
SNP Diversity and Genetic Structure of “Rogosija”, an Old Western Balkan Durum Wheat Collection
 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Results
2.1. Morphological Diversity in Western Balkan Durum Accessions
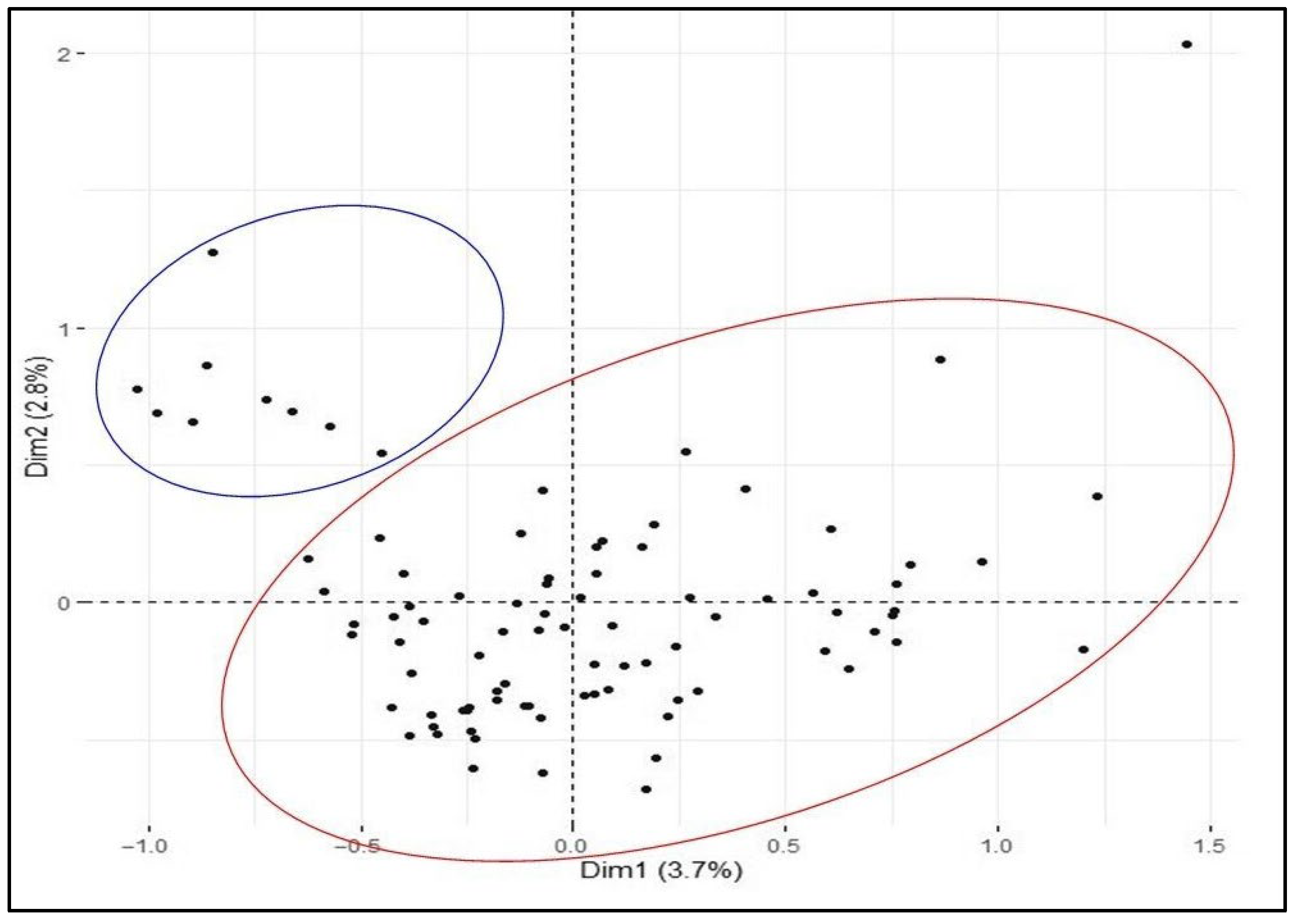
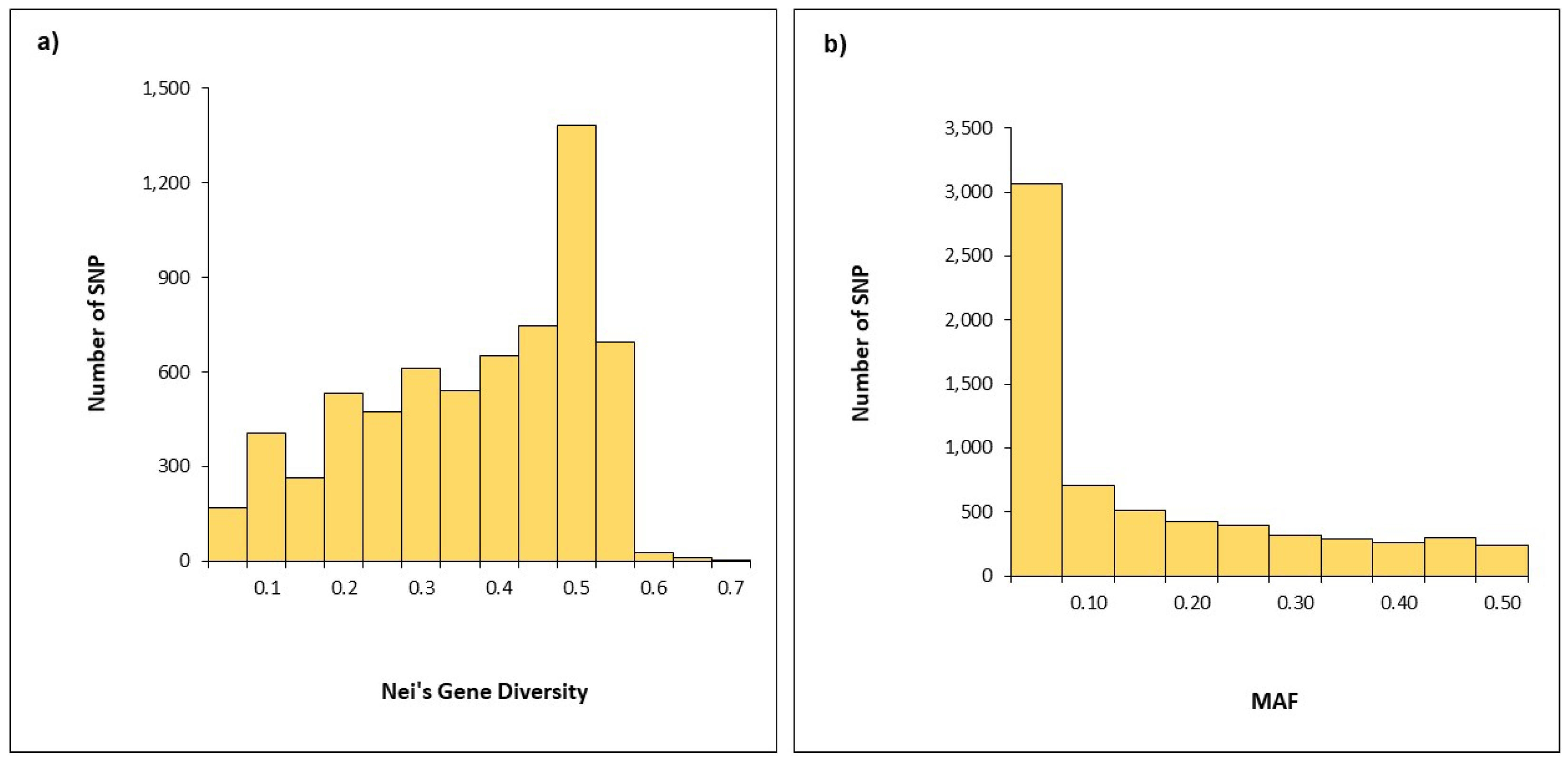
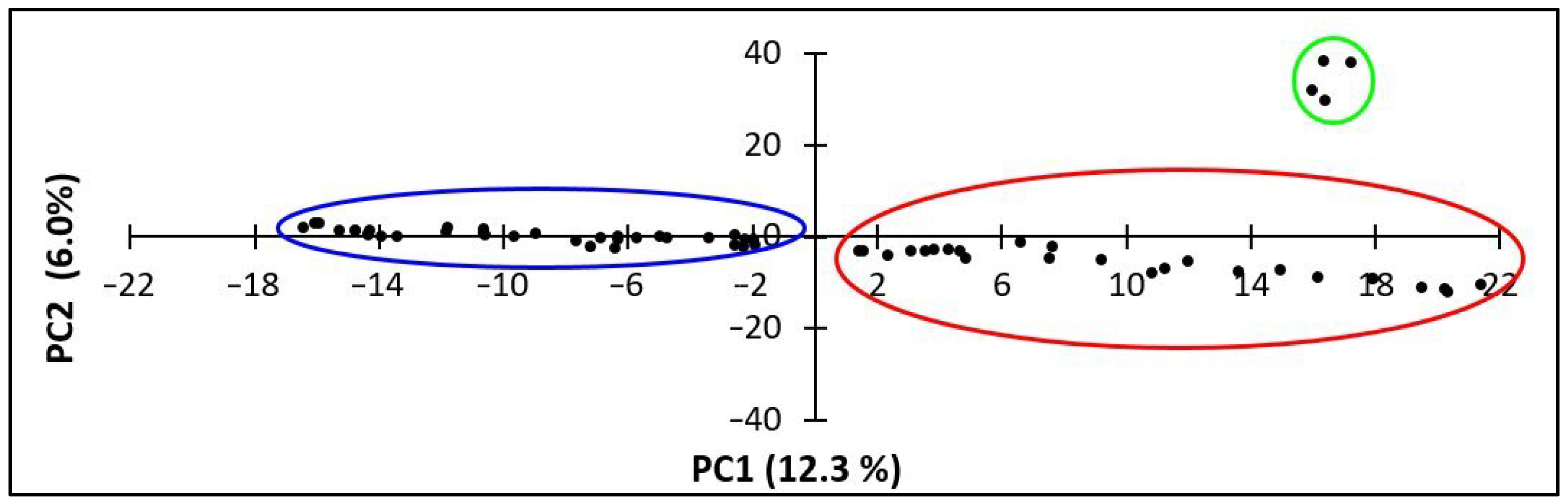
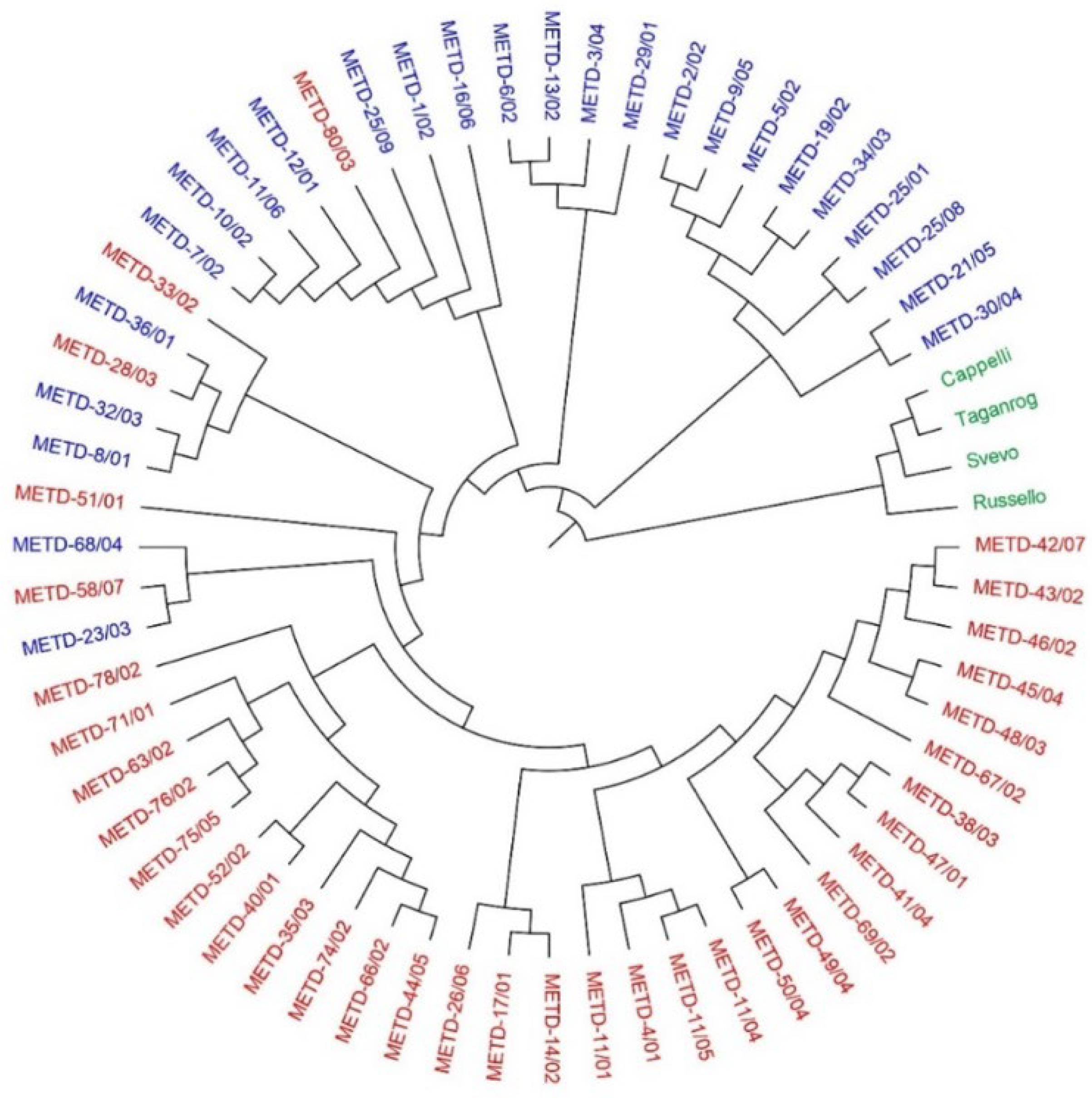
2.2. SNP Diversity in Western Balkan Durum Accessions
2.3. Relationships between Rogosija Durum Clusters and Eco-geographic Conditions
3. Discussion
4. Materials and Methods
4.1. Plant Material and Morphological Trait Characterization
4.2. DNA Extraction and SNP Genotyping
4.3. Genetic Diversity Analysis
4.4. Eco-Geographic Profile of the Germplasm Collection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Talini, R.F.; Brandolini, A.; Miculan, M.; Brunazzi, A.; Vaccino, P.; Enrico Pè, M.; Dell’Acqua, M. Genome-wide association study of agronomic and quality traits in a world collection of the wild wheat relative Triticum urartu. Plant J. 2020, 102, 555–568. [Google Scholar] [CrossRef]
