
Native Microbes Amplify Native Seedling Establishment and Diversity While Inhibiting a Non-Native Grass

Abstract
:1. Introduction
2. Results
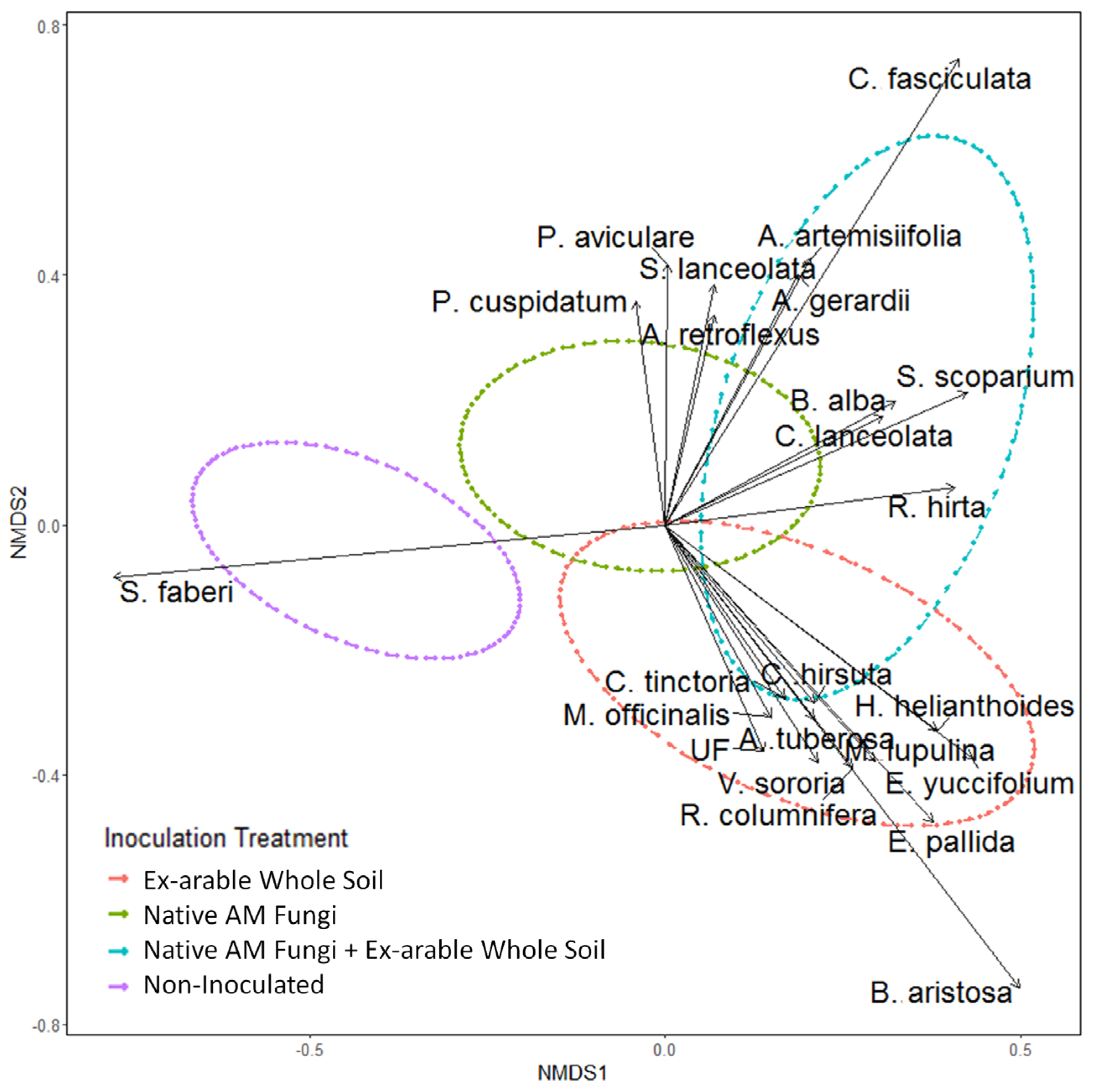
2.1. Plant Community Composition among Soil Inoculation Treatments
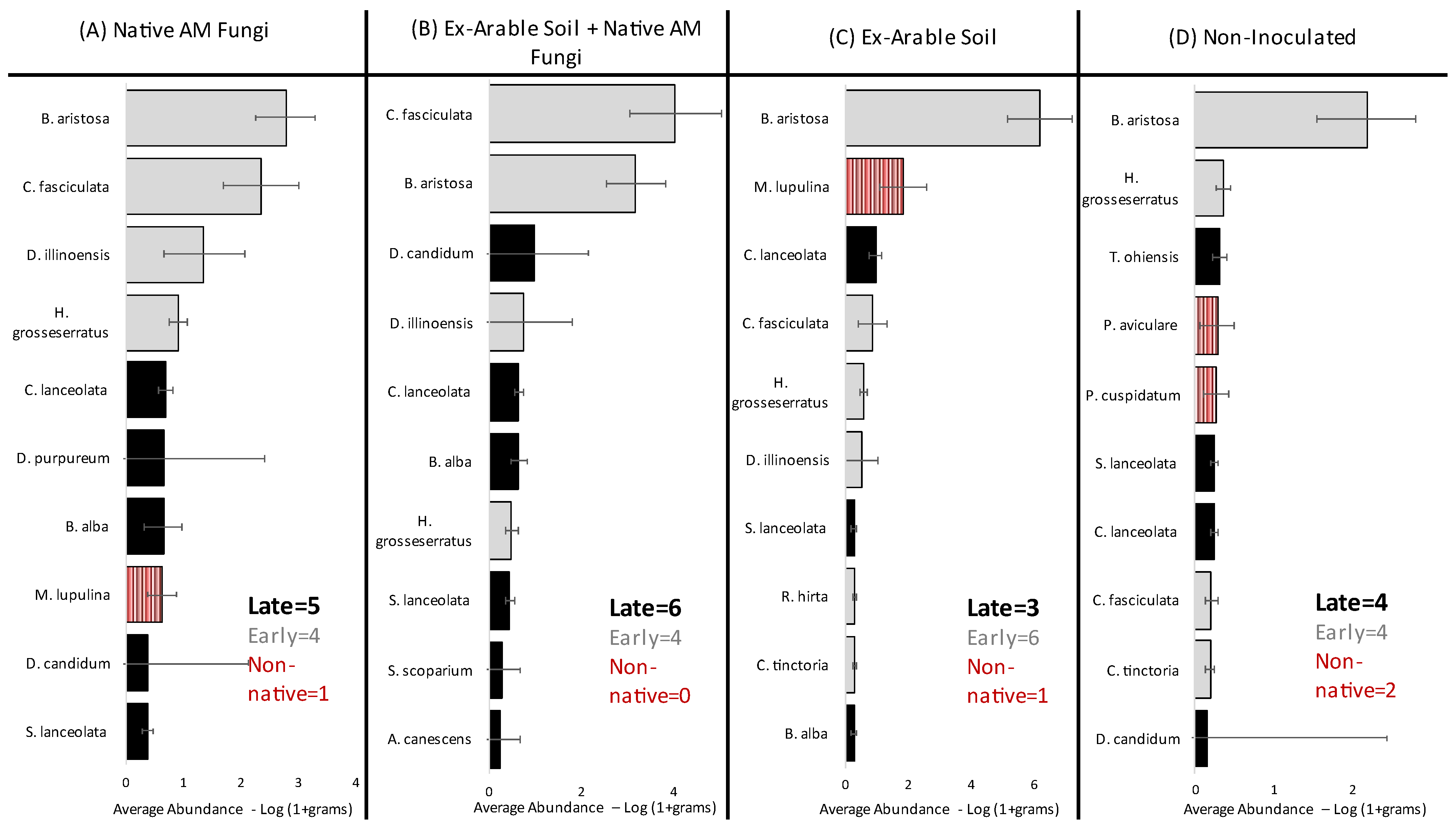
2.2. Species-Specific and Diversity Responses to Soil Inoculation Treatments
2.3. Inoculation Driven Shifts in Invasion, Successional Processes, and Functional Group Representation
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Methods
5.2. Inoculum & Seed Collection
5.3. Data Collection
5.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rowe, H.I. Tricks of the Trade: Techniques and Opinions from 38 Experts in Tallgrass Prairie Restoration. Restor. Ecol. 2010, 18, 253–262. [Google Scholar] [CrossRef]
- Török, P.; Vida, E.; Deák, B.; Lengyel, S.; Tóthmérész, B. Grassland restoration on former croplands in Europe: An assessment of applicability of techniques and costs. Biodivers. Conserv. 2011, 20, 2311–2332. [Google Scholar] [CrossRef]
- Riebkes, J.L.; Barak, R.S.; Kramer, A.T. Evaluating seed viability in prairie forbs: A test of three methods. Nativ. Plants J. 2015, 16, 96–106. [Google Scholar] [CrossRef]
- Shaw, N.; Barak, R.S.; Campbell, R.E.; Kirmer, A.; Pedrini, S.; Dixon, K.; Frischie, S. Seed use in the field: Delivering seeds for restoration success. Restor. Ecol. 2020, 28, S276–S285. [Google Scholar] [CrossRef]
- Kindscher, K.; Tieszen, L.L. Floristic and soil organic matter changes after five and thirty-five years of native tallgrass prairie restoration. Restor. Ecol. 1998, 6, 181–196. [Google Scholar] [CrossRef]
- Goldblum, D.; Glaves, B.P.; Rigg, L.S.; Kleiman, B. The Impact of Seed Mix Weight on Diversity and Species Composition in a Tallgrass Prairie Restoration Planting, Nachusa Grasslands, Illinois, USA. Ecol. Restor. 2013, 31, 154–167. [Google Scholar] [CrossRef]
- Allison, S.K. When Is a Restoration Successful? Results from a 45-Year-Old Tallgrass Prairie Restoration. Ecol. Restor. 2002, 20, 10–17. [Google Scholar] [CrossRef]
