
Plant Species’ Capacity for Range Shifts at the Habitat and Geographic Scales: A Trade-Off-Based Framework


Abstract
:1. Introduction
2. What’s in a Range? Defining Geographic and Habitat Ranges
3. Ecological Strategies and Trade-Offs
4. Colonization Ability

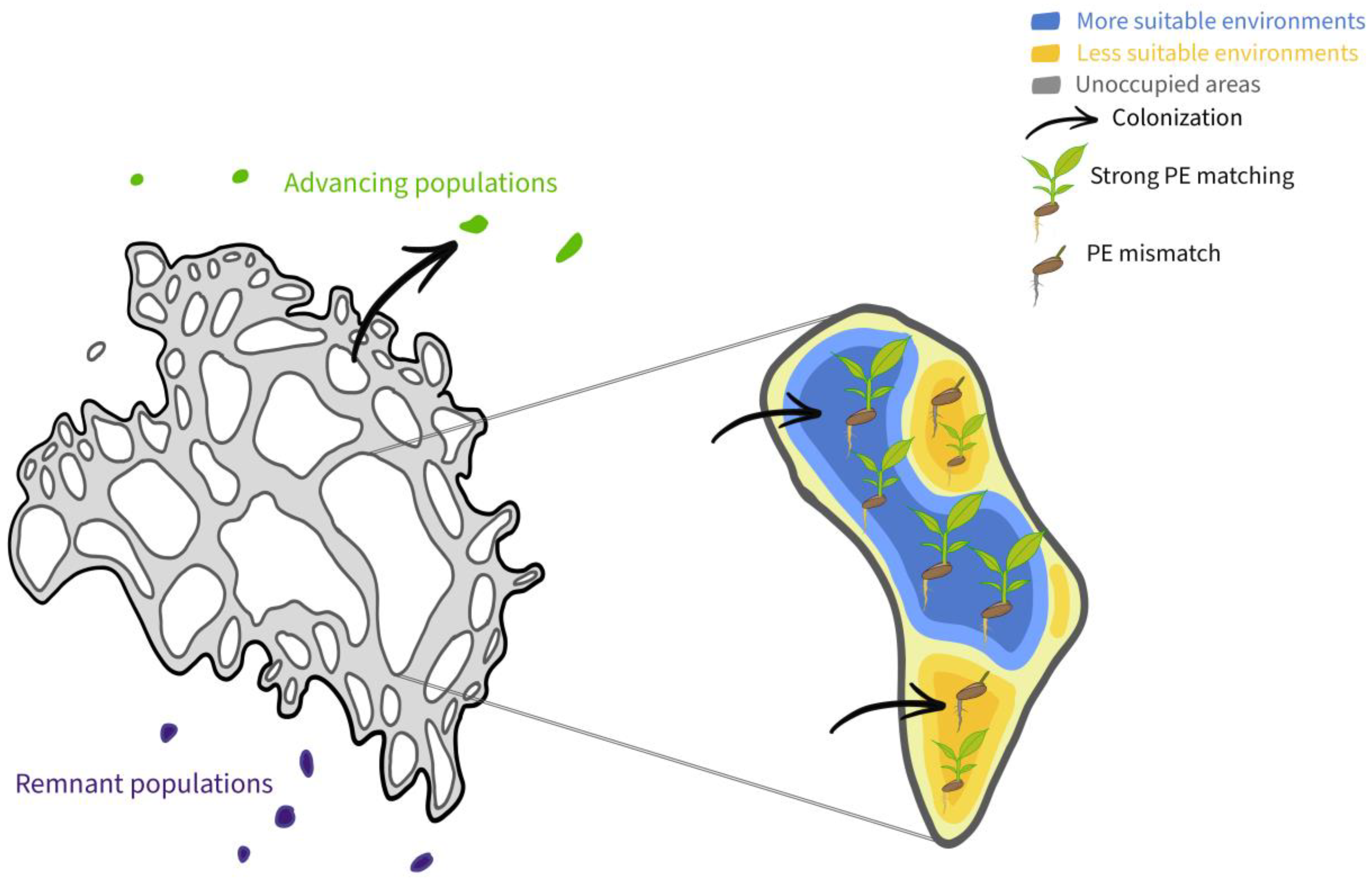{kind=link}
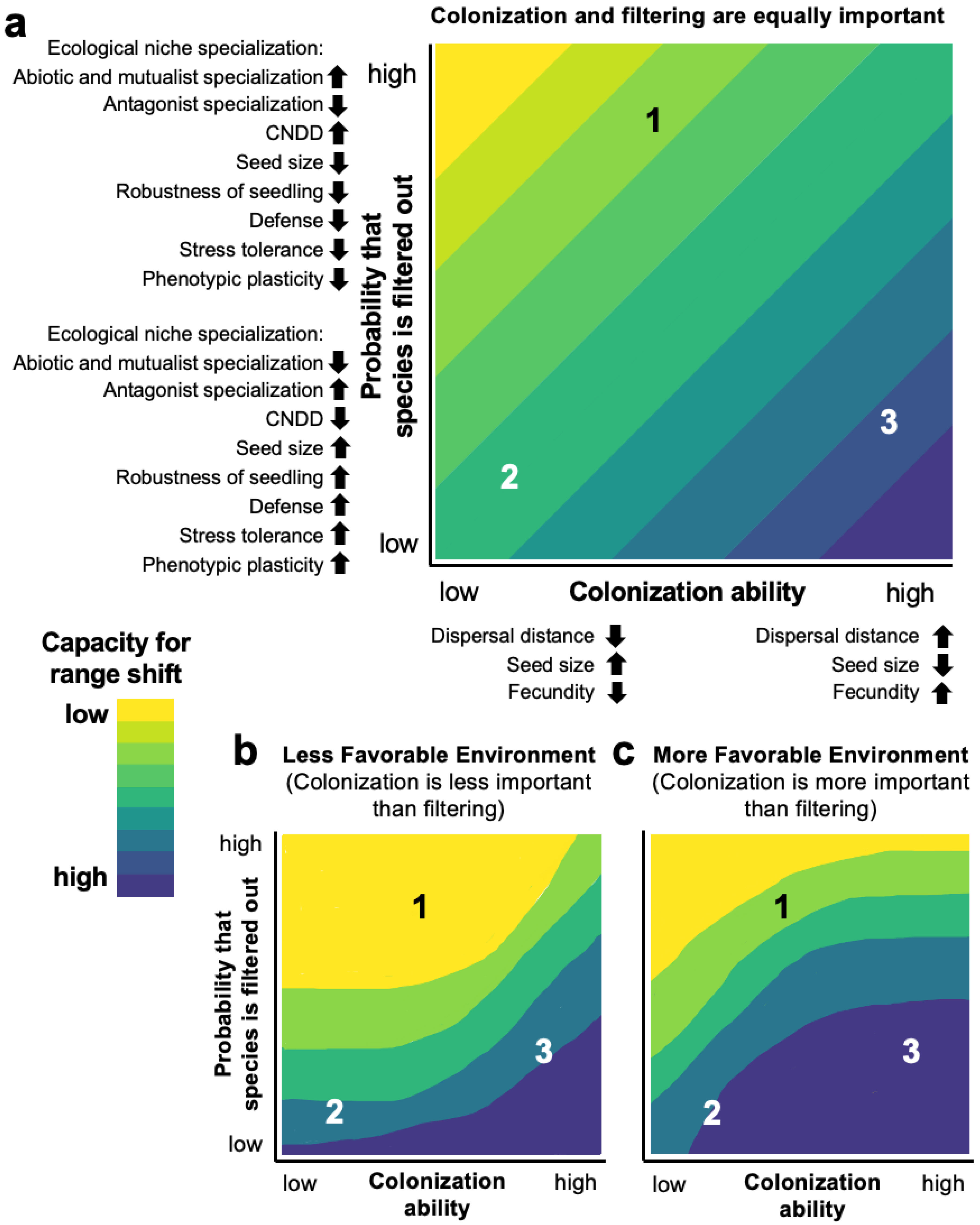{kind=link}
| Trade-Off | Combinations of Traits or Strategies | Effect on Habitat and Geographic Ranges | Citations |
|---|---|---|---|
| Growth–mortality trade-off (the fast-slow continuum) | Fast growth and short lifespan, or slow growth and long lifespan | Fast-growing individuals would be favored when colonization ability is more limiting and rapid climate shifts could favor a shorter lifespan (i.e., generation time) | [96,101,129,130,131,132] |
| Growth–defense trade-off | Greater investment in defensive compounds and/or carbohydrate storage, or in growth | Well-defended individuals with more carbohydrates (greater stress tolerance) should be less likely to be filtered out of a site than fast growers when habitat filtering is more limiting | [73,130,133,134,135] |
| Growth–maturation trade-off (or stature–recruitment trade-off) | Greater investment in sapling and adult stem growth and survival, or in reproduction at a younger age and smaller size (greater seedling recruitment and more years of reproduction) | Individuals with higher adult and sapling growth and survival should be less likely to be filtered out; otherwise, higher reproductive investment may be more beneficial | [129,136,137] |
| Seed size–seed number trade-off | Produce and disperse many small seeds or few large seeds | Smaller seed size and higher fecundity should increase colonization ability when colonization is more limiting, increasing capacity for range shift | [106,107,109,138] |
| Colonization–competition trade-off | Better colonizer (early arrival, fast growth, high fecundity); or better competitor (late arrival, high survival, low fecundity) | High fecundity may increase range shift capacity through increased colonization ability, but high survival may more effectively reduce probability of being filtered out | [58,116,139,140,141] |
| Tolerance–fecundity trade-off | Produce more, less stress tolerant seeds (better colonizer), or produce fewer, more stress-tolerant seeds (better competitor) | High fecundity may increase range shift capacity through increased colonization ability, but higher stress tolerance may reduce probability of being filtered out | [142,143,144,145] |
| Specialist–generalist trade-off (low or high ecological niche specialization) | Express ecological niche specialization (i.e., in abiotic conditions and relationships with mutualists and/or antagonists), or exhibit low ecological niche specialization (i.e., wider range of abiotic conditions and generalist relationships with mutualists and/or antagonists) | Higher specialization in abiotic conditions and/or with mutualists should increase the probability of being filtered out of a site; higher specialization with antagonists may decrease probability of being filtered out through reduced conspecific negative density-dependence | [85,146,147,148] |