- Jaradat, A.A. Phenotypic divergence in the meta-population of the Hourani durum wheat landrace. J. Food Agric. Environ. 2006, 4, 186–191. [Google Scholar]
- Lopes, M.S.; El-Basyoni, I.; Baenziger, P.S.; Singh, S.; Royo, C.; Ozbek, K.; Aktas, H.; Ozer, E.; Ozdemir, F.; Manickavelu, A.; et al. Exploiting genetic diversity from landraces in wheat breeding for adaptation to climate change. J. Exp. Bot. 2015, 66, 3477–3486. [Google Scholar] [CrossRef]
- Filipović, D. Southwest Asian founder-and other crops at Neolithic sites in Serbia. Bulg. E-J. Archeol. 2014, 4, 195–215. [Google Scholar]
- Pavićević, L.J. Tetraploid and diploid wheat in Montenegro and neighbouring areas. Acta Biol. 1975, 7, 217–307. [Google Scholar]
- Pavićević, L.J. Some positive features of local characteristics of diploid and tetraploid wheat. Genetika 1982, 14, 1–11. [Google Scholar]
- Pavićević, L. Nazivi pšnice. Poljprivreda Šumarstvo 1967, 1, 31–42. [Google Scholar]
- Kassai, M.K. Wheat production, old wheat varieties in Hungary and Vojvodina up to the great war. In IV Naučni Skup Istorija Poljoprivrede, Salaša i Sela, Tema Žito; Pčesa: Novi Sad, Serbia, 1988; Volume 28, pp. 17–25. [Google Scholar]
- Pavićević, L.J. Triticum dicoccum Schübl (krupnici). Matica Srp. 1990, 79, 57–92. [Google Scholar]
- Dimitrijević, M.; Petrović, S.; Cîmpeanu, C.; Bucur, D.; Belić, M. Cereals and Aegilops genus biodiversity survey in the west Balkans: Erosion and preservation. J. Food Agric. Environ. 2011, 9, 219–225. [Google Scholar]
- Díez, M.J.; De la Rosa, L.; Martín, I.; Guasch, L.; Cartea, M.E.; Mallor, C.; Casals, J.; Simó, J.; Rivera, A.; Anastasio, G.; et al. Plant Genebanks: Present situation and proposals for their improvement. The case of the Spanish network. Front. Plant Sci. 2018, 9, 1794. [Google Scholar] [CrossRef]
- Maccaferri, M.; Ricci, A.; Salvi, S.; Milner, S.G.; Noli, E.; Martelli, P.L.; Casadio, R.; Akhunov, E.; Scalabrin, S.; Vendramin, V.; et al. A high-density, SNP-based