- Koziol, L.; A Schultz, P.; House, G.L.; Bauer, J.T.; Middleton, E.L.; Bever, J.D. The Plant Microbiome and Native Plant Restoration: The Example of Native Mycorrhizal Fungi. Bioscience 2018, 68, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Koziol, L.; Bever, J.D. Mycorrhizal response trades off with plant growth rate and increases with plant successional status. Ecology 2015, 96, 1768–1774. [Google Scholar] [CrossRef]
- Cheeke, T.E.; Zheng, C.; Koziol, L.; Gurholt, C.R.; Bever, J.D. Sensitivity to AMF species is greater in late-successional native than early-successional native or non-native grassland plants. Ecology 2019, 100, e02855. [Google Scholar] [CrossRef] [Green Version]
- Middleton, E.L.; Richardson, S.; Koziol, L.; Palmer, C.E.; Yermakov, Z.; Henning, J.A.; Schultz, P.A.; Bever, J.D. Locally adapted arbuscular mycorrhizal fungi improve vigor and resistance to herbivory of native prairie plant species. Ecosphere 2015, 6, 276. [Google Scholar] [CrossRef]
- Koziol, L.; Bauer, J.T.; Duell, E.B.; Hickman, K.; House, G.L.; Schultz, P.A.; Tipton, A.G.; Wilson, G.W.T.; Bever, J.D. Manipulating plant microbiomes in the field: Native mycorrhizae advance plant succession and improve native plant restoration. J. Appl. Ecol. 2022, 59, 1976–1985. [Google Scholar] [CrossRef]
- Koziol, L.; Bever, J.D. The missing link in grassland restoration: Arbuscular mycorrhizal fungi inoculation increases plant diversity and accelerates succession. J. Appl. Ecol. 2017, 54, 1301–1309. [Google Scholar] [CrossRef] [Green Version]
- Duell, E.B.; Hickman, K.R.; Wilson, G.W.T.; Zhang, Y. Inoculation with native grassland soils improves native plant species germination in highly disturbed soil. Grassl. Res. 2022, 1, 75–83. [Google Scholar] [CrossRef]
- Ezáčová, V.; Řezáč, M.; Wilson, G.W.; Michalová, T. Arbuscular mycorrhiza can be disadvantageous for weedy annuals in competition with paired perennial plants. Sci. Rep. 2022, 12, 20703. [Google Scholar] [CrossRef]
- Duell, E.B.; O’Hare, A.; Wilson, G.W.T. Inoculation with native soil improves seedling survival and reduces non-native reinvasion in a grassland restoration. Restor. Ecol. 2022, 31, e13685. [Google Scholar] [CrossRef]
- Middleton, E.L.; Bever, J.D.; Schultz, P.A. The effect of restoration methods on the quality of the restoration and resistance to invasion by exotics. Restor. Ecol. 2010, 18, 181–187. [Google Scholar] [CrossRef]
