Abstract
Glutathione S-transferases (GSTs) are a critical superfamily of multifunctional enzymes in plants. As a ligand or binding protein, GSTs regulate plant growth and development and detoxification. Foxtail millet (Setaria italica (L.) P. Beauv) could respond to abiotic stresses through a highly complex multi-gene regulatory network in which the GST family is also involved. However, GST genes have been scarcely studied in foxtail millet. Genome-wide identification and expression characteristics analysis of the foxtail millet GST gene family were conducted by biological information technology. The results showed that 73 GST genes (SiGSTs) were identified in the foxtail millet genome and were divided into seven classes. The chromosome localization results showed uneven distribution of GSTs on the seven chromosomes. There were 30 tandem duplication gene pairs belonging to 11 clusters. Only one pair of SiGSTU1 and SiGSTU23 were identified as fragment duplication genes. A total of ten conserved motifs were identified in the GST family of foxtail millet. The gene structure of SiGSTs is relatively conservative, but the number and length of exons of each gene are still different. The cis-acting elements in the promoter region of 73 SiGST genes showed that 94.5% of SiGST genes possessed defense and stress-responsive elements. The expression profiles of 37 SiGST genes covering 21 tissues suggested that most SiGST genes were expressed in multiple organs and were highly expressed in roots and leaves. By qPCR analysis, we found that 21 SiGST genes were responsive to abiotic stresses and abscisic acid (ABA). Taken together, this study provides a theoretical basis for identifying foxtail millet GST family information and improving their responses to different stresses.
1. Introduction
Glutathione S-transferases (GSTs), a superfamily of enzymes encoded by multiple genes and having multiple functions, are ubiquitous in animals, plants, and microorganisms. Glutathione binds to harmful heterologous substances or oxidation products through GSTs, thereby promoting the metabolism, compartmentalization, or elimination of such substances [1]. The classical reaction mode is that GSTs catalyze the binding of glutathione to various hydrophobic and electrophilic electronic groups to form soluble S-glutathionylated products [2]. Fourteen categories have been identified according to the amino acid sequence similarity, among which eight categories are extensive, including eight subclasses: tau (U type), phi (F type), lambda (L type), theta (T type), zeta (Z type), γ-subunit of translation elongation factor (EF1G), dehydroascorbate reductase (DHAR), and tetrachloro hydroquinone dehalogenase (TCHQD) [3,4,5]. Among these classes of GSTs, tau, phi, lambda, and TCHQD are endemic to plants, and the tau and phi classes are the most abundant GST types in plants [6,7]. Although the sequence homology of the GST gene is low (about 25%), it has been found, by studying the structure of a large number of GSTs proteins [8,9], that these proteins have highly conserved structural characteristics.
Since the first discovery of glutathione S-transferases in maize in the 1970s, GSTs have been identified as a multigene family [10], and GSTs have been found in many plants. Genome-wide analyses revealed that there are 55 GST genes in Arabidopsis thaliana [11,12], 79 in Oryza sativa [13,14], 330 in Triticum aestivum [15,16], 84 in Hordeum vulgare [17], 59 in Gossypium Raimondi [18], 141 in Brassica napus [19], and 52 in Malus domestica [20]. In addition, the GST genes were identified in these plants, laying the foundation for isolating new GST genes from other plants.
Many studies have shown that the plant GST gene family can regulate the adaptability of plants to various kinds of stresses through electrophilic substitution, detoxification, and peroxide scavenging [21,22,23,24]. The expression of this gene family is not just affected by drought [25], saline-alkali [26], low temperature [27], pathogen infection [28], herbicides [29], heavy metals [30], and other stresses. It is also subject to ABA [31], auxins (IAA), ethylene [32], salicylic acid (SA) [32], jasmonic acid (JA) [33] and other plant hormones.
The expression of TaGSTU39 was significantly up-regulated throughout the treatment period under drought and salt stress treatments. TaGSTU62 of wheat could be down-regulated by gibberellin (GA) and up-regulated by ABA [15]. In A. thaliana, overexpression of grape GSTF13 could increase resistance to drought, salt, and methyl viologen stresses [34]. Multiple photoreceptors regulated the expression of AtGSTU17 and various development of A. thaliana seeds, including hypocotyl elongation and anthocyanin accumulation [35]. MdGSTF12 [20] and MdGSTU12 [36] were strongly induced by aminolevulinic acid (ALA), and they play an essential role in ALA-induced anthocyanin accumulation in apples. GmGSTU10 was induced explicitly by soybean mosaic virus and might have a highly efficient catalytic role in soybean [37].
A growing world population challenges global food and nutrition security [38]. In order to find suitable staple foods to overcome these difficult situations, millets are one of the potential candidates [39]. Foxtail millet is the oldest cultivated crop in the world, including China, and it is also the characteristic food crop in arid and semi-arid areas in northern China. Foxtail millet, as the main cultivated crop in dry green farming, has the characteristics of small genome size, short life cycle, self-pollination, drought resistance, and is able to grow in low fertility conditions, making it a model plant for studying C4 cereal crops [10,40,41,42]. However, there is no relevant report on the type, quantity, structure, and function of GST family in foxtail millet. Therefore, this study used genome and bioinformatics methods to analyze the GST gene family of foxtail millet transcription factors. A total of 73 GST genes were identified and divided into seven classes. The protein physicochemical properties, chromosomal distribution, gene structure, conserved motifs, cis-acting element, and gene expression of members of the GST family in foxtail millet were analyzed. We also exhibited the syntenic correlation between foxtail millet and A. thaliana genes. In addition, we investigated expression profiling of GST genes in different tissues and detected the expression of GST genes after various biotic and abiotic stresses in foxtail millet. Our study suggested that GST genes may play a role in regulating development and responding to various biotic and abiotic stresses and to ABA. Overall, this study provided a comprehensive identification of foxtail millet GST family members and provided a theoretical basis for further research on the functional analysis, gene editing, and genetic engineering of GST genes in foxtail millet.
2. Results
2.1. Identification of the Foxtail Millet GST Proteins and Analysis of Phylogenetic Relationship
The GST protein sequences of A. thaliana were searched against the protein sequences of foxtail millet to identify the GST proteins in foxtail millet. A total of 73 SiGST proteins were identified from the foxtail millet genome, based on the conserved GST-N and GST-C domains from these proteins (Figure S1). These GSTs were divided into seven distinct classes according to their conserved domains: tau, lambda, zeta, phi, DHAR, TCHQD, and MGST, and theta is absent in foxtail millet (Figure 1; Table S1). Among SiGST proteins, there are 44 tau proteins and 18 phi proteins, accounting for the vast majority, just as there are more tau and phi proteins in most plant GST families [43]. Next is lambda, which contains six members. The DHAR class had two members, and the zeta, TCHQD, and MGST classes had the least number, all of which had only one member.