consensus map of tetraploid wheat as a bridge to integrate durum and bread wheat genomics and breeding. Plant Biotechnol. J. 2015, 13, 648–663. [Google Scholar] [CrossRef] [PubMed]
- Babić, V.; Andjelkovic, V.; Jovovic, Z.; Babic, M.; Vasic, V.; Kravic, N. Diversity Assessment of the Montenegrin maize landrace gene pool maintained in two gene banks. Plants 2021, 10, 1503. [Google Scholar] [CrossRef] [PubMed]
- Mangini, G.; Nigro, D.; Margiotta, B.; De Vita, P.; Gadaleta, A.; Simeone, R.; Blanco, A. Exploring SNP diversity in wheat landraces germplasm and setting of a molecular barcode for fingerprinting. Cereal Res. Commun. 2018, 46, 377–387. [Google Scholar] [CrossRef]
- Cseh, A.; Poczai, P.; Kiss, T.; Balla, K.; Berki, Z.; Horváth, Á.; Kuti, C.; Karsai, I. Exploring the legacy of Central European historical winter wheat landraces. Sci. Rep. 2021, 11, 23915. [Google Scholar] [CrossRef] [PubMed]
- Ganugi, P.; Palchetti, E.; Gori, M.; Calamai, A.; Burridge, A.; Biricolti, S.; Benedettelli, S.; Masoni, A. Molecular diversity within a Mediterranean and European panel of tetraploid wheat (T. turgidum subsp.) landraces and modern germplasm inferred using a high-density SNP array. Agronomy 2021, 11, 414. [Google Scholar] [CrossRef]
- Miazzi, M.M.; Babay, E.; De Vita, P.; Montemurro, C.; Chaabane, R.; Taranto, F.; Mangini, G. Comparative genetic analysis of durum wheat landraces and cultivars widespread in Tunisia. Front. Plant Sci. 2022, 13, 939609. [Google Scholar] [CrossRef]
- Maccaferri, M.; Harris, N.S.; Twardziok, S.O.; Pasam, R.K.; Gundlach, H.; Spannagl, M.; Ormanbekova, D.; Lux, T.; Prade, V.M.; Milner, S.G.; et al. Durum wheat genome highlights past domestication signatures and future improvement targets. Nat. Genet. 2019, 51, 885–895. [Google Scholar] [CrossRef]
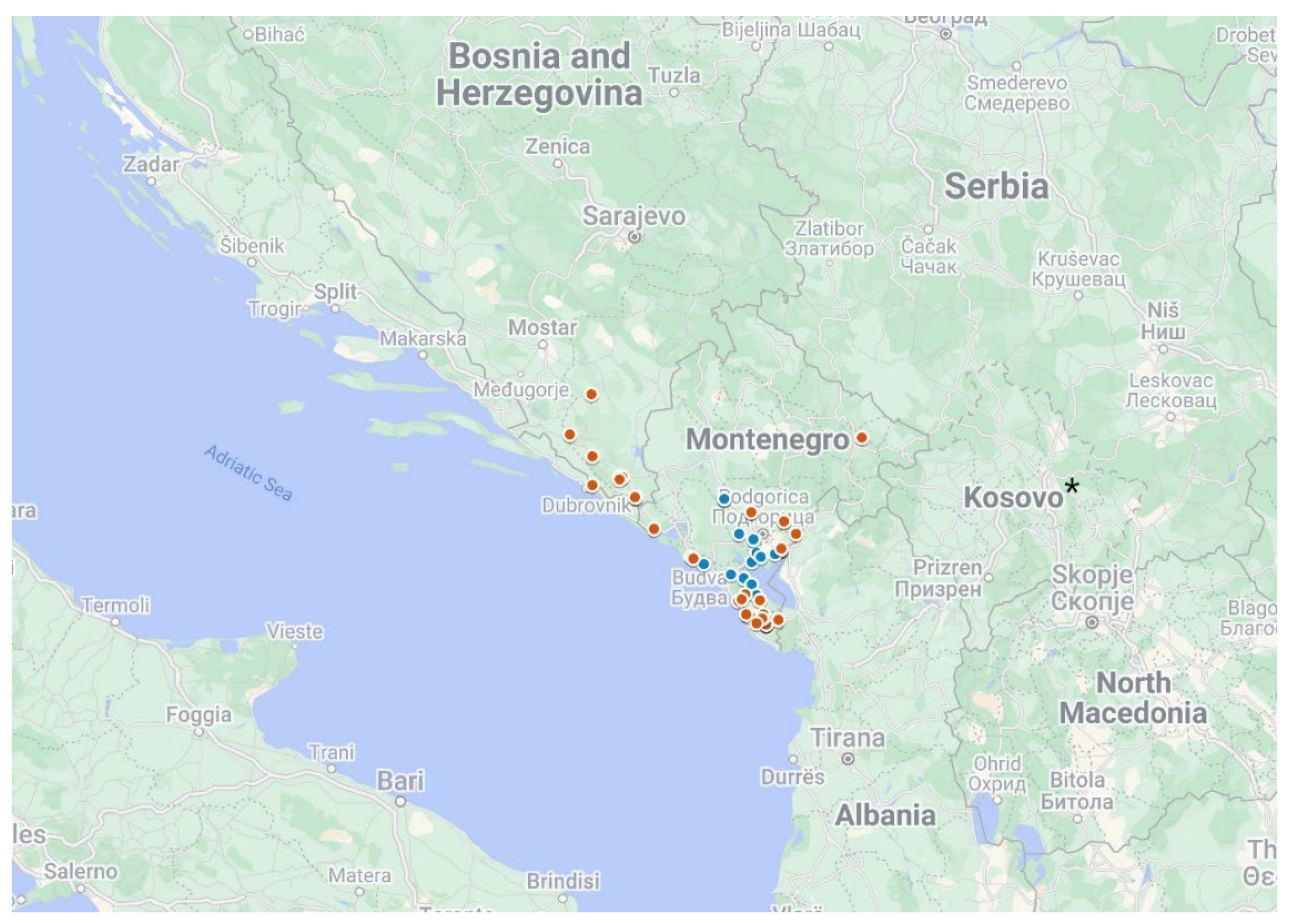
- Durum Wheat Accessions Sampling Sites Location [Google Maps]. Available online: https://www.google.com/maps/d/edit?mid=1oPuGW_FKZYWsRekwJp4Es5TvTYmUWnA&usp=sharing (accessed on 26 January 2023).
