A.; Socaciu, C.; Râjnoveanu, A.G. Melatonin, an Ubiquitous Metabolic Regulator: Functions, Mechanisms and Effects on Circadian Disruption and Degenerative Diseases. Rev. Endocr. Metab. Disord. 2020, 21, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Jan, J.E.; Reiter, R.J.; Wasdell, M.B.; Bax, M. The Role of the Thalamus in Sleep, Pineal Melatonin Production, and Circadian Rhythm Sleep Disorders. J. Pineal Res. 2009, 46, 1–7. [Google Scholar] [CrossRef]
- Luo, C.; Yang, Q.; Liu, Y.; Zhou, S.; Jiang, J.; Reiter, R.J.; Bhattacharya, P.; Cui, Y.; Yang, H.; Ma, H.; et al. The Multiple Protective Roles and Molecular Mechanisms of Melatonin and Its Precursor N-Acetylserotonin in Targeting Brain Injury and Liver Damage and in Maintaining Bone Health. Free Radic. Biol. Med. 2019, 130, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Poeggeler, B.; Tan, D.X.; Chen, L.; Manchester, L.; Guerrero, J. Antioxidant Capacity of Melatonin. A Novel Action Not Requiring a Receptor. Neuroendocrinol. Lett. 1993, 15, 103–116. [Google Scholar]
- Tan, D.X.; Poeggeler, B.; Reiter, R.J.; Chen, L.D.; Chen, S.; Lucien, M.C.; Barlow-Walden, L.R. The Pineal Hormone Melatonin Inhibits DNA-Adduct Formation Induced by the Chemical Carcinogen Safrole in Vivo. Cancer Lett. 1993, 70, 65–71. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin and Reactive Oxygen and Nitrogen Species: A Model for the Plant Redox Network. Melatonin Res. 2019, 2, 152–168. [Google Scholar] [CrossRef]
- Srinivasan, V.; Pandi-Perumal, S.R.; Maestroni, G.J.M.; Esquifino, A.I.; Hardeland, R.; Cardinali, D.P. Role of Melatonin in Neurodegenerative Diseases. Neurotoxicol. Res. 2005, 7, 293–318. [Google Scholar] [CrossRef]
- Miller, S.C.; Pandi-Perumal, S.R.; Esquifino, A.I.; Cardinali, D.P.; Maestroni, G.J.M. The Role of Melatonin in Inmmuno-Enhancement: Potential Application in Cancer. Int. J. Exp. Pathol. 2006, 87, 81–87. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Hardeland, R. Inflammaging, Metabolic Syndrome and Melatonin: A Call for Treatment Studies. Neuroendocrinology 2017, 104, 382–397. [Google Scholar] [CrossRef] [PubMed]
- Su, S.C.; Hsieh, M.J.; Yang, W.E.; Chung, W.H.; Reiter, R.J.; Yang, S.F. Cancer Metastasis: Mechanisms of Inhibition by Melatonin. J. Pineal Res. 2017, 62, e12370. [Google Scholar] [CrossRef]
- Cardinali, D.P. Melatonin and Healthy Aging. In Vitamins and Hormones Hormones and Aging; Litwack, G., Ed.; Academic Press: Cambridge, MA, USA, 2021; Volume 115, pp. 67–88. ISBN 0083-6729. [Google Scholar]
- Malabadi, R.B.; Kolkar, K.P.; Meti, N.T.; Chalannavar, R.K. Melatonin: One Molecule One- Medicine for Many Diseases, Coronavirus (SARS-CoV-2) Disease (Covid19); Function in Plants. IJRSI 2021, 8, 155–181. [Google Scholar] [CrossRef]
- Reiter, R.J.; Sharma, R.; Rodriguez, C.; Martin, V.; Rosales-Corral, S.; Zuccari, D.A.P.d.C.; Chuffa, L.G.d.A. Part-Time Cancers and Role of Melatonin in Determining Their Metabolic Phenotype. Life Sci. 2021, 278, 119597. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.; Sharma, R.; Rosales-Corral, S.; Manucha, W.; Chuffa, L.G.; Zuccari, D.A.P.C. Melatonin and Pathological Cell Interactions: Mitochondrial Glucose Processing in Cancer Cells. Int. J. Mol. Sci. 2021, 22, 12494. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, E.S.; Kampmann, U.; Pedersen, M.G.B.; Christensen, L.-L.; Jessen, N.; Møller, N.; Støy, J. Three Months of Melatonin Treatment Reduces Insulin Sensitivity in Patients with Type 2 Diabetes—A Randomized Placebo-Controlled Crossover Trial. J. Pineal Res. 2022, 73, e12809. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, N.; Huang, X.; Zhu, Y.; Gao, S.; Liu, Z.; Cao, F.; Wang, Y. Melatonin Engineered Adipose-Derived Biomimetic Nanovesicles Regulate Mitochondrial Functions and Promote Myocardial Repair in Myocardial Infarction. Front. Cardiovasc. Med. 2022, 9, 789203. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, E.; Valero, N.; Chacin-Bonilla, L.; Medina-Leendertz, S. Melatonin and Viral Infections. J. Pineal Res. 2004, 36, 73–79. [Google Scholar] [CrossRef]
- Tan, D.-X.; Korkmaz, A.; Reiter, R.J.; Manchester, L.C. Ebola Virus Disease: Potential Use of Melatonin as a Treatment. J. Pineal Res. 2014, 57, 381–384. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Brown, G.M.; Pandi-Perumal, S.R. Can Melatonin Be a Potential “Silver Bullet” in Treating COVID-19 Patients? Diseases 2020, 8, 44. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, X.; Ni, L.; Di, X.; Ma, B.; Niu, S.; Liu, C.; Reiter, R.J. COVID-19: Melatonin as a Potential Adjuvant Treatment. Life Sci. 2020, 250, 117583. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: A New Plant Hormone and/or a Plant Master Regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Cano, A.; Hernández-Ruiz, J. Phytomelatonin: An Unexpected Molecule with Amazing Performances in Plants. J. Exp. Bot. 2022, 73, 5779–5800. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Functions of Melatonin in Plants: A Review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. The Multi-Regulatory Properties of Melatonin in Plants. In Neurotransmitters in Plants; Ramakrishna, A., Roshchina, V.V., Eds.; CRC Press: Boca Raton, FL, USA, 2018; p. 448. ISBN 978-0-203-71148-4. [Google Scholar]