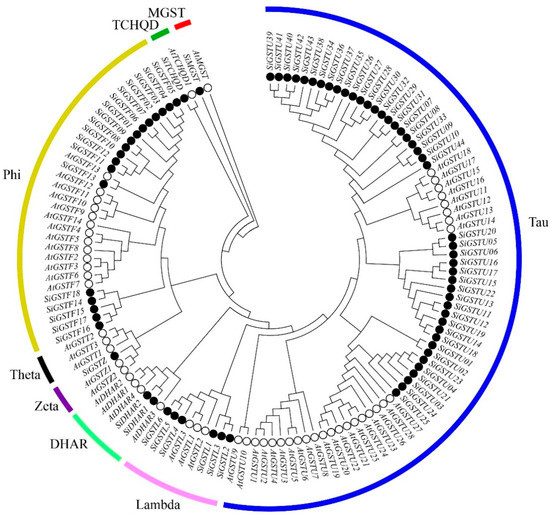
Figure 1.
Phylogenetic tree of GST proteins among foxtail millet and A. thaliana. The GST protein sequences of 73 foxtail millet and 55 A. thaliana are divided into eight classes. Different subfamilies are marked as different colors.
The analysis of physicochemical property showed that the sequence length of SiGST proteins varied from 182 (SiGSTF05) to 320 (SiGSTF18) amino acid residues, and the molecular weight (MW) was 20,177.45 (SiGSTF05)—36,490.24 (SiGSTF18) Da. The isoelectric point (pI) values were changed from 4.74 (SiGSTL1) to 9.05 (SiTCHQD). The instability index of proteins ranged from 23.76 (SiDHAR2) to 60.81 (SiGSTL5), of which 31 were instability proteins with an instability index greater than 40. According to the correlation principle of the grand average of hydropathicity (GRAVY), the amphiphilic protein is between −0.5 and 0.5, the positive is hydrophobic protein, and the negative is hydrophilic protein. SiGSTF14, SiGSTF18, and SiGSTU02 are hydrophilic proteins, and other proteins are amphotropic proteins, among which SiGSTU08 has a maximum value of 0.185 and SiGSTF18 has a minimum value of −1.085. The detailed data information for 73 SiGST protein sequences was tabulated (Table S2).
2.2. Chromosome Location and Gene Replication of SiGST Genes in Foxtail Millet
The chromosomal localization of 73 SiGST genes in foxtail millet revealed that SiGSTs were unevenly distributed on seven chromosomes. A high-density region containing GSTs was found on chromosomes III, V, and IX (Figure 2; Table S3). Among them, chromosome IX with 30 SiGST genes included the most members, followed by 22 on chromosome V. Eight SiGST genes were distributed on chromosome II, and nine SiGST genes were distributed on chromosome III. Only two SiGST genes were distributed on chromosome IV, and both chromosomes VII and VIII with one SiGST gene contained the least members. There is no SiGST gene on chromosomes I and VI.
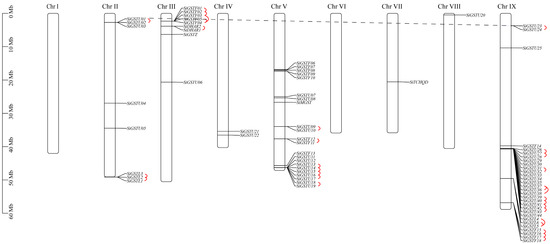
Figure 2.
Chromosome mapping of GST genes in foxtail millet. A total of 30 tandem duplication gene pairs belonging to 11 clusters were present (highlighted by full red lines). Only one pair of gene segmental duplication event was found (highlighted by dashed black line).
Segmental duplication and tandem duplication are considered to be two important factors for gene family expansion [44]. Among the 73 SiGST genes, a total of 30 gene pairs of 11 clusters were identified as the tandem duplication type (Figure 2). Among them, one pair of tandem duplication in DHAR class, four pairs in lambda class, eight pairs in phi class, and 17 pairs in tau class, indicating that a tandem duplication event contributed more to the expansion of the phi and tau classes. Only a pair of gene segmental duplication events (SiGSTU1 and SiGSTU23) occurred in all classes on seven chromosomes. Further analysis of the evolution of SiGST genes revealed that there was no syntenic relationships between SiGST gene and AtGST gene (Figure S2).
2.3. Conserved Motif and Gene Structure Analysis of SiGSTs
In order to better demonstrate the diversity and similarity of the SiGST motifs, the conserved structure of amino acids in the foxtail millet GST family was analyzed using the MEME database (Figure 3a,b). The results showed that ten conserved motifs were identified in the foxtail millet GST family, and each conserved motif length ranged from 11 to 50 amino acids (Figure S3). Tau and phi have more members, with motifs 1, 2, 3, 4, 5, and 6 found in 44 tau protein sequences, and motifs 4, 5, 7, and 8 found in 18 phi protein sequences. There were 72 SiGST proteins that contained motif 5, while 70 SiGST proteins contained motifs 2, 4, and 6. In lambda classes, there were coexistent motifs. In addition, some class members had completely identical motifs, such as DHAR, which contains motifs 1, 2, 4, 5, and 6.

Figure 3.
Gene structure and conserved motif of foxtail millet GSTs. (a) Neighbor-joining (NJ) phylogenetic tree based on SiGST protein sequences. (b) Conserved motif of SiGSTs. (c) Gene structure of SiGSTs.
Gain or loss of introns can alter gene structure and play a crucial role in the evolution of gene families [45]. Gene structure analysis showed that SiGSTU11, SiGSTU12, SiGSTU23, SiGSTU28, SiGSTU37, SiGSTF05, SiGSTF06, SiGSTF10, and SiGSTF12 did not contain upstream and downstream regulatory regions (UTR). SiGSTU02, SiGSTU03, SiGSTU10, SiGSTU25, SiGSTU29, SiGSTU30, SiGSTL5, and SiGSTF07 did not contain the upstream regulatory region outside. SiGSTU13 did not contain the downstream regulatory region outside. The other 55 members all have an upstream and downstream regulatory region (Figure 3c). At the same time, the gene sequence structure of the members of the GST family was analyzed, and the “exon-intron” structure diagram was obtained. Of these, most tau, phi, and TCHQD classes had one to three exons, while a phi member consisted of five exons. The DHAR and MGST classes exhibited six exons, while the zeta and lambda classes contained more exons than other classes (6–10 exons). The number of introns of the 73 GST genes in foxtail millet was less than ten, the most had nine, and the least had none. In addition, each member of the same subfamily has the same or similar gene structure. For example, most tau classes contained one intron; phi classes contained one to two introns, but SiGSTF06 contained four introns; TCHQD classes contained one intron; DHAR and MGST classes contained five introns; and zeta classes contained eight introns.