- Martin, L.M.; Wilsey, B.J. Native-species seed additions do not shift restored prairie plant communities from exotic to native states. Basic Appl. Ecol. 2014, 15, 297–304. [Google Scholar] [CrossRef]
- Schuster, M.J.; Wragg, P.D.; Reich, P.B. Using revegetation to suppress invasive plants in grasslands and forests. J. Appl. Ecol. 2018, 55, 2362–2373. [Google Scholar] [CrossRef]
- Hess, M.C.; Mesléard, F.; Young, T.P.; de Freitas, B.; Haveneers, N.; Buisson, E. Altering native community assembly history influences the performance of an annual invader. Basic Appl. Ecol. 2022, 59, 70–81. [Google Scholar] [CrossRef]
- Ren, Y.; Lü, Y.; Fu, B. Quantifying the impacts of grassland restoration on biodiversity and ecosystem services in China: A me-ta-analysis. Ecol. Eng. 2016, 95, 542–550. [Google Scholar] [CrossRef]
- Brudvig, L.A.; Barak, R.S.; Bauer, J.T.; Caughlin, T.T.; Laughlin, D.C.; Larios, L.; Matthews, J.W.; Stuble, K.L.; Turley, N.E.; Zirbel, C.R. Interpreting variation to advance predictive restoration science. J. Appl. Ecol. 2017, 54, 1018–1027. [Google Scholar] [CrossRef] [Green Version]
- Eldridge, D.J.; Travers, S.K.; Val, J.; Ding, J.; Wang, J.; Singh, B.K.; Delgado-Baquerizo, M. Experimental evidence of strong relationships between soil microbial communities and plant germination. J. Ecol. 2021, 109, 2488–2498. [Google Scholar] [CrossRef]
- Lubin, T.K.; Schultz, P.; Bever, J.D.; Alexander, H.M. Are two strategies better than one? Manipulation of seed density and soil community in an experimental prairie restoration. Restor. Ecol. 2019, 27, 1021–1031. [Google Scholar] [CrossRef]
- House, G.L.; Bever, J.D. Biochar soil amendments in prairie restorations do not interfere with benefits from inoculation with native arbuscular mycorrhizal fungi. Restor. Ecol. 2019, 28, 785–795. [Google Scholar] [CrossRef]
- Koziol, L.; McKenna, T.P.; Crews, T.E.; Bever, J.D. Native arbuscular mycorrhizal fungi promote native grassland diversity and suppress weeds 4 years following inoculation. Restor. Ecol. 2022, e13772, in press. [Google Scholar] [CrossRef]
- Koziol, L.; Crews, T.E.; Bever, J.D. Native plant abundance, diversity, and richness increases in prairie restoration with field inoculation density of native mycorrhizal amendments. Restor. Ecol. 2020, 28, S373–S380. [Google Scholar] [CrossRef]