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef]
- Pavićević, L. Golozrne tetraploidne pšenice u Crnoj Gori i Hercegovini. Matica Srp. 1972, 43, 125–137. [Google Scholar]
- Jovović, Z. Ratarske kulture. In Genetički Resursi u Biljnoj Proizvodnji Crne Gore, 1st ed.; Pulević, V., Ed.; Crnogorska Akademija Nauka i Umjetnosti: Podgorica, Montenegro, 2021; Volume 1, pp. 303–371. [Google Scholar]
- Jovović, Z.; Mandić, D.; Pržulj, N.; Velimirović, A.; Dolijanović, Ž. Genetic resources of wheat (Triticum sp.) in Montenegro. In Proceedings of the 22nd International Symposium on Biotechnology, Čačak, Serbia, 10–11 March 2017. [Google Scholar]
- Duminil, J.; Di Michele, M. Plant species delimitation: A comparison of morphological and molecular markers. Plant Biosyst. 2009, 143, 528–542. [Google Scholar] [CrossRef]
- Engels, J.M.; Ebert, A.W. A critical review of the current global ex situ conservation system for plant agrobiodiversity. II. Strengths and weaknesses of the current system and recommendations for its improvement. Plants 2021, 10, 1904. [Google Scholar] [CrossRef] [PubMed]
- Weise, S.; Lohwasser, U.; Oppermann, M. Document or lose it—On the importance of information management for genetic resources conservation in genebanks. Plants 2020, 9, 1050. [Google Scholar] [CrossRef] [PubMed]
- Mengistu, D.K.; Kidane, Y.G.; Fadda, C.; Pè, M.E. Genetic diversity in Ethiopian durum wheat (Triticum turgidum var durum) inferred from phenotypic variations. Plant Genet. Resour. 2016, 16, 39–49. [Google Scholar] [CrossRef]
- Fiore, M.C.; Mercati, F.; Spina, A.; Blangiforti, S.; Venora, G.; Dell’Acqua, M.; Lupini, A.; Preiti, G.; Monti, M.; Pè, M.E.; et al. High-throughput genotype, morphology, and quality traits evaluation for the assessment of genetic diversity of wheat landraces from Sicily. Plants 2019, 8, 116. [Google Scholar] [CrossRef]
- Ouaja, M.; Bahri, B.A.; Aouini, L.; Ferjaoui, S.; Medini, M.; Marcel, T.C.; Hamza, S. Morphological characterization and genetic diversity analysis of Tunisian durum wheat (Triticum turgidum var. durum) accessions. BMC Genom. Data 2021, 22, 3. [Google Scholar] [CrossRef] [PubMed]
- Takač, V.; Mikić, S.; Mirosavljević, M.; Momčilović, V.; Trkulja, D.; Kondić Spika, A.; Brbaklić, L. Characterisation of Serbian durum wheat genotypes based on UPOV–defined characteristics. Ratar. Povrt. 2019, 56, 97–102. [Google Scholar] [CrossRef]
- Kabbaj, H.; Sall, A.T.; Al-Abdallat, A.; Geleta, M.; Amri, A.; Filali-Maltouf, A.; Belkadi, B.; Ortiz, R.; Bassi, F.M. Genetic diversity within a global panel of durum wheat (Triticum durum) landraces and modern germplasm reveals the history of alleles exchange. Front. Plant Sci. 2017, 8, 1277. [Google Scholar] [CrossRef]
- Mangini, G.; Margiotta, B.; Marcotuli, I.; Signorile, M.A.; Gadaleta, A.; Blanco, A. Genetic diversity and phenetic analysis in wheat (Triticum turgidum subsp. durum and Triticum aestivum subsp. aestivum) landraces based on SNP markers. Genet. Res. Crop Evol. 2017, 64, 1269–1280. [Google Scholar]
- Mazzucotelli, E.; Sciara, G.; Mastrangelo, A.M.; Desiderio, F.; Xu, S.S.; Faris, J.; Hayden, M.J.; Tricker, P.J.; Ozkan, H.; Echenique, V.; et al. Global durum wheat panel (GDP): An international platform to identify and exchange beneficial alleles. Front. Plant Sci. 2020, 11, 569905. [Google Scholar] [CrossRef]
- Alemu, A.; Feyissa, T.; Letta, T.; Abeyo, B. Genetic diversity and population structure analysis based on the high-density SNP markers in Ethiopian durum wheat (Triticum turgidum ssp. durum). BMC Genet. 2020, 21, 18. [Google Scholar] [CrossRef]
- Dracatos, P.M.; Budak, H.; Sourdille, P.; Perovic, D. 2022) Editorial: Use of barley and wheat reference sequences: Downstream applications in breeding, gene isolation, GWAS, and evolution-volume II. Front. Plant Sci. 2022, 13, 1034744. [Google Scholar] [CrossRef] [PubMed]