2.4. Prediction of Cis-Acting Elements in Promoter Region of GSTs Gene Family Members in Foxtail Millet
Using PlantCARE to analyze the 5′-upstream promoter (2.0 kb) region of 73 SiGSTs, we found that the cis-acting elements in the promoter region of 73 SiGST genes mainly included 20 kinds, including defense and stress responsive elements involved in salt, drought, low-temperature and anaerobic, light responsive element, hormone-responsive element associated with IAA, ABA, Methyl Jasmonate, GA, SA, and other elements related to growth regulation and circadian control, including meristem expression element, cell cycle regulation element, endosperm expression element, seed-specific regulation element, root specific element and MYB (MER), MYBHv1 binding site (Figure 4 and Figure S4; Table S4). The promoter regions of 67 SiGST genes presented in defense and stress response elements, and the cis-acting elements of 67 SiGST gene promoters contained hormone responsive elements. In addition, more than 40 cis-acting elements were identified in the promoters of the SiGSTF07, SiGSTF13, SiGSTU21, and SiTCHQD transcripts. However, only two cis-elements were identified in the promoters of SiGSTU39.
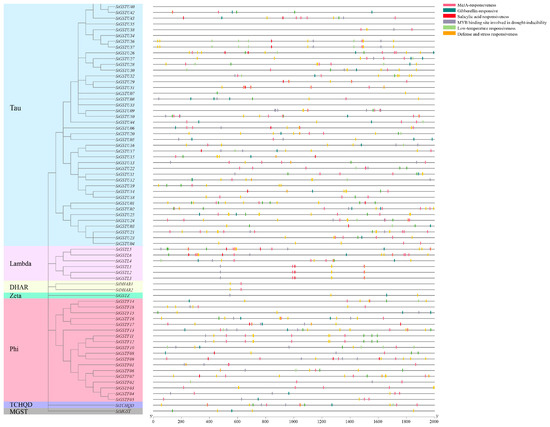
Figure 4.
Predicted cis-acting element in SiGST promoter of foxtail millet.
2.5. Relative Expression Patterns (FPKM Value) of GST Genes in Different Tissues
To explore the expression specificity of GST genes in foxtail millet at different tissues and developmental stages, we analyzed the expression profiles of 37 SiGST genes covering 21 tissues at different growth stages (Figure 5; Table S5). The results demonstrated partial differences in the expression of SiGST genes in different tissues of the same class. Additionally, most of the SiGST genes were expressed at higher levels in roots, which indicates that they may first play a role when the roots sense adversity. The transcription abundance of individual SiGST genes was low in all tissues and organs, such as SiGSTF07, which was weakly expressed in all tissues. However, some members had high transcript richness in all tissues and organs. SiGSTF11, SiGSTF12, and SiGSTF13 were strongly expressed in all tissues. There were tissue-specific expression characteristics in different SiGSTs members, such as SiGSTF01, SiGSTF03, SiGSTU05, and SiGSTU19, which are highly expressed in the leaves and roots of foxtail millet during grain fillinfigure s1 stage. The expression of SiGSTU06 in developing seeds and spikelets is relatively high, especially in panicle, which have the highest expression level. SiGSTL4 was slightly or even not expressed in different tissues. In addition, the genes were differentially expressed in different tissues and organs at the same developmental stage. SiGSTF11 and SiGSTF12 were highly expressed in roots during the filling stage and SiGSTF13 was highly expressed in neck-panicle-internodes during the filling stage. These results indicated that the relative expression pattern of SiGST genes in different tissues predicted its complex roles in foxtail millet growth and development.
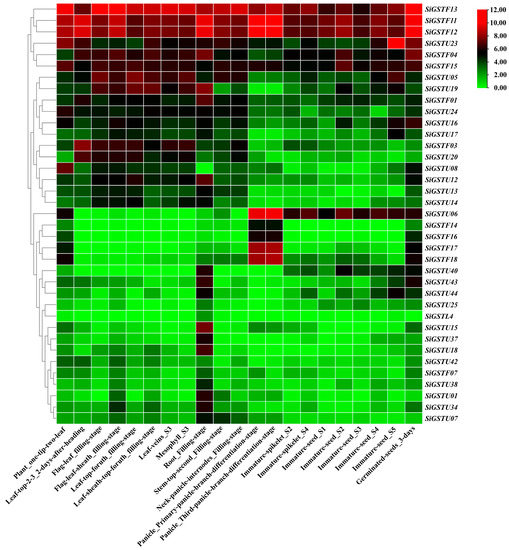
Figure 5.
Relative expression patterns (FPKM value) of GST genes involved in 21 tissues of foxtail millet. The FPKM values of SiGST genes in different tissues were obtained from the Setrari italica Multiomics database (http://foxtail-millet.biocloud.net/home, accessed on 8 June 2022). The color bar represents log2 expression levels (FPKM), with red indicating high gene expression levels and green indicating low gene expression levels.
2.6. Relative Expression Patterns of 21 SiGSTs under Abiotic Stresses and ABA Treatments in Foxtail Millet
To understand the responses of SiGST genes to ABA and other abiotic stresses, we selected 21 genes to analyze their expression in foxtail millet treated with osmotic (20% PEG 6000), salt (200 mM NaCl), cold stress (4 °C temperature), and 100 μM ABA, respectively (Figure 6). In general, SiGST gene expression did not show consistent characteristics under stress and hormone treatments. Under osmotic stress treatment, the expression of SiGSTF11, SiGSTF14, SiGSTU01, SiGSTU05, SiGSTU13, SiGSTU14, SiGSTU20, SiGSTU24, and SiGSTU42 were significantly up-regulated, and the expression level of SiGSTU1 was remarkably up-regulated. Its expression peaked at 12 h, which was 56.5 times that of 0 h. The expression of SiGSTF07, SiGSTU16, SiGSTU20, and SiGSTU23 peaked at 6 h; SiGSTU13 at 12 h; SiGSTF12, SiGSTF16, SiGSTU14, and SiGSTU17 at 24 h; and SiGSTF03 and SiGSTU44 at 48 h (Figure 6a).
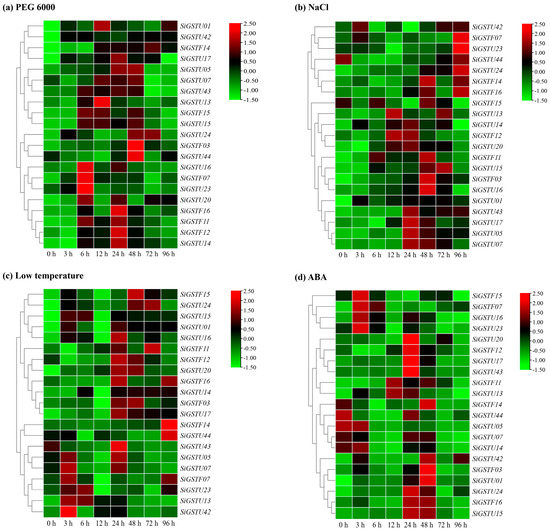
Figure 6.