| Acquisitive–conservative spectrum (leaf economics spectrum, wood economics spectrum) | Build low-cost, short-lived, thin leaves, and low-density, low- fiber wood with small carbohydrate storage; or high-cost, long-lived, thick leaves, and high-density, high-fiber wood with large carbohydrate storage | Acquisitive traits may increase an individual’s probability of being filtered out of a site when habitat filtering is more limiting, reducing capacity for range shifts | [103,104,149,150,151] |
| High versus low phenotypic plasticity | Express highly plastic, more costly phenotype, or less plastic, less costly phenotype | Individuals with high phenotypic plasticity should be less likely to be filtered out of a site than less plastic individuals when habitat filtering is more limiting | [29,42,43,152,153,154,155] |
5. Phenotype–Environment Matching
5.1. Abiotic Environment
5.2. Biotic Environment
5.3. Phenotypic Plasticity
6. Plant Species’ Capacity for Range Shifts
7. Synthesis and Future Directions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Glossary
| adaptation, adaptive evolution | a heritable change in a genetically determined phenotypic trait that results in higher fitness in a particular environment A |
| Allee effect | a form of positive density dependence; a positive relationship between population fitness and population density B,C |
| colonization ability | the capacity to arrive at a novel site via the dispersal of propagules D,E |
| conspecific negative density-dependence | decrease in individual-level components of fitness (i.e., growth, survival, reproduction) with increasing conspecific population densities, often due to host-specific natural enemies (e.g., pathogens, herbivores) and intraspecific competition for shared resources F,G. |
| dispersal limitation | factor(s) that impede a species from reaching suitable sites for recruitment H,I |
| ecological niche | the set of abiotic and biotic environmental conditions, including resources, mutualists, and antagonists, allowing a species or population to maintain positive population growth J,K |
| ecological strategy | attributes (e.g., phenotypic traits) from molecular to whole-organism scales that describe how individuals acquire and allocate resources to growth, survival, and reproduction, which may vary inter- and intraspecifically L,M |
| fundamental niche | a largely theoretical construct defining the complete set of abiotic environmental conditions under which a species can maintain positive population growth in the absence of biotic interactions K. |
| geographic range | the boundary defining the geographic area encompassing the complete set of locations where a species occurs N,O |
| habitat filtering | the abiotic and/or biotic factors preventing the establishment and persistence of individuals of a particular species at a particular site or in a particular habitat 16 P |
| habitat range | the local (i.e., small spatial scale) spatial distribution of populations of a species Q,R,S. |