- Xynias, I.N.; Mylonas, I.; Korpetis, E.G.; Ninou, E.; Tsaballa, A.; Avdikos, I.D.; Mavromatis, A.G. Durum wheat breeding in the Mediterranean region: Current status and future prospects. Agronomy 2020, 10, 432. [Google Scholar] [CrossRef]
- Pavićević, L. Važnije osobine i uzgoj nekih italijanskih sorti pšenice kod nas. Naša Poljopr. 1958, 4, 3–13. [Google Scholar]
- Fairchild, D.G. Saragolla Wheat. In Miscelaneous Papers, 1st ed.; Bulletin no. 25; Galloway, B.T., Ed.; U.S. Department of Agriculture, Bureau of Plant Industry: Washington, DC, USA, 1903. [Google Scholar]
- Nazco, R.; Villegas, D.; Ammar, K.; Peña, R.J.; Moragues, M.; Royo, C. Can Mediterranean durum wheat landraces contribute to improved grain quality attributes in modern cultivars? Euphytica 2012, 185, 1–17. [Google Scholar] [CrossRef]
- Dedkova, O.S.; Badaeva, E.D.; Amosova, A.V.; Martynov, S.P.; Ruanet, V.V.; Mitrofanova, O.P.; Pukhal’skiy, V.A. Diversity and the origin of the European population of Triticum dicoccum (Schrank) Schuebl. as revealed by chromosome analysis. Russ. J. Genet. 2009, 45, 1082–1091. [Google Scholar] [CrossRef]
- Slim, A.; Piarulli, L.; Chennaoui Kourda, H.; Rouaissi, M.; Robbana, C.; Chaabane, R.; Pignone, D.; Montemurro, C.; Mangini, G. Genetic structure analysis of a collection of Tunisian durum wheat germplasm. Int. J. Mol. Sci. 2019, 20, 3362. [Google Scholar] [CrossRef]
- Fadida-Myers, A.; Fuerst, D.; Tzuberi, A.; Yadav, S.; Nashef, K.; Roychowdhury, R.; Sansaloni, C.P.; Hübner, S.; Ben-David, R. Emmer wheat eco-geographic and genomic congruence shapes phenotypic performance under Mediterranean climate. Plants 2022, 11, 1460. [Google Scholar] [CrossRef]
- Zdunić, G.; Lukšić, K.; Nagy, Z.A.; Mucalo, A.; Hančević, K.; Radić, T.; Butorac, L.; Jahnke, G.G.; Kiss, E.; Ledesma-Krist, G.; et al. Genetic structure and relationships among wild and cultivated grapevines from central Europe and part of the Western Balkan Peninsula. Genes 2020, 11, 962. [Google Scholar] [CrossRef]
- Pavan, S.; Bardaro, N.; Fanelli, V.; Marcotrigiano, A.R.; Mangini, G.; Taranto, F.; Catalano, D.; Montemurro, C.; De Giovanni, C.; Lotti, C.; et al. Genotyping by sequencing of cultivated lentil (Lens culinaris Medik.) highlights population structure in the mediterranean gene pool associated with geographic patterns and phenotypic variables. Front. Genet. 2019, 10, 872. [Google Scholar] [CrossRef]
- Ninou, E.G.; Mylonas, I.G.; Tsivelikas, A.L.; Ralli, P.E. Phenotypic diversity of Greek dill (Anethum graveolens L.) landraces. Acta Agric. Scand. Sect. B Soil Plant Sci. 2017, 67, 318–325. [Google Scholar]
- International Union for the Protection of new Varieties of Plants. Guidelines for the Conduct of Tests for Distinctness, Homogeneity and Stability, Durum Wheat (Triticum durum Desf.); International Union for the Protection of New Varieties of Plants: Geneva, Switzerland, 1988; pp. 3–32. [Google Scholar]
- Soleimani, B.; Lehnert, H.; Keilwagen, J.; Plieske, J.; Ordon, F.; Naseri Rad, S.; Ganal, M.; Beier, S.; Perovic, D. Comparison between core set selection methods using different Illumina marker platforms: A case study of assessment of diversity in wheat. Front. Plant Sci. 2020, 11, 1040. [Google Scholar] [CrossRef] [PubMed]
- Winfield, M.O.; Allen, A.M.; Burridge, A.; Barker, G.L.A.; Benbow, H.R.; Wilkinson, P.A.; Coghill, J.; Waterfall, C.; Davassi, A.; Scopes, G.; et al. High-density SNP genotyping array for hexaploid wheat and its secondary and tertiary gene pool. Plant Biotechnol. J. 2016, 14, 1195–1206. [Google Scholar] [CrossRef] [PubMed]
- Jolliffe, I.T. Principal Component Analysis, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Andrew, R. FigTree: Tree Figure Drawing Tool, Version 1.4.3; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2016.