- Arnao, M.B.; Hernández-Ruiz, J. Is Phytomelatonin a New Plant Hormone? Agronomy 2020, 10, 95. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin in Flowering, Fruit Set and Fruit Ripening. Plant Reprod. 2020, 33, 77–87. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Huang, Y.; Bie, Z.; Ahmad, W.; Reiter, R.J.; Niu, M.; Hameed, S. Melatonin: Current Status and Future Perspectives in Plant Science. Front. Plant Sci. 2016, 6, 1230. [Google Scholar] [CrossRef]
- Sharif, R.; Xie, C.; Zhang, H.; Arnao, M.; Ali, M.; Ali, Q.; Muhammad, I.; Shalmani, A.; Nawaz, M.; Chen, P.; et al. Melatonin and Its Effects on Plant Systems. Molecules 2018, 23, 2352. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J.; Cano, A.; Reiter, R.J. Melatonin and Carbohydrate Metabolism in Plant Cells. Plants 2021, 10, 1917. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Mukherjee, S.; Flores, F.B.; Arnao, M.B.; Luo, Z.; Corpas, F.J. Functions of Melatonin During Postharvest of Horticultural Crops. Plant Cell Physiol. 2021, 63, 1764–1786. [Google Scholar] [CrossRef] [PubMed]
- Giraldo-Acosta, M.; Cano, A.; Hernández-Ruiz, J.; Arnao, M.B. Melatonin as a Possible Natural Safener in Crops. Plants 2022, 11, 890. [Google Scholar] [CrossRef]
- Buttar, Z.A.; Wu, S.N.; Arnao, M.B.; Wang, C.; Ullah, I.; Wang, C. Melatonin Suppressed the Heat Stress-Induced Damage in Wheat Seedlings by Modulating the Antioxidant Machinery. Plants 2020, 9, 809. [Google Scholar] [CrossRef]
- Moustafa-Farag, M.; Elkelish, A.; Dafea, M.; Khan, M.; Arnao, M.B.; Abdelhamid, M.T.; El-Ezz, A.A.; Almoneafy, A.; Mahmoud, A.; Awad, M.; et al. Role of Melatonin in Plant Tolerance to Soil Stressors: Salinity, PH and Heavy Metals. Molecules 2020, 25, 5359. [Google Scholar] [CrossRef] [PubMed]
- Moustafa-Farag, M.; Mahmoud, A.; Arnao, M.B.; Sheteiwy, M.; Dafea, M.; Soltan, M.; Elkelish, A.; Hasanuzzaman, M.; Ai, S. Melatonin-Induced Water Stress Tolerance in Plants: Recent Advances. Antioxidants 2020, 9, 809. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin as a Regulatory Hub of Plant Hormone Levels and Action in Stress Situations. Plant Biol. 2021, 23, 7–19. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin Against Environmental Plant Stressors: A Review. Curr. Protein Pept. Sci. 2021, 22, 413–429. [Google Scholar] [CrossRef] [PubMed]
- Altaf, M.A.; Shahid, R.; Ren, M.X.; Mora-Poblete, F.; Arnao, M.B.; Naz, S.; Anwar, M.; Altaf, M.M.; Shahid, S.; Shakoor, A.; et al. Phytomelatonin: An Overview of the Importance and Mediating Functions of Melatonin against Environmental Stresses. Physiol. Plant. 2021, 172, 820–846. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Mostafa, S.; Lu, Z.; Jin, B. Melatonin-Mediated Abiotic Stress Tolerance in Plants. Front. Plant Sci. 2022, 13, 847175. [Google Scholar] [CrossRef]
- Zhao, C.; Nawaz, G.; Cao, Q.; Xu, T. Melatonin Is a Potential Target for Improving Horticultural Crop Resistance to Abiotic Stress. Sci. Hortic. 2022, 291, 110560. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J.; Cano, A. Role of Melatonin and Nitrogen Metabolism in Plants: Implications under Nitrogen-Excess or Nitrogen-Low. Int. J. Mol. Sci. 2022, 23, 15217. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.-K. Abiotic Stress Responses in Plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Menhas, S.; Yang, X.; Hayat, K.; Aftab, T.; Bundschuh, J.; Arnao, M.B.; Zhou, Y.; Zhou, P. Exogenous Melatonin Enhances Cd Tolerance and Phytoremediation Efficiency by Ameliorating Cd-Induced Stress in Oilseed Crops: A Review. J. Plant Growth Regul. 2022, 41, 922–935. [Google Scholar] [CrossRef]
- Bhardwaj, R.; Aghadm, M.; Arnao, M.B.; Brecht, J.; Fawole, O.; Pareek, S. Melatonin Alleviates Chilling Injury Symptom Development in Mango Fruit by Maintaining Intracellular Energy and Cell Wall and Membrane Stability. Front. Nutr. 2022, 9, 936932. [Google Scholar] [CrossRef]