Relative expression patterns of 21 SiGSTs in leaves of foxtail millet under abiotic stresses and ABA treatments. Relative expression patterns of 21 SiGST genes were analyzed with qPCR under osmotic (20% PEG 6000) (a), salt (200 mM NaCl) (b), cold stress (4 °C temperature) (c), and 100 μM ABA (d). The relative expression levels of each gene were calculated by the 2−∆∆Ct method. The heat map of SiGSTs expression condition was constructed by TBtools, expression value was standardized by Log2. The expression level of target gene at 0 h was used as control.
Under salt stress treatment, the SiGSTF03, SiGSTF11, SiGSTU01, SiGSTU15, SiGSTU24, and SiGSTU43 genes were induced more significantly, and the expression was higher than 0 h. The expression peak for SiGSTF12 and SiGSTU13 appeared at 12 h and up-regulated 7.2-fold and 6.7-fold compared to the control group. The expression levels of SiGSTU05, SiGSTU07, SiGSTU14, SiGSTU17, SiGSTU20, and SiGSTU43 genes were the highest at 24 h, and the SiGSTF03, SiGSTF11, SiGSTF14, SiGSTF15, and SiGSTU16 genes showed an expression peak at 48 h. The expression peak of SiGSTF07, SiGSTF16, SiGSTU23, and SiGSTU24 appeared at 96 h, and SiGSTU24 expression at 96 h, which is about 81 times that of 0 h (Figure 6b).
The expressions of SiGSTF11, SiGSTF12, SiGSTF16, and SiGSTU43 were inhibited and lower than those of the control group in the early stage of 4 °C temperature treatment. However, SiGSTU15, SiGSTU17, SiGSTU20, and SiGSTU24 expression levels were significantly higher than the control group during the whole 4 °C temperature treatment, and the SiGSTU20 induced more remarkably. The expression levels of SiGSTF12, SiGSTF16, SiGSTU05, SiGSTU16, SiGSTU17, SiGSTU20, and SiGSTU43 were significantly higher than the control group at 24 h. At 48 h, SiGSTF12 and SiGSTU20 were still significantly higher than in control group (Figure 6c).
Additionally, they could respond to ABA. For instance, the expression of SiGSTU5, SiGSTU07, and SiGSTU44 were down-regulated by ABA; and the expression of SiGSTF07, SiGSTU01, SiGSTU17, SiGSTU24, and SiGSTU42 could be up-regulated by ABA. The expression of SiGSTF12, SiGSTU15, SiGSTU17, SiGSTU20, SiGSTU24, and SiGSTU43 peaked at 24 h. The expression peak for SiGSTF03, SiGSTU01, and SiGSTU42 appeared at 48 h, up-regulated 16.1-fold, 19.2-fold, and 74.6-fold compared to the control group, respectively (Figure 6d).
3. Discussion
3.1. Identification and Analysis of GST Genes in Foxtail Millet
GSTs comprise a large and diverse gene family that is ubiquitous in a wide variety of organisms. Currently, the GST gene family has been confirmed to be involved in regulating the growth and development, stress resistance, and other processes of plants, such as A. thaliana [11,12] and rice [13], which has important biological significance. However, the identification and analysis of GST gene family in foxtail millet are still lacking. A total of 73 SiGST genes were identified from the foxtail millet genome, which were divided into seven categories. Among them, tau and phi contain the most SiGST family members, with 44 and 18 SiGST genes, respectively, which are the same as the findings of other plants such as soybean [46], rice [13], and pepper [47]. Tau and phi, known as plant-specific GSTs, had the most members and abundant content compared with other subfamilies [47]. A possible reason is that, compared with animals, in addition to detoxification, GST in higher plants also has unique functions such as transporting flavonoids, and regulating plant growth and development [30,48].
Gene structure is of great significance to the study of gene evolution [49]. The rearrangements and fusions of different chromosomal fragments may result in the gain or loss of an exon or intron, which play an important role in the diversification of multi-gene families [45]. We found that the number of GST exons is generally conserved within the same class in foxtail millet. GSTUs had one or two exons, GSTFs had two or three exons (except for SiGSTF06), DHAR and MGST had six exons, zeta had nine exons, and TCHQDs had two exons (Figure 3c). Similarly, the structural characteristics of GST genes were conserved in wheat, apple, and melon [15,20,50]. In addition, previous studies have shown that the earliest gene had the fewest number of introns, and as the replication progressed, the number of gene members continued to increase, and the number of introns gradually increased. It is pointed out that some of the origins of introns in the gene structure exist within themselves, and some are generated or increased with the insertion of transposons during gene replication [51,52]. In this study, through the analysis of the structure and conserved motifs of GST, it was found that most of the tau and phi classes contained one intron, the DHAR and MGST classes contained five introns, and the zeta class contained eight introns (Figure 3a,b). Therefore, according to the number of members of each subfamily and the number of introns in the gene, it is speculated that tau and phi may first appear in the GST family of foxtail millet. This was similar to the structure of GST family genes reported in wheat, apple, and radish [15,20,53].
The expansion of plant gene families is mainly achieved through different gene duplication methods (including fragment duplication, cascade duplication, gene transfer, and genome duplication, etc.) [18,54]. In addition, the expansion of GST family genes in plants was mainly completed by the tandem duplication of genes of tau and phi classes, making plant-specific tau and phi the two classes with the most members [6,47,55]. GST protein sequences of foxtail millet were compared, and the phylogenetic tree was constructed. It was divided into eight large groups (tau, phi, lambda, zeta, theta, DHAR, TCHQD, and MGST) (Figure 1). There were 28 A. thaliana genes in tau, accounting for 50.9% of the A. thaliana GST family. However, there were 44 members of the foxtail millet GST family in tau, accounting for 60.2%. This amplification in tau may be related to gene duplication during evolution. Indeed, we found that only one pair of segment duplication genes were in foxtail millet, and 30 pairs of genes were tandem duplications, and tandem duplications were more frequent than segment duplications (Figure 2). Similar phenomena have been observed in rice [13], apple [20], and Capsella rubella [56]. Gene duplication not only expands the content of the genome, but is also important for generating new gene functions, thereby enabling organisms to further adapt to complex environments [44]. Collinearity analysis indicated that strong selection pressure was subjected to during the evolution of GST gene family in foxtail millet.