- Gardner, H.W. Wet-Mesic to Wet Soil-Preferring Species. In Tallgrass Prairie Restoration in the Midwestern and Eastern United States; Springer: Berlin/Heidelberg, Germany, 2011; pp. 145–160. [Google Scholar]
- Haddock, M.J.; Freeman, C.C.; Bare, J. Kansas Wildflowers and Weeds; University Press of Kansas: Lawrence, KS, USA, 2015. [Google Scholar]
- Koziol, L.; Bever, J.D. Mycorrhizal feedbacks generate positive frequency dependence accelerating grassland succession. J. Ecol. 2019, 107, 622–632. [Google Scholar] [CrossRef] [Green Version]
- Bauer, J.T.; Koziol, L.; Bever, J.D. Ecology of Floristic Quality Assessment: Testing for correlations between coefficients of conservatism, species traits and mycorrhizal responsiveness. AoB Plants 2018, 10, plx073. [Google Scholar] [CrossRef]
- Grman, E.; Allen, J.; Galloway, E.; McBride, J.; Bauer, J.T.; Price, P.A. Inoculation with remnant prairie soils increased the growth of three native prairie legumes but not necessarily their associations with beneficial soil microbes. Restor. Ecol. 2020, 28, S393–S399. [Google Scholar] [CrossRef]
- Middleton, E.; Bever, J.D. Inoculation with a Native Soil Community Advances Succession in a Grassland Restoration. Restor. Ecol. 2012, 20, 218–226. [Google Scholar] [CrossRef]
- Wubs, E.R.J.; Van Der Putten, W.H.; Mortimer, S.R.; Korthals, G.W.; Duyts, H.; Wagenaar, R.; Bezemer, M. Single introductions of soil biota and plants generate long-term legacies in soil and plant community assembly. Ecol. Lett. 2019, 22, 1145–1151. [Google Scholar] [CrossRef] [PubMed]
- Vahter, T.; Bueno, C.G.; Davison, J.; Herodes, K.; Hiiesalu, I.; Kasari-Toussaint, L.; Oja, J.; Olsson, P.A.; Sepp, S.; Zobel, M.; et al. Co-introduction of native mycorrhizal fungi and plant seeds accelerates restoration of post-mining landscapes. J. Appl. Ecol. 2020, 57, 1741–1751. [Google Scholar] [CrossRef]
- Wang, G.; Schultz, P.; Tipton, A.; Zhang, J.; Zhang, F.; Bever, J.D. Soil microbiome mediates positive plant diversity-productivity relationships in late successional grassland species. Ecol. Lett. 2019, 22, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Stevens, M.H.H.; Oksanen, M.J.; Suggests, M.A.S.S. The vegan package. Community Ecol. Package 2007, 10, 719. [Google Scholar]