- Yang, X.; Ren, J.; Li, J.; Lin, X.; Xia, X.; Yan, W.; Zhang, Y.; Deng, X.; Ke, Q. Meta-Analysis of the Effect of Melatonin Application on Abiotic Stress Tolerance in Plants. Plant Biotechnol. Rep. 2022. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Lal, M.K.; Naga, K.C.; Kumar, R.; Chourasia, K.N.; Subhash, S.; Kumar, D.; Sharma, S. Emerging Roles of Melatonin in Mitigating Abiotic and Biotic Stresses of Horticultural Crops. Sci. Hortic. 2020, 272, 109592. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Lal, M.K.; Kumar, R.; Mangal, V.; Altaf, M.A.; Sharma, S.; Singh, B.; Kumar, M. Insight into Melatonin-Mediated Response and Signaling in the Regulation of Plant Defense under Biotic Stress. Plant Mol. Biol. 2021, 109, 385–399. [Google Scholar] [CrossRef] [PubMed]
- Onaga, G.; Wydra, K. Advances in Plant Tolerance to Biotic Stresses. In Plant Genomics; Abdurakhmonov, I.Y., Ed.; InTech: St. Paul, MN, USA, 2016; ISBN 978-953-51-2455-9. [Google Scholar]
- Hadidi, A.; Barba, M.; Candresse, T.; Jelkmann, W. Virus and Virus-Like Diseases of Pome and Stone Fruits; APS Press/American Phytopathological Society: St. Paul, MN, USA, 2011; ISBN 978-0-89054-501-0. [Google Scholar]
- Zhao, L.; Chen, L.; Gu, P.; Zhan, X.; Zhang, Y.; Hou, C.; Wu, Z.; Wu, Y.-F.; Wang, Q.-C. Exogenous Application of Melatonin Improves Plant Resistance to Virus Infection. Plant Pathol. 2019, 68, 1287–1295. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: Synthesis from Tryptophan and Its Role in Higher Plant. In Amino Acids in Higher Plants; D’Mello, J.P.F., Ed.; CABI: Boston, MA, USA, 2015; pp. 390–435. [Google Scholar] [CrossRef]
- Tan, D.-X.; Reiter, R.J. An Evolutionary View of Melatonin Synthesis and Metabolism Related to Its Biological Functions in Plants. J. Exp. Bot. 2020, 71, 4677–4689. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, C.L.; Esteban-Zubero, E.; Zhou, Z.; Reiter, J.R. Melatonin as a Potent and Inducible Endogenous Antioxidant: Synthesis and Metabolism. Molecules 2015, 20, 18886–18906. [Google Scholar] [CrossRef]
- Back, K.; Tan, D.-X.; Reiter, R.J. Melatonin Biosynthesis in Plants: Multiple Pathways Catalyze Tryptophan to Melatonin in the Cytoplasm or Chloroplasts. J. Pineal Res. 2016, 61, 426–437. [Google Scholar] [CrossRef]
- Byeon, Y.; Lee, H.Y.; Lee, K.; Back, K. Caffeic Acid O-Methyltransferase Is Involved in the Synthesis of Melatonin by Methylating N-Acetylserotonin in Arabidopsis. J. Pineal Res. 2014, 57, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Back, K. Melatonin Metabolism, Signaling and Possible Roles in Plants. Plant J. 2021, 105, 376–391. [Google Scholar] [CrossRef]
- Tan, D.X.; Hardeland, R.; Back, K.; Manchester, L.C.; Latorre-Jimenez, M.A.; Reiter, R.J. On the Significance of an Alternate Pathway of Melatonin Synthesis via 5-Methoxytryptamine: Comparisons across Species. J. Pineal Res. 2016, 61, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Tripathi, D.K.; Deshmukh, R.; Pratap Singh, V.; Corpas, F.J. Revisiting the Role of ROS and RNS in Plants under Changing Environment. Environ. Exp. Bot. 2019, 161, 1–398. [Google Scholar] [CrossRef]
- Melchiorri, D.; Reiter, R.J.; Attia, A.M.; Hara, M.; Burgos, A.; Nistico, G. Potent Protective Effect of Melatonin on in Vivo Paraquat-Induced Oxidative Damage in Rats. Life Sci. 1995, 56, 83–89. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Qi, W.; Manchester, L.C.; Karbownik, M.; Calvo, J.R. Pharmacology and Physiology of Melatonin in the Reduction of Oxidative Stress in Vivo. Biol. Signals Recept. 2000, 9, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchester, L.C.; Terron, M.P.; Flores, L.J.; Reiter, R.J. One Molecule, Many Derivates: A Never-Ending Interaction of Melatonin with Reactive Oxygen and Nitrogen Species? J. Pineal Res. 2007, 42, 28–42. [Google Scholar] [CrossRef]
- Prasad, A.; Sedlářová, M.; Balukova, A.; Rác, M.; Pospíšil, P. Reactive Oxygen Species as a Response to Wounding: In Vivo Imaging in Arabidopsis Thaliana. Front. Plant Sci. 2020, 10, 1660. [Google Scholar] [CrossRef]
- Li, D.; Wei, J.; Peng, Z.; Ma, W.; Yang, Q.; Song, Z.; Sun, W.; Yang, W.; Yuan, L.; Xu, X.; et al. Daily Rhythms of Phytomelatonin Signaling Modulate Diurnal Stomatal Closure via Regulating Reactive Oxygen Species Dynamics in Arabidopsis. J. Pineal Res. 2020, 68, e12640. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernandez-Ruiz, J. Melatonin in Its Relationship to Plant Hormones. Ann. Bot. 2018, 121, 195–207. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Growth Activity, Rooting Capacity, and Tropism: Three Auxinic Precepts Fulfilled by Melatonin. Acta Physiol. Plant. 2017, 39, 127. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Protective Effect of Melatonin against Chlorophyll Degradation during the Senescence of Barley Leaves. J. Pineal Res. 2009, 46, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Caputo, G.A.; Wadl, P.A.; McCarty, L.; Adelberg, J.; Jennings, K.M.; Cutulle, M. In Vitro Safening of Bentazon by Melatonin in Sweetpotato (Ipomoea Batatas). Hortscience 2020, 55, 1406–1410. [Google Scholar] [CrossRef]