3.2. The Expressions of SiGST Genes in Foxtail Millet
Tissue expression profiling analysis of SiGST genes indicated that most SiGST genes were expressed in multiple organs and highly expressed in roots and leaves, indicating that most SiGST genes may play a role in roots and leaves (Figure 5). We also found functional differentiation of GSTs in foxtail millet, and SiGSTs were expressed differently in different tissue parts of foxtail millet. For example, SiGSTF03 was highly expressed in parietal leaf at heading stage. SiGSTU06, SiGSTF17, and SiGSTF18 were highly expressed in panicle_primary-panicle-branch-differentiation-stage and panicle_third-panicle-branch-differentiation-stage. SiGSTL4, SiGSTU15, and SiGSTU18 were specifically expressed in the underground part of foxtail millet. In some tandem repeat pairs, the expression levels of the two genes differed, suggesting that the retention of gene duplication may be related to the process of tissue expression differences [57,58]. For example, SiGSTF15 was highly expressed in 21 different tissues, while SiGSTF07 was less expressed.
Plant growth and development are affected by various abiotic stresses, which activate the molecular mechanisms of plants to adapt to adverse conditions [59]. Drought, high salinity, and extreme temperatures limit the geographical distribution of plants because they cause dehydration and, eventually, cell death [53]. The GSTs promoter region of foxtail millet contains a large number of light response elements and stress response elements (low temperature, drought, stress defense, and stress), and the cis-acting elements in promoter regions of SiGST genes were found to be involved in the response to diverse biotic and abiotic stresses, as well as hormones (Figure 4 and Figure S4; Table S4). The 94.5% of SiGST genes possessed defense and stress responsive elements, and only six of them had no hormones responsive elements.
Previous studies have shown that the GST gene family is induced by abiotic stresses and hormones. For example, MdGSTF12 [20] and MdGSTU12 [36] were strongly induced by ALA. TaGSTU62 could be induced by osmotic stress, salt stress, and ABA [15]. However, OsGSTU4 could only be induced by salt stress [60]. The expression profiles of 21 SiGST genes under three abiotic stresses and one hormone (osmotic, salt, cold stress, and ABA) were analyzed by qPCR, showing that SiGST genes could be induced by abiotic stresses and ABA, and they might play a key role in abiotic stress response through corresponding hormone-dependent pathways (Figure 6). This study found that the SiGSTU17, SiGSTU24, and SiGSTF03 could be induced by abiotic stresses and ABA. Although these results indirectly proved that SiGST genes are involved in the response of Jingu 21 to stress, further analysis is still needed to confirm its ability to resist stresses.
4. Materials and Methods
4.1. GST Gene Identification, Phylogenetic Analysis and Physicochemical Properties of Foxtail Millet
To identify the SiGST proteins, the Setaria italica genome data were downloaded from the NCBI database (https://www.ncbi.nlm.nih.gov/genome/?term=Setaria+italica+, accessed on 14 August 2021). HMMER 3.0 software was used to identify SiGST genes, and the GST gene domain sequence (PF02798 and PF00043) was downloaded from the Pfam database (https://pfam.xfam.org/, accessed on 7 June 2022). The National Center for Biotechnology Information conserved domain database (www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi, accessed on 7 June 2022) was used to detect GST domains which were then mapped to the conserved domain. Neighbor-joining (NJ) phylogenetic tree was constructed using MEGA 7 software by the neighbor-joining method (bootstraps = 1000). The GST protein sequences of A. thaliana (55 numbers) were downloaded from The Arabidopsis Information Resource (TAIR, http://www.arabidopsis.org, accessed on 27 October 2020). Length of the protein sequence, protein molecular weight (MW), genomic position, isoelectric point (pI), instability index, and aliphatic index were predicted using ExPASy-ProParam (https://web.expasy.org/protparam/, accessed on 9 June 2022).
4.2. Distribution and Duplication Analysis of SiGSTs
TBtools was used to display the distribution of GST genes on Setaria italica chromosomes. Two or more GSTs separated by no more than three genes on the chromosome are called GST gene clusters. The gene sequences were aligned using BLASTp to determine the form of gene replication with an e-value of 1 × 10−1.
4.3. Gene Structure, Motif Compositions, and Gene Synteny of SiGST Genes
The gene structure map was produced, and an intron-exon map was compiled based on the S. italica genome annotation information (v 2.0). The MEME database (http://meme-suite.org/tools/meme, accessed on 9 June 2022) was used to conduct protein motif analysis. Chromosomal positions of SiGST genes were analyzed, and MCScanX was used to detect collinear regions between SiGST genes, as well as collinear blocks of SiGST genes with A. thaliana genes. The A. thaliana genome data were downloaded from the NCBI database. All above were visualized using TBtools [61].
4.4. Prediction of Cis-Acting Elements in the Promoter of SiGST Genes
Using the genome sequence of SiGSTs obtained in the phytozome database, the 2.0 kb DNA sequence upstream of SiGST genes was submitted to the PlantCARE database (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 17 June 2022) to predict cis-acting element. Promoter cis-acting regulatory element predictions were performed and visualized in TBtools.
4.5. Analysis of the Expression Pattern of SiGST Genes in Different Tissues
The FPKM values of SiGST genes, in the Multi-omics Database for Setrari italica (http://foxtail-millet.biocloud.net/home, accessed on 8 June 2022) in different tissues such as roots, stems, leaves, and flowers were extracted, and the TBtools software was used to draw gene expression heat maps for visualization.
4.6. Plant Materials and Treatments
The foxtail millet seeds of Jingu 21 were in a germination box with a temperature of 26 °C, a relative humidity of 65%, and a light cycle of 16 h/8 h. Two days later, the seedlings were transplanted into a plastic container with a modified half-strength Hoagland nutrient medium for further cultivation. For osmotic stress, salt stress, and ABA treatments, the 12-day-old seedlings were allowed to grow in 1/2 Hoagland solution containing 20% PEG 6000, 200 mM NaCl, and 100 μM ABA, respectively. For cold stress treatment, the 12-day-old seedlings were placed in a low temperature (4 °C) incubator to grow. The seedlings in 1/2 Hoagland solution without treatment at 26 ° C were regarded as controls. The leaves of the seedlings were sampled at 0 h, 3 h, 6 h, 12 h, 24 h, 48 h, 72 h, and 96 h after 20% PEG 6000, 200 mM NaCl, and 100 μM ABA treatment. All samples were immediately frozen in liquid nitrogen and stored in a −80 °C refrigerator.
4.7. RNA Isolation and qPCR
The acquisition of RNA from leaves was accomplished with TRIzol kit (Accurate Biology, Changsha, China), cDNA was synthesized using reverse transcription kit (Accurate Biology, Changsha, China), and real-time PCR was performed with SYBR Green dye method (Accurate Biology, Changsha, China). PCR primers were designed using Primer Premier 5 software (Table S6). The qRT-PCR reaction was performed in a Bio-Rad CFX system, a 20 µL reaction system containing 10 µL 2× SYBR Green Pro Taq HS premix, 2 µL cDNA, 0.4 µL each of forward primer and reverse primer, 7.2 µL RNase free water. The following cycling conditions were used: 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s, and 60 °C for 30 s. The SiActin (SETIT_026509mg) was used as internal standard, and the expression levels of each gene were calculated by the 2−ΔΔCt method. The heat map of SiGSTs expression condition was constructed by TBtools, expression value was standardized by Log2. Each independent experiment was repeated at least three times.