- Hervé, M.; Hervé, M.M. Package ‘RVAideMemoire’. 2020. Available online: https://CRANR-projectorg/package=RVAideMemoire (accessed on 19 January 2023).
- SAS. SAS 9.4 User’s Guide: Survey Data Analysis; SAS Institute Inc.: Cary, NC, USA, 2015. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | NMDS1 | NMDS2 | p-Value |
|---|---|---|---|
| B. aristosa | 0.4978811 | −0.741185 | 0.001 |
| E. pallida | 0.3758625 | −0.475515 | 0.001 |
| C. fasciculata | 0.4120677 | 0.7460587 | 0.001 |
| S. faberi | −0.77588 | −0.083553 | 0.001 |
| E. yuccifolium | 0.430899 | −0.373672 | 0.002 |
| H. helianthoides | 0.3823069 | −0.329738 | 0.003 |
| R. columnifera | 0.2631774 | −0.388539 | 0.004 |
| M. lupulina | 0.2941195 | −0.378259 | 0.004 |
| A. artemisiifolia | 0.2044813 | 0.4260718 | 0.005 |
| S. scoparium | 0.4244076 | 0.2123334 | 0.009 |
| P. aviculare | 0.0028758 | 0.4169241 | 0.016 |
| V. sororia | 0.2141607 | −0.380134 | 0.018 |
| R. hirta | 0.4068963 | 0.0594642 | 0.024 |
| UF (Unknown Forb) | 0.1385269 | −0.362878 | 0.03 |
| B. alba | 0.323966 | 0.1972003 | 0.035 |
| S. lanceolata | 0.0683162 | 0.3843318 | 0.036 |
| A. tuberosa | 0.2083887 | −0.310465 | 0.045 |
| P. cuspidatum | −0.042617 | 0.3576915 | 0.048 |
| A. gerardii | 0.1858767 | 0.4012776 | 0.049 |
| C. lanceolata | 0.3055674 | 0.1741912 | 0.071 |
| A. retroflexus | 0.0682149 | 0.3365135 | 0.076 |
| C. hirsuta | 0.2094112 | −0.284899 | 0.08 |
| C. tinctoria | 0.1679516 | −0.278248 | 0.09 |
| M. officinalis | 0.1491952 | −0.308403 | 0.097 |
| (A) | Total Diversity | Total Abundance | Native Abundance | Non-Native Abundance | Relative Invasion | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Predictors | DF | F Value | p Value | F Value | p Value | F Value | p Value | F Value | p Value | F Value | p Value |
| Inoculation Treatment | 3 | 8.62 | 0.0003 | 1.2 | 0.326 | 16.43 | <0.0001 | 9.41 | 0.0002 | 19.72 | <0.0001 |
| Block | 10 | 1.58 | 0.165 | 2.13 | 0.054 | 2.43 | 0.0292 | 0.65 | 0.7586 | 1.66 | 0.1367 |
| Contrasts | |||||||||||
| Inoculated vs. Non- Inoculated | 1 | 15 | 0.0006 | 1.85 | 0.184 | 48.17 | <0.0001 | 17.26 | 0.0002 | 51.92 | <0.0001 |