- Abu-Qare, A.W.; Duncan, H.J. Herbicide Safeners: Uses, Limitations, Metabolism, and Mechanisms of Action. Chemosphere 2002, 48, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Giraldo-Acosta, M.; Martínez-Andújar, C.; Martínez-Melgarejo, P.A.; Cano, A.; Hernández-Ruiz, J.; Arnao, M.B. Protective Effect (Safener) of Melatonin on Vigna Radiata L. Seedlings in the Presence of the Fungicide Copper Oxychloride. J. Plant Growth Regul. 2022. [Google Scholar] [CrossRef]
- Hernández-Ruiz, J.; Ruiz-Cano, D.; Giraldo-Acosta, M.; Cano, A.; Arnao, M.B. Melatonin in Brassicaceae: Role in Postharvest and Interesting Phytochemicals. Molecules 2022, 27, 1523. [Google Scholar] [CrossRef]
- Wei, D.; Yang, J.; Xiang, Y.; Meng, L.; Pan, Y.; Zhang, Z. Attenuation of Postharvest Browning in Rambutan Fruit by Melatonin Is Associated With Inhibition of Phenolics Oxidation and Reinforcement of Antioxidative Process. Front. Nutr. 2022, 9, 905006. [Google Scholar] [CrossRef]
- Zhu, L.; Hu, H.; Luo, S.; Wu, Z.; Li, P. Melatonin Delaying Senescence of Postharvest Broccoli by Regulating Respiratory Metabolism and Antioxidant Activity. Trans. Chin. Soc. Agric. Eng. 2018, 34, 300–308. [Google Scholar]
- Cano, A.; Giraldo-Acosta, M.; García-Sánchez, S.; Hernández-Ruiz, J.; Arnao, M.B. Effect of Melatonin in Broccoli Postharvest and Possible Melatonin Ingestion Level. Plants 2022, 11, 2000. [Google Scholar] [CrossRef]
- Moustafa-Farag, M.; Almoneafy, A.; Mahmoud, A.; Elkelish, A.; Arnao, M.B.; Li, L.; Ai, S. Melatonin and Its Protective Role against Biotic Stress Impacts on Plants. Biomolecules 2020, 10, 54. [Google Scholar] [CrossRef]
- Collinge, D.B.; Jørgensen, H.J.L.; Latz, M.A.C.; Manzotti, A.; Ntana, F.; Rojas, E.C.; Jensen, B. Searching for Novel Fungal Biological Control Agents for Plant Disease Control Among Endophytes. In Endophytes for a Growing World; Murphy, B.R., Doohan, F.M., Saunders, M.J., Hodkinson, T.R., Eds.; Cambridge University Press: Cambridge, UK, 2019; pp. 25–51. ISBN 978-1-108-47176-3. [Google Scholar]
- Lamichhane, J.R.; Dachbrodt-Saaydeh, S.; Kudsk, P.; Messéan, A. Toward a Reduced Reliance on Conventional Pesticides in European Agriculture. Plant Dis. 2016, 100, 10–24. [Google Scholar] [CrossRef]
- Geiger, F.; Bengtsson, J.; Berendse, F.; Weisser, W.W.; Emmerson, M.; Morales, M.B.; Ceryngier, P.; Liira, J.; Tscharntke, T.; Winqvist, C.; et al. Persistent Negative Effects of Pesticides on Biodiversity and Biological Control Potential on European Farmland. Basic Appl. Ecol. 2010, 11, 97–105. [Google Scholar] [CrossRef]
- Kim, K.-H.; Kabir, E.; Jahan, S.A. Exposure to Pesticides and the Associated Human Health Effects. Sci. Total Environ. 2017, 575, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Raymaekers, K.; Ponet, L.; Holtappels, D.; Berckmans, B.; Cammue, B.P.A. Screening for Novel Biocontrol Agents Applicable in Plant Disease Management—A Review. Biol. Control 2020, 144, 104240. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Smart, L.E.; Birch, A.N.E.; Blok, V.C.; MacKenzie, K.; Guerrieri, E.; Cascone, P.; Luna, E.; Ton, J. Prospects for Plant Defence Activators and Biocontrol in IPM—Concepts and Lessons Learnt so Far. Crop Prot. 2017, 97, 128–134. [Google Scholar] [CrossRef]
- Fravel, D.R. Commercialization and Implementation of Biocontrol. Annu. Rev. Phytopathol. 2005, 43, 337–359. [Google Scholar] [CrossRef]
- Vos, C.M.F.; De Cremer, K.; Cammue, B.P.A.; De Coninck, B. The Toolbox of Trichoderma Spp. in the Biocontrol of Botrytis Cinerea Disease. Mol. Plant Pathol. 2015, 16, 400–412. [Google Scholar] [CrossRef]
- Burketova, L.; Trda, L.; Ott, P.G.; Valentova, O. Bio-Based Resistance Inducers for Sustainable Plant Protection against Pathogens. Biotechnol. Adv. 2015, 33, 994–1004. [Google Scholar] [CrossRef]
- Yin, L.; Wang, P.; Li, M.; Ke, X.; Li, C.; Liang, D.; Wu, S.; Ma, X.; Li, C.; Zou, Y.; et al. Exogenous Melatonin Improves Malus Resistance to Marssonina Apple Blotch. J. Pineal Res. 2013, 54, 426–434. [Google Scholar] [CrossRef]
- Wei, Y.; Hu, W.; Wang, Q.; Zeng, H.; Li, X.; Yan, Y.; Reiter, R.J.; He, C.; Shi, H. Identification, Transcriptional and Functional Analysis of Heat-Shock Protein 90s in Banana (Musa Acuminata L.) Highlight Their Novel Role in Melatonin-Mediated Plant Response to Fusarium Wilt. J. Pineal Res. 2017, 62, e12367. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, X.; Reiter, R.J.; Feng, S.; Wang, Y.; Liu, S.; Jin, L.; Li, Z.; Datla, R.; Ren, M. Melatonin Attenuates Potato Late Blight by Disrupting Cell Growth, Stress Tolerance, Fungicide Susceptibility and Homeostasis of Gene Expression in Phytophthora Infestans. Front. Plant Sci. 2017, 8, 1993. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Fard, J.R. Melatonin Treatment Attenuates Postharvest Decay and Maintains Nutritional Quality of Strawberry Fruits (Fragaria x Anannasa Cv. Selva) by Enhancing GABA Shunt Activity. Food Chem. 2017, 221, 1650–1657. [Google Scholar] [CrossRef] [PubMed]