5. Conclusions
Through the appraisal of foxtail millet SiGST gene families, system evolution, and expression analysis, identified 73 millet SiGST genes, and the chromosomal location, the physicochemical properties, gene structure, and conservative structure domain analysis, this study forecast the SiGSTs promoter segment in response to ABA treatment, several abiotic stresses, and developmental stages. The expression profile spectrum analyses of SiGST genes showed that most of the SiGST genes were highly expressed in the roots and leaves. The qPCR analyses of 21 SiGST genes confirmed that SiGST genes were widely involved in stress and hormone responses such as drought, salt, low temperature, and ABA. These results provide a reference for further research on the gene function of foxtail millet molecular breeding and mining potential genetic resources.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12051138/s1, Figure S1: Conserved domain of foxtail millet GSTs. Figure S2: Collinearity analysis of GSTs in Setaria italica and Arabidopsis thaliana. Figure S3: The putative motifs of SiGST proteins in foxtail millet. Figure S4: Predicted cis-acting element in SiGST promoter of foxtail millet. Table S1: GST protein sequences of foxtail millet and A. thaliana used to construct the phylogenetic tree. Table S2: The detailed information of 73 SiGST genes, including gene name, accession number, number of amino acids, MW, chromosomal location, theoretical PI, instability index, aliphatic index, and grand average of hydropathicity. Table S3: The distributions of SiGST class members on foxtail millet chromosomes. Table S4: Putative cis-acting elements identified in the promoter regions of SiGSTs. Table S5: The expression levels (FPKM value) of SiGST genes involved in 21 tissues. Table S6: Primers used for qPCR.
Author Contributions
Conceptualization, X.L. and M.C.; Methodology, L.W., H.F. and X.L.; Software, L.W., H.F. and X.L.; Formal analysis, L.W., X.L. and M.C.; Investigation, L.W., H.F., J.Z., S.D. and H.F.; Resources, S.D., X.L. and H.F.; Data curation, L.W., J.W., J.Z., S.D, X.Y. and X.L.; Writing—original draft preparation, L.W.; Writing—Review & Editing, L.W., H.F., X.L. and M.C.; Visualization, X.L. and M.C.; Supervision, S.D., X.Y. and X.L.; Project Administration, J.W., S.D., X.Y., X.L. and M.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by State Key Laboratory of Sustainable Dryland Agriculture (in preparation), Shanxi Agricultural University (YJHZKF2104 and 202003-5), the Shanxi Province Basic Research Program Project (20210302124699), the Doctoral Research Start Project of Shanxi Agricultural University (2021BQ20), the Shanxi Province Doctoral Graduates and Postdoctoral Researchers to Work Award Fund Research Project (SXBYKY2021044), the China Agriculture Research System of MOF and MARA (CARS-06-14.5-A28), the National Natural Science Foundation of China (32272229 and 32200222), and the National Laboratory of Minor Crops Germplasm Innovation and Molecular Breeding (in preparation) (K462202040-32).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All GST protein sequences are provided in Table S1.
Acknowledgments
We thank Hongbo Fu (College of Biological and Agricultural Sciences, Honghe University) for his great contribution to the data analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nutricati, E.; Miceli, A.; Blando, F.; De Bellis, L. Characterization of two Arabidopsis thaliana glutathione S-transferases. Plant Cell Rep. 2006, 25, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Abdul Kayum, M.; Nath, U.K.; Park, J.I.; Biswas, M.K.; Choi, E.K.; Song, J.Y.; Kim, H.T.; Nou, I.S. Genome-Wide Identification, Characterization, and Expression Profiling of Glutathione S-Transferase (GST) Family in Pumpkin Reveals Likely Role in Cold-Stress Tolerance. Genes 2018, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- Lallement, P.A.; Brouwer, B.; Keech, O.; Hecker, A.; Rouhier, N. The still mysterious roles of cysteine-containing glutathione transferases in plants. Front. Pharmacol. 2014, 5, 192. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.P.; Edwards, R. Glutathione Transferases. In The Arabidopsis Book, Biologists; American Society of Plant Biologists: Rockville, MD, USA, 2010; Volume 8, pp. 1–15. [Google Scholar]
- Sasan, M.; Maryam, E.; Fateme, M.; Maryam, S.; Babak, S.; Hassan, M. Plant glutathione S-transferase classification, structure and evolution. Afr. J. Biochem. Res. 2011, 10, 8160–8165. [Google Scholar]
- Islam, S.; Rahman, I.A.; Islam, T.; Ghosh, A. Genome-wide identification and expression analysis of glutathione S-transferase gene family in tomato: Gaining an insight to their physiological and stress-specific roles. PLoS ONE 2017, 12, e0187504. [Google Scholar] [CrossRef]
- Dixon, D.P.; Lapthorn, A.; Edwards, R. Plant glutathione transferases. Genome Biol. 2002, 3, 1–10. [Google Scholar] [CrossRef]
- Brian McGonigle, S.J.K.; Sze-Mei, C.L.; Koeppe, M.K.; O’Keefe, D.P. A Genomics Approach to the Comprehensive Analysis of the Glutathione S-Transferase Gene Family in Soybean and Maize. Plant Physiol. 2000, 124, 1105–1120. [Google Scholar] [CrossRef]
- Thom, R.; Dixon, D.P.; Edwards, R.; Cole, D.J.; Lapthorn, A.J. The Structure of a Zeta Class Glutathione S-Transferase from Arabidopsis thaliana: Characterisation of a GST with Novel Active-site Architecture and a Putative Role in Tyrosine Catabolism. J. Mol. Biol. 2001, 308, 949–962. [Google Scholar] [CrossRef]
- Shimabukuro, R.H.; Swanson, H.R.; Walsh, W.C. Glutathione Conjugation. Plant Physiol. 1970, 46, 103–107. [Google Scholar] [CrossRef]
- Sappl, P.G.; Carroll, A.J.; Clifton, R.; Lister, R.; Whelan, J.; Harvey Millar, A.; Singh, K.B. The Arabidopsis glutathione transferase gene family displays complex stress regulation and co-silencing multiple genes results in altered metabolic sensitivity to oxidative stress. Plant J. 2009, 58, 53–68. [Google Scholar] [CrossRef]