| metapopulation and source-sink dynamics | a network of many discrete populations connected by dispersal, including high-quality habitat patches (sources) where population birth rates are greater than death rates, allowing for emigration to other patches, and low-quality habitat patches (sinks) where population death rates are greater than birth rates and therefore require immigration in order to be maintained T,U |
| phenotype-environment matching | maximizing fitness by expressing the optimal phenotype for the local environment V,W. |
| phenotypic integration | the coordination of the expression of functionally related traits within an individual that constrains trait variation to values that maximize fitness, given the values of all other traits 24 X. |
| phenotypic plasticity | the ability of a single genotype to express different phenotypes in different environments Y,Z |
| range shift | movement of a population into new, potentially more suitable, areas at the habitat or geographic range scale N,AA |
| realized niche | the set of abiotic environmental conditions under which a species can maintain positive population growth, after accounting for the effects of interspecific interactions (i.e., antagonists and mutualists) K,U |
| taxon cycle | phases of a species’ geographic range expansion and contraction that arise from the evolution of ecological strategies and phenotypes that favor expansion, followed by ecologically and evolutionarily driven shifts to a phenotype that promotes range contraction BB,CC |
| vital rates | the rates of growth, survival, and reproduction of individuals DD |
| Glossary References: A [208]; B [352]; C [300]; D [58]; E [59]; F [285]; G [286]; H [353]; I [65]; J [23]; K [22]; L [94]; M [354]; N [27]; O [28]; P [200]; Q [24]; R [26]; S [25]; T [48]; U [51]; V [60]; W [61]; X [308]; Y [43]; Z [44]; AA [9]; BB [91]; CC [93]; DD [355]. | |
References
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Weiskopf, S.R.; Rubenstein, M.A.; Crozier, L.G.; Gaichas, S.; Griffis, R.; Halofsky, J.E.; Hyde, K.J.W.; Morelli, T.L.; Morisette, J.T.; Muñoz, R.C.; et al. Climate change effects on biodiversity, ecosystems, ecosystem services, and natural resource management in the United States. Sci. Total Environ. 2020, 733, 137782. [Google Scholar] [CrossRef] [PubMed]
- Wiens, J.J. Climate-Related Local Extinctions Are Already Widespread among Plant and Animal Species. PLoS Biol. 2016, 14, e2001104. [Google Scholar] [CrossRef] [Green Version]
- Morueta-Holme, N.; Enquist, B.J.; McGill, B.J.; Boyle, B.; Jørgensen, P.M.; Ott, J.E.; Peet, R.K.; Šímová, I.; Sloat, L.L.; Thiers, B.; et al. Habitat area and climate stability determine geographical variation in plant species range sizes. Ecol. Lett. 2013, 16, 1446–1454. [Google Scholar] [CrossRef]
- Diffenbaugh, N.S.; Field, C.B. Changes in Ecologically Critical Terrestrial Climate Conditions. Science 2013, 341, 486–492. [Google Scholar] [CrossRef] [Green Version]
- Gougherty, A.V.; Keller, S.R.; Fitzpatrick, M.C. Maladaptation, migration and extirpation fuel climate change risk in a forest tree species. Nat. Clim. Chang. 2021, 11, 166–171. [Google Scholar] [CrossRef]
- Williams, J.W.; Jackson, S.T. Novel climates, no-analog communities, and ecological surprises. Front. Ecol. Environ. 2007, 5, 475–482. [Google Scholar] [CrossRef]