- Mandal, M.K.; Suren, H.; Ward, B.; Boroujerdi, A.; Kousik, C. Differential Roles of Melatonin in Plant-Host Resistance and Pathogen Suppression in Cucurbits. J. Pineal Res. 2018, 65, e12505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Liu, S.; Zhang, J.; Reiter, R.J.; Wang, Y.; Qiu, D.; Luo, X.; Khalid, A.R.; Wang, H.; Feng, L.; et al. Synergistic Anti-Oomycete Effect of Melatonin with a Biofungicide against Oomycetic Black Shank Disease. J. Pineal Res. 2018, 65, e12492. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; He, Q.; Zhang, F.; Yu, J.; Li, C.; Zhao, T.; Zhang, Y.; Xie, Q.; Su, B.; Mei, L.; et al. Melatonin Enhances Cotton Immunity to Verticillium Wilt via Manipulating Lignin and Gossypol Biosynthesis. Plant J. 2019, 100, 784–800. [Google Scholar] [CrossRef]
- Liu, C.; Chen, L.; Zhao, R.; Li, R.; Zhang, S.; Yu, W.; Sheng, J.; Shen, L. Melatonin Induces Disease Resistance to Botrytis cinerea in Tomato Fruit by Activating Jasmonic Acid Signaling Pathway. J. Agric. Food Chem. 2019, 67, 6116–6124. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xu, Y.; Bi, Y.; Zhang, B.; Shen, S.; Jiang, T.; Zheng, X. Melatonin Treatment Inhibits Gray Mold and Induces Disease Resistance in Cherry Tomato Fruit during Postharvest. Postharvest Biol. Technol. 2019, 157, 110962. [Google Scholar] [CrossRef]
- Li, T.; Wu, Q.; Zhu, H.; Zhou, Y.; Jiang, Y.; Gao, H.; Yun, Z. Comparative Transcriptomic and Metabolic Analysis Reveals the Effect of Melatonin on Delaying Anthracnose Incidence upon Postharvest Banana Fruit Peel. BMC Plant Biol. 2019, 19, 289. [Google Scholar] [CrossRef]
- Lin, Y.; Fan, L.; Xia, X.; Wang, Z.; Yin, Y.; Cheng, Y.; Li, Z. Melatonin Decreases Resistance to Postharvest Green Mold on Citrus Fruit by Scavenging Defense-Related Reactive Oxygen Species. Postharvest Biol. Technol. 2019, 153, 21–30. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, Z.; Lan, G.; Jiao, C.; Sun, Y. Effect of Exogenous Melatonin on Resistance of Cucumber to Downy Mildew. Sci. Hortic. 2019, 255, 231–241. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Mao, Q.; Yan, Y.; Wu, M.; Wang, Y.; Ren, J.; Guo, P.; Liu, A.; Chen, S. Role of Melatonin in Arbuscular Mycorrhizal Fungi-Induced Resistance to Fusarium Wilt in Cucumber. Phytopathology 2020, 110, 999–1009. [Google Scholar] [CrossRef]
- Ali, M.; Lamin-Samu, A.; Muhammad, I.; Farghal, M.; Khattak, M.; Jan, I.; Ul Haq, S.; Khan, A.; Gong, Z.H.; Lu, G. Melatonin Mitigates the Infection of Colletotrichum gloeosporioides via Modulation of the Chitinase Gene and Antioxidant Activity in Capsicum annuum L. Antioxidants 2020, 10, 7. [Google Scholar] [CrossRef]
- Teng, Z.; Yu, Y.; Zhu, Z.; Hong, S.-B.; Yang, B.; Zang, Y. Melatonin Elevated Sclerotinia Sclerotiorum Resistance via Modulation of ATP and Glucosinolate Biosynthesis in Brassica rapa ssp. Pekinensis. J. Proteom. 2021, 243, 104264. [Google Scholar] [CrossRef] [PubMed]
- Jannatizadeh, A.; Aminian-Dehkordi, R.; Razavi, F. Effect of Exogenous Melatonin Treatment on Aspergillus Decay, Aflatoxin B1 Accumulation and Nutritional Quality of Fresh “Akbari” Pistachio Fruit. J. Food Process. Preserv. 2021, 45, e15518. [Google Scholar] [CrossRef]
- Kong, M.; Sheng, T.; Liang, J.; Ali, Q.; Gu, Q.; Wu, H.; Chen, J.; Liu, J.; Gao, X. Melatonin and Its Homologs Induce Immune Responses via Receptors TrP47363-TrP13076 in Nicotiana benthamiana. Front. Plant Sci. 2021, 12, 691835. [Google Scholar] [CrossRef]
- Huang, K.; Sui, Y.; Miao, C.; Chang, C.; Wang, L.; Cao, S.; Huang, X.; Li, W.; Zou, Y.; Sun, Z.; et al. Melatonin Enhances the Resistance of Ginger Rhizomes to Postharvest Fungal Decay. Postharvest Biol. Technol. 2021, 182, 111706. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, T.; Liu, G.; Hu, M.; Yun, Z.; Duan, X.; Cai, K.; Jiang, G. Inhibition of Downy Blight and Enhancement of Resistance in Litchi Fruit by Postharvest Application of Melatonin. Food Chem. 2021, 347, 129009. [Google Scholar] [CrossRef]
- Xie, X.; Han, Y.; Yuan, X.; Zhang, M.; Li, P.; Ding, A.; Wang, J.; Cheng, T.; Zhang, Q. Transcriptome Analysis Reveals That Exogenous Melatonin Confers Lilium Disease Resistance to Botrytis Elliptica. Front. Genet. 2022, 13, 892674. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Bi, R.; Cai, H.; Zhao, J.; Sun, P.; Xu, W.; Zhou, Y.; Yang, W.; Zheng, L.; Chen, X.-L.; et al. Melatonin Functions as a Broad-Spectrum Antifungal by Targeting a Conserved Pathogen Protein Kinase. J. Pineal Res. 2022, 74, e12839. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Byeon, Y.; Back, K. Melatonin as a Signal Molecule Triggering Defense Responses against Pathogen Attack in Arabidopsis and Tobacco. J. Pineal Res. 2014, 57, 262–268. [Google Scholar] [CrossRef]