- Wagner, U.; Edwards, R.; Dixon, D.P.; Mauch, F. Probing the Diversity of the Arabidopsis glutathione S-Transferase Gene Family. Plant Mol. Biol. 2002, 49, 515–532. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Ghanashyam, C.; Bhattacharjee, A. Comprehensive expression analysis suggests overlapping and specific roles of rice glutathione S-transferase genes during development and stress responses. BMC Genom. 2010, 11, 73. [Google Scholar] [CrossRef] [PubMed]
- Soranzo, N.; Sari Gorla, M.; Mizzi, L.; De Toma, G.; Frova, C. Organisation and structural evolution of the rice glutathione S-transferase gene family. Mol. Genet. Genom. 2004, 271, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Ma, J.; Zhang, Q.; Wu, C.; Zhao, H.; Wu, Y.; Yang, G.; He, G. Genome-wide identification and expression profiling of glutathione transferase gene family under multiple stresses and hormone treatments in wheat (Triticum aestivum L.). BMC Genom. 2019, 20, 986. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Xu, S.; Lyu, Z.; Wang, H.; Kong, L.; Sun, S. Comparative Analysis of the Glutathione S-Transferase Gene Family of Four Triticeae Species and Transcriptome Analysis of GST Genes in Common Wheat Responding to Salt Stress. Int. J. Genom. 2021, 2021, 6289174. [Google Scholar] [CrossRef]
- Rezaei, M.K.; Shobbar, Z.S.; Shahbazi, M.; Abedini, R.; Zare, S. Glutathione S-transferase (GST) family in barley: Identification of members, enzyme activity, and gene expression pattern. J. Plant Physiol. 2013, 170, 1277–1284. [Google Scholar] [CrossRef]
- Dong, Y.; Li, C.; Zhang, Y.; He, Q.; Daud, M.K.; Chen, J.; Zhu, S. Glutathione S-Transferase Gene Family in Gossypium raimondii and G. arboreum: Comparative Genomic Study and their Expression under Salt Stress. Front. Plant Sci. 2016, 7, 139. [Google Scholar] [CrossRef]
- Wei, L.; Zhu, Y.; Liu, R.; Zhang, A.; Zhu, M.; Xu, W.; Lin, A.; Lu, K.; Li, J. Genome wide identification and comparative analysis of glutathione transferases (GST) family genes in Brassica napus. Sci. Rep. 2019, 9, 9196. [Google Scholar] [CrossRef]
- Fang, X.; An, Y.; Zheng, J.; Shangguan, L.; Wang, L. Genome-wide identification and comparative analysis of GST gene family in apple (Malus domestica) and their expressions under ALA treatment. 3 Biotech 2020, 10, 307. [Google Scholar] [CrossRef]
- Enayati, A.A.; Ranson, H.; Hemingway, J. Insect glutathione transferases and insecticide resistance. Insect Mol. Biol. 2005, 14, 3–8. [Google Scholar] [CrossRef]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef] [PubMed]
- Townsend, D.M.; Tew, K.D. The role of glutathione-S-transferase in anti-cancer drug resistance. Oncogene 2003, 22, 7369–7375. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, G.M.D.; Foley, V.M.; Dowd, C.A. Structure, function and evolution of glutathione transferases: Implications for classification of non-mammalian members of an ancient enzyme superfamily. Biochem. J. 2001, 360, 1–16. [Google Scholar] [CrossRef] [PubMed]
- George, S.; Venkataraman, G.; Parida, A. A chloroplast-localized and auxin-induced glutathione S-transferase from phreatophyte Prosopis juliflora confer drought tolerance on tobacco. J. Plant Physiol. 2010, 167, 311–318. [Google Scholar] [CrossRef]
- Ji, W.; Zhu, Y.; Li, Y.; Yang, L.; Zhao, X.; Cai, H.; Bai, X. Over-expression of a glutathione S-transferase gene, GsGST, from wild soybean (Glycine soja) enhances drought and salt tolerance in transgenic tobacco. Biotechnol. Lett. 2010, 32, 1173–1179. [Google Scholar] [CrossRef]
- Huang, C.; Guo, T.; Zheng, S.C.; Feng, Q.L.; Liang, J.H.; Li, L. Increased cold tolerance in Arabidopsis thaliana transformed with Choristoneura fumiferana glutathione S-transferase gene. Biol. Plant. 2009, 53, 183–187. [Google Scholar] [CrossRef]
- Yan, Y.; Jia, H.; Wang, F.; Wang, C.; Liu, S.; Guo, X. Overexpression of GhWRKY27a reduces tolerance to drought stress and resistance to Rhizoctonia solani infection in transgenic Nicotiana benthamiana. Front. Physiol. 2015, 6, 00265. [Google Scholar] [CrossRef]
- Lo Cicero, L.; Madesis, P.; Tsaftaris, A.; Lo Piero, A.R. Tobacco plants over-expressing the sweet orange tau glutathione transferases (CsGSTUs) acquire tolerance to the diphenyl ether herbicide fluorodifen and to salt and drought stresses. Phytochemistry 2015, 116, 69–77. [Google Scholar] [CrossRef]
- Moons, A. Regulatory and Functional Interactions of Plant Growth Regulators and Plant Glutathione S-Transferases (GSTs). Vitam. Horm. 2005, 72, 155–202. [Google Scholar]
- Dixon, D.P.; Cole, D.J.; Edwards, R. Purification, regulation and cloning of a glutathione transferase (GST) from maize resembling the auxin-inducible type-III GSTs. Plant Mol. Biol. 1998, 36, 75–87. [Google Scholar] [CrossRef]
- Han, Q.; Chen, R.; Yang, Y.; Cui, X.; Ge, F.; Chen, C.; Liu, D. A glutathione S—transferase gene from Lilium regale Wilson confers transgenic tobacco resistance to Fusarium oxysporum. Sci. Hortic. 2016, 198, 370–378. [Google Scholar] [CrossRef]
- Kumar, S.; Asif, M.H.; Chakrabarty, D.; Tripathi, R.D.; Dubey, R.S.; Trivedi, P.K. Expression of a rice Lambda class of glutathione S-transferase, OsGSTL2, in Arabidopsis provides tolerance to heavy metal and other abiotic stresses. J. Hazard. Mater. 2013, 248–249, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zheng, A.Q.; Xing, X.J.; Chen, L.; Fu, X.Y.; Peng, R.H.; Tian, Y.S.; Yao, Q.H. Transgenic Arabidopsis Plants Expressing Grape Glutathione S-Transferase Gene (VvGSTF13) Show Enhanced Tolerance to Abiotic Stress. Biochemistry 2018, 83, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.W.; Liu, M.J.; Chen, I.C.; Huang, C.H.; Chao, L.Y.; Hsieh, H.L. A glutathione S-transferase regulated by light and hormones participates in the modulation of Arabidopsis seedling development. Plant Physiol. 2010, 154, 1646–1658. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.W.; Wang, C.K.; Huang, X.Y.; Hu, D.G. Genome-Wide Analysis of the Glutathione S-Transferase (GST) Genes and Functional Identification of MdGSTU12 Reveals the Involvement in the Regulation of Anthocyanin Accumulation in Apple. Genes 2021, 12, 1733. [Google Scholar] [CrossRef] [PubMed]