- Jackson, S.T.; Overpeck, J.T. Responses of plant populations and communities to environmental changes of the late Quaternary. Paleobiology 2000, 26, 28. [Google Scholar] [CrossRef]
- Davis, M.B.; Shaw, R.G. Range Shifts and Adaptive Responses to Quaternary Climate Change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Savolainen, O.; Pyhäjärvi, T.; Knürr, T. Gene Flow and Local Adaptation in Trees. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 595–619. [Google Scholar] [CrossRef]
- McLachlan, J.S.; Clark, J.S.; Manos, P.S. Molecular indicators of tree migration capacity under rapid climate change. Ecology 2005, 86, 2088–2098. [Google Scholar] [CrossRef] [Green Version]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef]
- Birks, H.J.B. Holocene Isochrone Maps and Patterns of Tree-Spreading in the British Isles. J. Biogeogr. 1989, 16, 503. [Google Scholar] [CrossRef]
- Clark, J.S.; Fastie, C.; Hurtt, G.; Jackson, S.T.; Johnson, C.; King, G.A.; Lewis, M.; Lynch, J.; Pacala, S.; Prentice, C.; et al. Reid’s Paradox of Rapid Plant Migration. BioScience 1998, 48, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Gear, A.J.; Huntley, B. Rapid Changes in the Range Limits of Scots Pine 4000 Years Ago. Science 1991, 251, 544–547. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.B. Lags in vegetation response to greenhouse warming. Clim. Chang. 1989, 15, 75–82. [Google Scholar] [CrossRef]
- Iverson, L.R.; Schwartz, M.W.; Prasad, A.M. How fast and far might tree species migrate in the eastern United States due to climate change?: Migration of tree species due to climate change. Glob. Ecol. Biogeogr. 2004, 13, 209–219. [Google Scholar] [CrossRef]
- Scheller, R.; Mladenoff, D. Simulated effects of climate change, fragmentation, and inter-specific competition on tree species migration in northern Wisconsin, USA. Clim. Res. 2008, 36, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Andrus, R.; Bergeron, Y.; Bogdziewicz, M.; Bragg, D.C.; Brockway, D.; Cleavitt, N.L.; Courbaud, B.; Das, A.J.; Dietze, M.; et al. North American tree migration paced by climate in the West, lagging in the East. Proc. Natl. Acad. Sci. USA 2022, 119, e2116691118. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological Niches and Geographic Distributions; MPB-49; Princeton University Press: Princeton, NJ, USA, 2012. [Google Scholar]
- Hutchinson, G.E. Concluding remarks. Quant. Biol. 1957, 22, 415–427. [Google Scholar] [CrossRef]
- Chase, J.M.; Leibold, M.A. Ecological Niches: Linking Classical and Contemporary Approaches; University of Chicago Press: Chicago, IL, USA, 2003. [Google Scholar]
- Brown, J.H. On the Relationship between Abundance and Distribution of Species. Am. Nat. 1984, 124, 255–279. [Google Scholar] [CrossRef]
- Sexton, J.P.; Dickman, E.E. What can local and geographic population limits tell us about distributions? Am. J. Bot. 2016, 103, 129–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaston, K.J. Geographic range limits: Achieving synthesis. Proc. R. Soc. B Biol. Sci. 2009, 276, 1395–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.H.; Stevens, G.C.; Kaufman, D.M. The Geographic Range: Size, Shape, Boundaries, and Internal Structure. Annu. Rev. Ecol. Syst. 1996, 27, 597–623. [Google Scholar] [CrossRef] [Green Version]