- Lee, H.Y.; Byeon, Y.; Tan, D.-X.; Reiter, R.J.; Back, K. Arabidopsis Serotonin N-Acetyltransferase Knockout Mutant Plants Exhibit Decreased Melatonin and Salicylic Acid Levels Resulting in Susceptibility to an Avirulent Pathogen. J. Pineal Res. 2015, 58, 291–299. [Google Scholar] [CrossRef]
- Qian, Y.; Tan, D.-X.; Reiter, R.J.; Shi, H. Comparative Metabolomic Analysis Highlights the Involvement of Sugars and Glycerol in Melatonin-Mediated Innate Immunity against Bacterial Pathogen in Arabidopsis. Sci. Rep. 2015, 5, 15815. [Google Scholar] [CrossRef]
- Shi, H.; Chen, Y.; Tan, D.-X.; Reiter, R.J.; Chan, Z.; He, C. Melatonin Induces Nitric Oxide and the Potential Mechanisms Relate to Innate Immunity against Bacterial Pathogen Infection in Arabidopsis. J. Pineal Res. 2015, 59, 102–108. [Google Scholar] [CrossRef]
- Zhao, H.; Xu, L.; Su, T.; Jiang, Y.; Hu, L.; Ma, F. Melatonin Regulates Carbohydrate Metabolism and Defenses against Pseudomonas syringae pv. Tomato DC3000 Infection in Arabidopsis thaliana. J. Pineal Res. 2015, 59, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Back, K. Mitogen-Activated Protein Kinase Pathways Are Required for Melatonin-Mediated Defense Responses in Plants. J. Pineal Res. 2016, 60, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Back, K. Melatonin Is Required for H2O2—And NO-Mediated Defense Signaling through MAPKKK3 and OXI1 in Arabidopsis thaliana. J. Pineal Res. 2017, 62, e12379. [Google Scholar] [CrossRef]
- Chen, X.; Sun, C.; Laborda, P.; He, Y.; Zhao, Y.; Li, C.; Liu, F. Melatonin Treatments Reduce the Pathogenicity and Inhibit the Growth of Xanthomonas oryzae pv. Oryzicola. Plant Pathol. 2019, 68, 288–296. [Google Scholar] [CrossRef]
- Chen, X.; Laborda, P.; Liu, F. Exogenous Melatonin Enhances Rice Plant Resistance Against Xanthomonas oryzae pv. Oryzae. Plant Dis. 2020, 104, 1701–1708. [Google Scholar] [CrossRef] [PubMed]
- Nehela, Y.; Killiny, N. Melatonin Is Involved in Citrus Response to the Pathogen Huanglongbing via Modulation of Phytohormonal Biosynthesis. Plant Physiol. 2020, 184, 2216–2239. [Google Scholar] [CrossRef]
- Zhu, G.; Sha, P.F.; Zhu, X.X.; Shi, X.; Shahriar, M.; Zhou, Y.D.; Wang, S.; Laborda, P. Application of Melatonin for the Control of Food-Borne Bacillus Species in Cherry Tomatoes. Postharvest Biol. Technol. 2021, 181, 111656. [Google Scholar] [CrossRef]
- Yang, Q.; Peng, Z.; Ma, W.; Zhang, S.; Hou, S.; Wei, J.; Dong, S.; Yu, X.; Song, Y.; Gao, W.; et al. Melatonin Functions in Priming of Stomatal Immunity in Panax Notoginseng and Arabidopsis Thaliana. Plant Physiol. 2021, 187, 2837–2851. [Google Scholar] [CrossRef]
- Liu, Q.; Atta, U.R.; Wang, R.; Liu, K.; Ma, X.; Weng, Q. Defense-Related Hormone Signaling Coordinately Controls the Role of Melatonin during Arabidopsis thaliana-Pseudomonas syringae Interaction. Eur. J. Plant Pathol. 2021, 160, 707–716. [Google Scholar] [CrossRef]
- Wei, Y.; Zhu, B.; Ma, G.; Shao, X.; Xie, H.; Cheng, X.; Zeng, H.; Shi, H. The Coordination of Melatonin and Anti-Bacterial Activity by EIL5 Underlies Ethylene-Induced Disease Resistance in Cassava. Plant J. 2022, 111, 683–697. [Google Scholar] [CrossRef]
- Li, L.; Du, C.; Wang, L.; Lai, M.; Fan, H. Exogenous Melatonin Improves the Resistance to Cucumber Bacterial Angular Leaf Spot Caused by Pseudomonas syringae pv. Lachrymans. Physiol. Plant. 2022, 174, e13724. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, M.-R.; Li, J.-W.; Feng, C.-H.; Cui, Z.-H.; Zhao, L.; Wang, Q.-C. Exogenous Application of Melatonin Improves Eradication of Apple Stem Grooving Virus from the Infected in Vitro Shoots by Shoot Tip Culture. Plant Pathol. 2019, 68, 997–1006. [Google Scholar] [CrossRef]
- Lu, R.; Liu, Z.; Shao, Y.; Sun, F.; Zhang, Y.; Cui, J.; Zhou, Y.; Shen, W.; Zhou, T. Melatonin Is Responsible for Rice Resistance to Rice Stripe Virus Infection through a Nitric Oxide-Dependent Pathway. Virol. J. 2019, 16, 141. [Google Scholar] [CrossRef] [PubMed]
- Sofy, A.R.; Sofy, M.R.; Hmed, A.A.; Dawoud, R.A.; Refaey, E.E.; Mohamed, H.I.; El-Dougdoug, N.K. Molecular Characterization of the Alfalfa Mosaic Virus Infecting Solanum melongena in Egypt and the Control of Its Deleterious Effects with Melatonin and Salicylic Acid. Plants 2021, 10, 459. [Google Scholar] [CrossRef]
- Balconi, C.; Stevanato, P.; Motto, M.; Biancardi, E. Breeding for Biotic Stress Resistance/Tolerance in Plants. In Crop Production for Agricultural Improvement; Ashraf, M., Öztürk, M., Ahmad, M.S.A., Aksoy, A., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 57–114. ISBN 978-94-007-4116-4. [Google Scholar]