- Skopelitou, K.; Muleta, A.W.; Papageorgiou, A.C.; Chronopoulou, E.; Labrou, N.E. Catalytic features and crystal structure of a tau class glutathione transferase from Glycine max specifically upregulated in response to soybean mosaic virus infections. Biochim. Biophys. Acta 2015, 1854, 166–177. [Google Scholar] [CrossRef]
- Saxena, R.; Vanga, S.; Wang, J.; Orsat, V.; Raghavan, V. Millets for Food Security in the Context of Climate Change: A Review. Sustainability 2018, 10, 2228. [Google Scholar] [CrossRef]
- Chanwala, J.; Khadanga, B.; Jha, D.K.; Sandeep, I.S.; Dey, N. MYB Transcription Factor Family in Pearl Millet: Genome-Wide Identification, Evolutionary Progression and Expression Analysis under Abiotic Stress and Phytohormone Treatments. Plants 2023, 12, 355. [Google Scholar] [CrossRef]
- Muthamilarasan, M.; Prasad, M. Advances in Setaria genomics for genetic improvement of cereals and bioenergy grasses. Theor. Appl. Genet. 2015, 128, 1–14. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, H.; Li, X.; Shen, H.; Gao, J.; Hou, S.; Zhang, B.; Mayes, S.; Bennett, M.; Ma, J.; et al. A mini foxtail millet with an Arabidopsis-like life cycle as a C4 model system. Nat. Plants 2020, 6, 1167–1178. [Google Scholar] [CrossRef]
- Bennetzen, J.L.; Schmutz, J.; Wang, H.; Percifield, R.; Hawkins, J.; Pontaroli, A.C.; Estep, M.; Feng, L.; Vaughn, J.N.; Grimwood, J.; et al. Reference genome sequence of the model plant Setaria. Nat. Biotechnol. 2012, 30, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Edwards, R.; Dixon, D.P. Plant Glutathione Transferases. Meth. Enzymol. 2005, 401, 169–186. [Google Scholar]
- Freeling, M. Bias in plant gene content following different sorts of duplication: Tandem, whole-genome, segmental, or by transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef]
- Xu, G.; Guo, C.; Shan, H.; Kong, H. Divergence of duplicate genes in exon-intron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef]
- Liu, H.J.; Tang, Z.X.; Han, X.M.; Yang, Z.L.; Zhang, F.M.; Yang, H.L.; Liu, Y.J.; Zeng, Q.Y. Divergence in Enzymatic Activities in the Soybean GST Supergene Family Provides New Insight into the Evolutionary Dynamics of Whole-Genome Duplicates. Mol. Biol. Evol. 2015, 32, 2844–2859. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Sajib, S.D.; Jui, Z.S.; Arabia, S.; Islam, T.; Ghosh, A. Genome-wide identification of glutathione S-transferase gene family in pepper, its classification, and expression profiling under different anatomical and environmental conditions. Sci. Rep. 2019, 9, 9101. [Google Scholar] [CrossRef]
- Frova, C. Glutathione transferases in the genomics era: New insights and perspectives. Biochem. Eng. 2006, 23, 149–169. [Google Scholar] [CrossRef]
- Kang, L.; Teng, Y.; Cen, Q.; Fang, Y.; Tian, Q.; Zhang, X.; Wang, H.; Zhang, X.; Xue, D. Genome-Wide Identification of R2R3-MYB Transcription Factor and Expression Analysis under Abiotic Stress in Rice. Plants 2022, 11, 1928. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Z.; Wu, J.; Han, X.; Wang-Pruski, G.; Zhang, Z. Genome-wide identification, characterization, and expression analysis related to autotoxicity of the GST gene family in Cucumis melo L. Plant Physiol. Biochem. 2020, 155, 59–69. [Google Scholar] [CrossRef]
- Tijet, N.; Helvig, C.; Feyereisen, R. The cytochrome P450 gene superfamily in Drosophila melanogaster: Annotation, intron-exon organization and phylogeny. Gene 2001, 262, 189–198. [Google Scholar] [CrossRef]
- Jo, B.S.; Choi, S.S. Introns: The Functional Benefits of Introns in Genomes. Genom. Inform. 2015, 13, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Chen, B.; Lin, H.; Liu, Y.; Wei, Y.; Chen, F.; Li, W. Identification and characterization of the glutathione S-Transferase (GST) family in radish reveals a likely role in anthocyanin biosynthesis and heavy metal stress tolerance. Gene 2020, 743, 144484. [Google Scholar] [CrossRef] [PubMed]
- Mu, D.; Chen, W.; Shao, Y.; Wilson, I.W.; Zhao, H.; Luo, Z.; Lin, X.; He, J.; Zhang, Y.; Mo, C.; et al. Genome-Wide Identification and Expression Analysis of WRKY Transcription Factors in Siraitia siamensis. Plants 2023, 12, 288. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, Y.; Meng, Q.; Shi, F.; Liu, J.; Li, Y. Molecular cloning, identification of GSTs family in sunflower and their regulatory roles in biotic and abiotic stress. World J. Microbiol. Biotechnol. 2018, 34, 109. [Google Scholar] [CrossRef]
- He, G.; Guan, C.N.; Chen, Q.X.; Gou, X.J.; Liu, W.; Zeng, Q.Y.; Lan, T. Genome-Wide Analysis of the Glutathione S-Transferase Gene Family in Capsella rubella: Identification, Expression, and Biochemical Functions. Front. Plant Sci. 2016, 7, 1325. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Dopazo, J.; Huynen, M.A.; Gabaldon, T. Evidence for short-time divergence and long-time conservation of tissue-specific expression after gene duplication. Brief. Bioinform. 2011, 12, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Ganko, E.W.; Meyers, B.C.; Vision, T.J. Divergence in expression between duplicated genes in Arabidopsis. Mol. Biol. Evol. 2007, 24, 2298–3309. [Google Scholar] [CrossRef] [PubMed]
- Filyushin, M.A.; Kochieva, E.Z.; Shchennikova, A.V. ZmDREB2.9 Gene in Maize (Zea mays L.): Genome-Wide Identification, Characterization, Expression, and Stress Response. Plants 2022, 11, 3060. [Google Scholar] [CrossRef] [PubMed]
- Moons, A. Osgstu3andosgtu4, encoding tau class glutathioneS-transferases, are heavy metal- and hypoxic stress-induced and differentially salt stress-responsive in rice roots1. FEBS Lett. 2003, 553, 427–432. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).