- Gaston, K.J. Geographic range limits of species. Proc. R. Soc. B Biol. Sci. 2009, 276, 1391–1393. [Google Scholar] [CrossRef]
- Edelaar, P.; Jovani, R.; Gomez-Mestre, I. Should I Change or Should I Go? Phenotypic Plasticity and Matching Habitat Choice in the Adaptation to Environmental Heterogeneity. Am. Nat. 2017, 190, 506–520. [Google Scholar] [CrossRef]
- Paul, J.R.; Sheth, S.N.; Angert, A.L. Quantifying the Impact of Gene Flow on Phenotype-Environment Mismatch: A Demonstration with the Scarlet Monkeyflower Mimulus cardinalis. Am. Nat. 2011, 178, S62–S79. [Google Scholar] [CrossRef] [Green Version]
- Baldeck, C.A.; Harms, K.E.; Yavitt, J.B.; John, R.; Turner, B.L.; Valencia, R.; Navarrete, H.; Davies, S.J.; Chuyong, G.B.; Kenfack, D.; et al. Soil resources and topography shape local tree community structure in tropical forests. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122532. [Google Scholar] [CrossRef]
- Baltzer, J.L.; Davies, S.J. Rainfall seasonality and pest pressure as determinants of tropical tree species’ distributions. Ecol. Evol. 2012, 2, 2682–2694. [Google Scholar] [CrossRef]
- Hamann, E.; Denney, D.; Day, S.; Lombardi, E.; Jameel, M.I.; MacTavish, R.; Anderson, J.T. Review: Plant eco-evolutionary responses to climate change: Emerging directions. Plant Sci. 2021, 304, 110737. [Google Scholar] [CrossRef]
- Moorcroft, P.R.; Pacala, S.W.; Lewis, M.A. Potential role of natural enemies during tree range expansions following climate change. J. Theor. Biol. 2006, 241, 601–616. [Google Scholar] [CrossRef]
- Canham, C.D.; Murphy, L. The demography of tree species response to climate: Seedling recruitment and survival. Ecosphere 2016, 7, e01424. [Google Scholar] [CrossRef]
- Higgins, S.I.; Clark, J.S.; Nathan, R.; Hovestadt, T.; Schurr, F.; Fragoso, J.M.V.; Aguiar, M.R.; Ribbens, E.; Lavorel, S. Forecasting plant migration rates: Managing uncertainty for risk assessment. J. Ecol. 2003, 91, 341–347. [Google Scholar] [CrossRef] [Green Version]
- Nathan, R.; Casagrandi, R. A simple mechanistic model of seed dispersal, predation and plant establishment: Janzen-Connell and beyond. J. Ecol. 2004, 92, 733–746. [Google Scholar] [CrossRef]
- Purves, D.W. The demography of range boundaries versus range cores in eastern US tree species. Proc. R. Soc. B Biol. Sci. 2009, 276, 1477–1484. [Google Scholar] [CrossRef] [Green Version]
- Corlett, R.T.; Westcott, D.A. Will plant movements keep up with climate change? Trends Ecol. Evol. 2013, 28, 482–488. [Google Scholar] [CrossRef]
- Merilä, J.; Hendry, A.P. Climate change, adaptation, and phenotypic plasticity: The problem and the evidence. Evol. Appl. 2014, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jump, A.S.; Peñuelas, J. Running to stand still: Adaptation and the response of plants to rapid climate change. Ecol. Lett. 2005, 8, 1010–1020. [Google Scholar] [CrossRef]
- Padilla, D.K.; Adolph, S.C. Plastic inducible morphologies are not always adaptive: The importance of time delays in a stochastic environment. Evol. Ecol. 1996, 10, 105–117. [Google Scholar] [CrossRef]
- Schlichting, C.D. The Evolution of Phenotypic Plasticity in Plants. Annu. Rev. Ecol. Syst. 1986, 17, 28. [Google Scholar] [CrossRef]