- Sofy, A.R.; Sofy, M.R.; Hmed, A.A.; El-Dougdoug, N.K. Potential Effect of Plant Growth-Promoting Rhizobacteria (PGPR) on Enhancing Protection Against Viral Diseases. In Field Crops: Sustainable Management by PGPR; Sustainable Development and Biodiversity; Maheshwari, D.K., Dheeman, S., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 411–445. ISBN 978-3-030-30926-8. [Google Scholar]
- Wang, S.M.; Hou, X.L.; Ying, L.; Cao, X.W.; Zhang, S.; Wang, F. Effects of Turnip mosaic virus (TuMV) on endogenous hormones and transcriptional level of related genes in infected non-heading Chinese cabbage. J. Nanjing Agric. Univ. 2011, 5, 13–19. [Google Scholar]
- Iriti, M.; Faoro, F. Abscisic Acid Is Involved in Chitosan-Induced Resistance to Tobacco Necrosis Virus (TNV). Plant Physiol. Biochem. 2008, 46, 1106–1111. [Google Scholar] [CrossRef]
- Zhao, L.; Feng, C.; Wu, K.; Chen, W.; Chen, Y.; Hao, X.; Wu, Y. Advances and Prospects in Biogenic Substances against Plant Virus: A Review. Pestic. Biochem. Physiol. 2017, 135, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Dodds, P.N.; Rathjen, J.P. Plant Immunity: Towards an Integrated View of Plant–Pathogen Interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, F.; Tsuda, K. Understanding the Plant Immune System. Mol. Plant Microbe Interact. 2010, 23, 1531–1536. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal Modulation of Plant Immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef]
- Yang, Y.-X.; Ahammed, G.J.; Wu, C.; Fan, S.; Zhou, Y.-H. Crosstalk among Jasmonate, Salicylate and Ethylene Signaling Pathways in Plant Disease and Immune Responses. Curr. Protein Pept. Sci. 2015, 16, 450–461. [Google Scholar] [CrossRef]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone Crosstalk in Plant Disease and Defense: More than Just Jasmonate-Salicylate Antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Bari, R.; Jones, J.D.G. Role of Plant Hormones in Plant Defence Responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J. Contrasting Mechanisms of Defense against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Wang, S.; Fu, J. Insights into Auxin Signaling in Plant–Pathogen Interactions. Front. Plant Sci. 2011, 2, 74. [Google Scholar]
- Ton, J.; Flors, V.; Mauch-Mani, B. The Multifaceted Role of ABA in Disease Resistance. Trends Plant Sci. 2009, 14, 310–317. [Google Scholar] [CrossRef]
- Suarez-Fernandez, M.; Marhuenda-Egea, F.C.; Lopez-Moya, F.; Arnao, M.B.; Cabrera-Escribano, F.; Nueda, M.J.; Gunsé, B.; Lopez-Llorca, L.V. Chitosan Induces Plant Hormones and Defenses in Tomato Root Exudates. Front. Plant Sci. 2020, 11, 572087. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, J.; Burgess, P.; Rossi, S.; Huang, B. Interactive Effects of Melatonin and Cytokinin on Alleviating Drought-Induced Leaf Senescence in Creeping Bentgrass (Agrostis stolonifera). Environ. Exp. Bot. 2018, 145, 1–11. [Google Scholar] [CrossRef]
- Zeng, H.; Bai, Y.; Wei, Y.; Reiter, R.J.; Shi, H. Phytomelatonin as a Central Molecule in Plant Disease Resistance. J. Exp. Bot. 2022, 73, 5874–5885. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Type of Pathogen | Pathogen Name | Plant Name | Melatonin Dose (µM) | Comments | Ref. |
|---|---|---|---|---|---|
| Fungi | Diplocarpon mali | Malus prunifolia | 50, 100, 500 | Resistance increased and number of lesions reduced | [105] |
| Fungi | Fusarium oxysporum | Musa acuminata | 100 | Reprogramming of defense-related plant hormones and confers disease resistance | [106] |
| Fungi | Phytophthora infestans | Solanum tuberosum | 1000, 3000, 6000, 8000, 10,000 | Suppression of the virulence and control disease | [107] |
| Fungi | Botrytis cinerea and Rhizopus stolonifer | Fragaria ananassa | 1, 10, 100, 1000 | Fruits with lower decay | [108] |
| Fungi | Podosphaera xanthii and Phythophthora capsici | Citrullus lanatus | 1000 | Improved plant immunity and suppression pathogen growth | [109] |
| Fungi | Phytophthora nicotianae | Nicotiana benthamiana | 500, 1000, 2000, 3000, 5000 | Suppression of the virulence and control disease | [110] |
| Fungi | Verticillium dahliae | Gossypium hirsutum | 10, 25, 50, 100 | Improved disease resistance | [111] |
| Fungi | Botrytis cinerea | Solanum lycopersicum | 1, 25, 50, 100 | Positive role resistance, regulating JA | [112] |
| Fungi | Botrytis cinerea | Solanum lycopersicum | 50–500 | Improved disease resistance and inhibited gray mold development | [113] |
| Fungi | Colletotrichum musae | Musa acuminate | 10,000 | Delayed senescence and anthracnose incidence | [114] |
| Fungi | Penicillium digitatum | Citrus reticulata | 50 | Decreased disease resistance | [115] |