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic analysis in excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Micev, B.; Pavićević, D. Meteorological Data of Montenegro. Unpublished work. 2022. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chromosome | Nei’s Gene Diversity | MAF |
|---|---|---|
| 1A | 0.361 | 0.149 |
| 2A | 0.338 | 0.136 |
| 3A | 0.327 | 0.136 |
| 4A | 0.353 | 0.155 |
| 5A | 0.328 | 0.118 |
| 6A | 0.331 | 0.133 |
| 7A | 0.328 | 0.117 |
| A-Genome | 0.338 | 0.135 |
| 1B | 0.345 | 0.149 |
| 2B | 0.352 | 0.136 |
| 3B | 0.368 | 0.134 |
| 4B | 0.339 | 0.159 |
| 5B | 0.313 | 0.136 |
| 6B | 0.355 | 0.157 |
| 7B | 0.331 | 0.116 |
| B-Genome | 0.343 | 0.141 |
| Whole Genome | 0.341 | 0.138 |
| Source of Variation | df | SS | MS | Est. Var. | % | p Values |
|---|---|---|---|---|---|---|
| Among clusters | 1 | 6722.42 | 6722.42 | 194.23 | 16% | <0.001 |
| Within cluster | 58 | 61,333.55 | 1057.47 | 1057.47 | 84% | |
| Total | 59 | 68,055.97 | 1251.70 | 100% |
| Western Balkan Cluster | Number of Accession | Ne | I | h | PPL |
|---|---|---|---|---|---|
| 1 | 25 | 1.443 | 0.416 | 0.266 | 88.24 |
| 2 | 35 | 1.577 | 0.510 | 0.321 | 88.55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velimirović, A.; Jovović, Z.; Perović, D.; Lehnert, H.; Mikić, S.; Mandić, D.; Pržulj, N.; Mangini, G.; Finetti-Sialer, M.M. SNP Diversity and Genetic Structure of “Rogosija”, an Old Western Balkan Durum Wheat Collection. Plants 2023, 12, 1157. https://doi.org/10.3390/plants12051157
Velimirović A, Jovović Z, Perović D, Lehnert H, Mikić S, Mandić D, Pržulj N, Mangini G, Finetti-Sialer MM. SNP Diversity and Genetic Structure of “Rogosija”, an Old Western Balkan Durum Wheat Collection. Plants. 2023; 12(5):1157. https://doi.org/10.3390/plants12051157
Chicago/Turabian StyleVelimirović, Ana, Zoran Jovović, Dragan Perović, Heike Lehnert, Sanja Mikić, Dragan Mandić, Novo Pržulj, Giacomo Mangini, and Mariella Matilde Finetti-Sialer. 2023. "SNP Diversity and Genetic Structure of “Rogosija”, an Old Western Balkan Durum Wheat Collection" Plants 12, no. 5: 1157. https://doi.org/10.3390/plants12051157
APA StyleVelimirović, A., Jovović, Z., Perović, D., Lehnert, H., Mikić, S., Mandić, D., Pržulj, N., Mangini, G., & Finetti-Sialer, M. M. (2023). SNP Diversity and Genetic Structure of “Rogosija”, an Old Western Balkan Durum Wheat Collection. Plants, 12(5), 1157. https://doi.org/10.3390/plants12051157