- Whitman, D.; Agrawal, A. What is Phenotypic Plasticity and Why is it Important? In Phenotypic Plasticity of Insects; Whitman, D., Ananthakrishnan, T., Eds.; CRC Press: Boca Raton, FL, USA, 2009; ISBN 978-1-57808-423-4. [Google Scholar]
- Bradshaw, A.D. Evolutionary Significance of Phenotypic Plasticity in Plants. Adv. Genet. 1965, 13, 115–155. [Google Scholar] [CrossRef]
- Harrison, S.P.; Sanchez Goñi, M.F. Global patterns of vegetation response to millennial-scale variability and rapid climate change during the last glacial period. Quat. Sci. Rev. 2010, 29, 2957–2980. [Google Scholar] [CrossRef]
- Lalejini, A.; Ferguson, A.J.; Grant, N.A.; Ofria, C. Adaptive Phenotypic Plasticity Stabilizes Evolution in Fluctuating Environments. Front. Ecol. Evol. 2021, 9, 715381. [Google Scholar] [CrossRef]
- Hanski, I. Metapopulation dynamics. Nature 1998, 396, 41–49. [Google Scholar] [CrossRef]
- Hanski, I.; Mononen, T.; Ovaskainen, O. Eco-Evolutionary Metapopulation Dynamics and the Spatial Scale of Adaptation. Am. Nat. 2011, 177, 29–43. [Google Scholar] [CrossRef] [Green Version]
- Negrín Dastis, J.O.; Milne, R.; Guichard, F.; Derry, A.M. Phenotype–environment mismatch in metapopulations—Implications for the maintenance of maladaptation at the regional scale. Evol. Appl. 2019, 12, 1475–1486. [Google Scholar] [CrossRef]
- Pulliam, H.R. Sources, Sinks, and Population Regulation. Am. Nat. 1988, 132, 652–661. [Google Scholar] [CrossRef]
- Van Horne, B. Density as a Misleading Indicator of Habitat Quality. J. Wildl. Manag. 1983, 47, 893. [Google Scholar] [CrossRef]
- Abrams, M.D.; Copenheaver, C.A.; Black, B.A.; van de Gevel, S. Dendroecology and climatic impacts for a relict, old-growth, bog forest in the Ridge and Valley Province of central Pennsylvania, U.S.A. Can. J. Bot. 2001, 79, 58–69. [Google Scholar]
- Chesson, P. Multispecies competition in variable environments. Theor. Popul. Biol. 1994, 45, 227–276. [Google Scholar] [CrossRef]
- Hampe, A.; Jump, A.S. Climate Relicts: Past, Present, Future. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 313–333. [Google Scholar] [CrossRef] [Green Version]
- Kaul, R.B.; Kantak, G.E.; Churchill, S.P. The Niobrara River Valley, a Postglacial Migration Corridor and Refugium of Forest Plants and Animals in the Grasslands of Central North America. Bot. Rev. 1988, 54, 44–81. [Google Scholar] [CrossRef]
- Schupp, E.W.; Howe, H.F.; Augspurger, C.K.; Levey, D.J. Arrival and Survival in Tropical Treefall Gaps. Ecology 1989, 70, 562–564. [Google Scholar] [CrossRef]
- Levins, R.; Culver, D. Regional Coexistence of Species and Competition between Rare Species. Proc. Natl. Acad. Sci. USA 1971, 68, 1246–1248. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.W.; Wilson, H.B. The Competition-Colonization Trade-off Is Dead; Long Live the Competition-Colonization Trade-off. Am. Nat. 2001, 158, 49–63. [Google Scholar] [CrossRef]
- DeWitt, T.J.; Sih, A.; Wilson, D.S. Costs and limits of phenotypic plasticity. Trends Ecol. Evol. 1998, 13, 77–81. [Google Scholar] [CrossRef]
- Levins, R. Evolution in Changing Environments: Some Theoretical Explorations; Monographs in Population Biology; Princeton University Press: Princeton, NJ, USA, 1968. [Google Scholar]