| Fungi | Pseudoperonospora cubensis | Cucumis sativus | 100 | Reduced index disease | [116] |
| Fungi | Fusarium oxysporum | Cucumis sativus | 100 | Melatonin and arbuscular mycorrhizal enhances resistance | [117] |
| Fungi | Colletotrichum gloeosporioides | Capsicum annuum L. and A. thaliana | 50, 100, 500 | Mitigated the infection | [118] |
| Fungi | Sclerotinia sclerotiorum | Brassica rapa ssp. Pekinensis | 50, 100 | Induce defense mechanisms | [119] |
| Fungi | Aspergillus flavus | Pistacia vera cv. Akbari | 1000 | Alleviated oxidative stress and fungal decayed, decreased mycotoxin | [120] |
| Fungi | Phytophthora nicotianae | Nicotiana benthamiana | 50 | Induced immune responses | [121] |
| Fungi | Fusarium oxysporum and Penicillium brevicompactum | Zingiber officinale Roscoe | 100 | Reduced postharvest decays | [122] |
| Fungi | Peronophythora litchii | Litchi chinensis | 250 | Improved disease resistance and restricted lesion expansion | [123] |
| Fungi | Botrytis elliptica | Lilium cultivar “Sorbonne” | 20, 200, 2000, 20,000 | Improved plant resistance by MAPK signaling cascade | [124] |
| Fungi | Magnaporthe oryzae | Oryza sativa and Hordeum vulgare | 5000, 10,000 | Reduced disease severity and inhibited pathogen growth | [125] |
| Bacteria | Pseudomonas syringae pv. tomato DC3000 | Arabidopsis thaliana and N. benthamiana | 1, 10 | Molecule signaling of defense and inhibition of pathogen propagation | [126] |
| Bacteria | Pseudomonas syringae pv. tomato DC3000 | A. thaliana | 1, 2 | Increased resistance against pathogen, SA dependent | [127] |
| Bacteria | Pseudomonas syringae pv. tomato DC3000 | A. thaliana | 20 | Involvement of sugars and glycerol in melatonin-mediated innate immunity | [128] |
| Bacteria | Pseudomonas syringae pv. tomato DC3000 | A. thaliana | 20 | Improvement of disease resistance by nitric oxide (NO) | [129] |
| Bacteria | Pseudomonas syringae pv. tomato DC3000 | A. thaliana | 50 | Improved plant resistance by cell-wall reinforcement | [130] |
| Bacteria | Pseudomonas syringae pv. tomato DC3000 | A. thaliana and N. benthamiana | 1 | Improved plant resistance by MAPK signaling cascade | [131] |
| Bacteria | Pseudomonas syringae pv. tomato DC3000 | A. thaliana | 1 | Improved plant resistance by MAPK signaling cascade | [132] |
| Bacteria | Xanthomonas oryzae pv. oryzae | Oryza sativa and N. benthamiana | 861 | Reduces pathogenicity and incidence disease | [133] |
| Bacteria | Xanthomonas oryzae pv. oryzae | Oryza sativa | 86 | Improved plant resistance by NO and activation of defense-related genes | [134] |
| Bacteria | Candidatus liberibacter and Diaphorina citri | Citrus sinensis | 50–1000 | Increased disease resistance by modulation hormonal pathways | [135] |
| Bacteria | Bacillus cereus, B. licheniformis and B. subtilis | Solanum lycopersicum var. Cerasiforme | 5–10,000 | Reduces pathogenicity and incidence disease by ethylene and activation of defense-related genes | [136] |
| Bacteria | Pseudomonas syringe pv. tomato (Pst) DC3000 | Panax notoginseng and A. thaliana | 10 | Reduces pathogenicity and incidence disease | [137] |
| Bacteria | Pseudomonas syringe pv. tomato (Pst) DC3000 | A. thaliana | 10, 50 | Alteration bacterial resistance in some defense-related hormone signaling | [138] |
| Bacteria | Xanthomonas axonopodis pv. Manihotis | Manihot esculenta Crantz | 50, 100, 150 | Increased disease resistance by ethylene | [139] |
| Bacteria | Pseudomonas syringae pv. Lachrymans | Cucumis sativus | 10,100, 1000, 10,000 | Reduces pathogenicity and incidence disease | [140] |
| Virus | Tobacco Mosaic Virus (TMV) | Nicotiana glutinosa and S. lycopersicum | 100, 200 | Improve plant resistance to infection via SA and NO | [71] |
| Virus | Apple Stem Grooving Virus (ASGV) | Malus domestica | 10, 15, 20 | Efficient eradication plant virus | [141] |
| Virus | Rice Stripe Virus (RSV) | Oryza sativa | 10 | Improve disease resistance | [142] |
| Virus | Alfalfa Mosaic Virus (AMV) | Solanum melongena | 100 | Promotion resistance infection | [143] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Ruiz, J.; Giraldo-Acosta, M.; El Mihyaoui, A.; Cano, A.; Arnao, M.B. Melatonin as a Possible Natural Anti-Viral Compound in Plant Biocontrol. Plants 2023, 12, 781. https://doi.org/10.3390/plants12040781
Hernández-Ruiz J, Giraldo-Acosta M, El Mihyaoui A, Cano A, Arnao MB. Melatonin as a Possible Natural Anti-Viral Compound in Plant Biocontrol. Plants. 2023; 12(4):781. https://doi.org/10.3390/plants12040781
Chicago/Turabian StyleHernández-Ruiz, Josefa, Manuela Giraldo-Acosta, Amina El Mihyaoui, Antonio Cano, and Marino B. Arnao. 2023. "Melatonin as a Possible Natural Anti-Viral Compound in Plant Biocontrol" Plants 12, no. 4: 781. https://doi.org/10.3390/plants12040781