| Inoculated with Native AM Fungi vs. Not | 1 | 15.08 | 0.0006 | 0.75 | 0.393 | 18.23 | 0.0002 | 7.16 | 0.0119 | 21.01 | <0.0001 |
| Ex-arable Soil vs. Native AM Fungi + Ex-Arable Soil | 1 | 8.95 | 0.0057 | 0.33 | 0.569 | 0.61 | 0.4409 | 3.77 | 0.0616 | 3.13 | 0.0871 |
| Differences Among Live Inocula | 2 | 5.43 | 0.0102 | 0.88 | 0.426 | 0.56 | 0.5785 | 5.48 | 0.0093 | 3.62 | 0.0391 |
| (B) | Early Successional Plants | Late Successional Plants | Native Forbs | Native Grasses | Native Legumes | ||||||
| Predictors | DF | F Value | pValue | F Value | pValue | F Value | pValue | F Value | pValue | F Value | pValue |
| Inoculation Treatment | 3 | 10.52 | <0.0001 | 4.93 | 0.007 | 9.87 | 0.0001 | 5.03 | 0.0061 | 16.88 | <0.0001 |
| Block | 10 | 2.41 | 0.0305 | 1.15 | 0.359 | 3.23 | 0.0062 | 2.57 | 0.0221 | 0.94 | 0.5102 |
| Contrasts | |||||||||||
| Inoculated vs. Non- Inoculated | 1 | 30.52 | <0.0001 | 13.04 | 0.001 | 19.46 | 0.0001 | 8.55 | 0.0065 | 29.62 | <0.0001 |
| Inoculated with Native AM Fungi vs. Not | 1 | 8.52 | 0.0066 | 9.9 | 0.004 | 0 | 0.959 | 3.82 | 0.0601 | 46.89 | <0.0001 |
| Ex-arable Soil vs. Native AM Fungi +Ex-Arable Soil | 1 | 0.04 | 0.8472 | 1.51 | 0.229 | 7.82 | 0.0089 | 2.4 | 0.1315 | 18.16 | 0.0002 |
| Differences Among Live Inocula | 2 | 0.52 | 0.6018 | 0.87 | 0.431 | 5.07 | 0.0127 | 3.27 | 0.0518 | 10.52 | 0.0003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koziol, L.; McKenna, T.P.; Bever, J.D. Native Microbes Amplify Native Seedling Establishment and Diversity While Inhibiting a Non-Native Grass. Plants 2023, 12, 1184. https://doi.org/10.3390/plants12051184
Koziol L, McKenna TP, Bever JD. Native Microbes Amplify Native Seedling Establishment and Diversity While Inhibiting a Non-Native Grass. Plants. 2023; 12(5):1184. https://doi.org/10.3390/plants12051184
Chicago/Turabian StyleKoziol, Liz, Thomas P. McKenna, and James D. Bever. 2023. "Native Microbes Amplify Native Seedling Establishment and Diversity While Inhibiting a Non-Native Grass" Plants 12, no. 5: 1184. https://doi.org/10.3390/plants12051184
APA StyleKoziol, L., McKenna, T. P., & Bever, J. D. (2023). Native Microbes Amplify Native Seedling Establishment and Diversity While Inhibiting a Non-Native Grass. Plants, 12(5), 1184. https://doi.org/10.3390/plants12051184