- Grinnell, J. The Niche-Relationships of the California Thrasher. Auk 1917, 34, 427–433. [Google Scholar] [CrossRef]
- Elton, C.S. Animal Ecology; University of Chicago Press: Chicago, IL, USA, 1927. [Google Scholar]
- Ackerly, D.D. Community Assembly, Niche Conservatism, and Adaptive Evolution in Changing Environments. Int. J. Plant Sci. 2003, 164, S165–S184. [Google Scholar] [CrossRef]
- Pulliam, H.R. On the relationship between niche and distribution. Ecol. Lett. 2000, 3, 349–361. [Google Scholar] [CrossRef]
- Gaston, K.J. How Large Is a Species’ Geographic Range? Oikos 1991, 61, 434. [Google Scholar] [CrossRef]
- Gaston, K.J. Species-range size distributions: Products of speciation, extinction and transformation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1998, 353, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Angert, A.L. The niche, limits to species’ distributions, and spatiotemporal variation in demography across the elevation ranges of two monkeyflowers. Proc. Natl. Acad. Sci. USA 2009, 106, 19693–19698. [Google Scholar] [CrossRef] [Green Version]
- Holt, R.D.; Keitt, T.H.; Lewis, M.A.; Maurer, B.A.; Taper, M.L. Theoretical models of species’ borders: Single species approaches. Oikos 2005, 108, 18–27. [Google Scholar] [CrossRef]
- Watkinson, A.R. On the Abundance of Plants Along an Environmental Gradient. J. Ecol. 1985, 73, 569. [Google Scholar] [CrossRef]
- Holt, R.D. On the evolutionary ecology of species’ ranges. Evol. Ecol. Res. 2003, 5, 159–178. [Google Scholar]
- Desprez, J.; Iannone, B.V.; Yang, P.; Oswalt, C.M.; Fei, S. Northward migration under a changing climate: A case study of blackgum (Nyssa sylvatica). Clim. Chang. 2014, 126, 151–162. [Google Scholar] [CrossRef]
- Fine, P.V.A.; Miller, Z.J.; Mesones, I.; Irazuzta, S.; Appel, H.M.; Stevens, M.H.H.; Sääksjärvi, I.; Schultz, J.C.; Coley, P.D. The growth-defense trade-off and habitat specialization by plants in Amazonian forests. Ecology 2006, 87, S150–S162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortunel, C.; Paine, C.E.T.; Fine, P.V.A.; Mesones, I.; Goret, J.; Burban, B.; Cazal, J.; Baraloto, C. There’s no place like home: Seedling mortality contributes to the habitat specialisation of tree species across Amazonia. Ecol. Lett. 2016, 19, 1256–1266. [Google Scholar] [CrossRef] [PubMed]
- Case, T.J.; Taper, M.L. Interspecific Competition, Environmental Gradients, Gene Flow, and the Coevolution of Species’ Borders. Am. Nat. 2000, 155, 23. [Google Scholar] [CrossRef] [PubMed]
- Burrows, M.T.; Schoeman, D.S.; Richardson, A.J.; Molinos, J.G.; Hoffmann, A.; Buckley, L.B.; Moore, P.J.; Brown, C.J.; Bruno, J.F.; Duarte, C.M.; et al. Geographical limits to species-range shifts are suggested by climate velocity. Nature 2014, 507, 492–495. [Google Scholar] [CrossRef] [Green Version]
- Sheth, S.N.; Jiménez, I.; Angert, A.L. Identifying the paths leading to variation in geographical range size in western North American monkeyflowers. J. Biogeogr. 2014, 41, 2344–2356. [Google Scholar] [CrossRef]
- Saupe, E.E.; Myers, C.E.; Peterson, A.T.; Soberón, J.; Singarayer, J.; Valdes, P.; Qiao, H. Non-random latitudinal gradients in range size and niche breadth predicted by spatial patterns of climate. Glob. Ecol. Biogeogr. 2019, 28, 928–942. [Google Scholar] [CrossRef]
- Slatyer, R.A.; Hirst, M.; Sexton, J.P. Niche breadth predicts geographical range size: A general ecological pattern. Ecol. Lett. 2013, 16, 1104–1114. [Google Scholar] [CrossRef]
- Xu, W.; Svenning, J.-C.; Chen, G.; Chen, B.; Huang, J.; Ma, K. Plant geographical range size and climate stability in China: Growth form matters. Glob. Ecol. Biogeogr. 2018, 27, 506–517. [Google Scholar] [CrossRef]
- Seliger, B.J.; McGill, B.J.; Svenning, J.; Gill, J.L. Widespread underfilling of the potential ranges of North American trees. J. Biogeogr. 2021, 48, 359–371. [Google Scholar] [CrossRef]
- Shay, J.E.; Pennington, L.K.; Mandussi Montiel-Molina, J.A.; Toews, D.J.; Hendrickson, B.T.; Sexton, J.P. Rules of Plant Species Ranges: Applications for Conservation Strategies. Front. Ecol. Evol. 2021, 9, 700962. [Google Scholar] [CrossRef]
- Gaston, K.J. Species-range-size distributions: Patterns, mechanisms and implications. Trends Ecol. Evol. 1996, 11, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Waldron, A. Null Models of Geographic Range Size Evolution Reaffirm Its Heritability. Am. Nat. 2007, 170, 221–231. [Google Scholar] [CrossRef]
- Emery, N.C.; Forrestel, E.J.; Jui, G.; Park, M.S.; Baldwin, B.G.; Ackerly, D.D. Niche evolution across spatial scales: Climate and habitat specialization in California Lasthenia (Asteraceae). Ecology 2012, 93, S151–S166. [Google Scholar] [CrossRef]
- Kirkpatrick, M.; Barton, N.H. Evolution of a Species’ Range. Am. Nat. 1997, 150, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Leão, T.C.C.; Lughadha, E.N.; Reich, P.B. Evolutionary patterns in the geographic range size of Atlantic Forest plants. Ecography 2020, 43, 1510–1520. [Google Scholar] [CrossRef]
- Sheth, S.N.; Angert, A.L. The evolution of environmental tolerance and range size: A comparison of geographically restricted and widespread Mimulus. Evolution 2014, 68, 2917–2931. [Google Scholar] [CrossRef] [PubMed]
- Wiens, J.J. The niche, biogeography and species interactions. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2336–2350. [Google Scholar] [CrossRef] [Green Version]
- Webb, T.J.; Gaston, K.J. On the Heritability of Geographic Range Sizes. Am. Nat. 2003, 161, 553–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Economo, E.P.; Sarnat, E.M. Revisiting the Ants of Melanesia and the Taxon Cycle: Historical and Human-Mediated Invasions of a Tropical Archipelago. Am. Nat. 2012, 180, E1–E16. [Google Scholar] [CrossRef]
- Rull, V. Taxon Cycles in Neotropical Mangroves. Plants 2023, 12, 244. [Google Scholar] [CrossRef]
- Wilson, E.O. The Nature of the Taxon Cycle in the Melanesian Ant Fauna. Am. Nat. 1961, 95, 169–193. [Google Scholar] [CrossRef]
- Grime, J.P. Vegetation classification by reference to strategies. Nature 1974, 250, 26–31. [Google Scholar] [CrossRef]
- Loehle, C. Strategy Space and the Disturbance Spectrum: A Life-History Model for Tree Species Coexistence. Am. Nat. 2000, 156, 14–33. [Google Scholar] [CrossRef]
- Reich, P.B. The world-wide ‘fast-slow’ plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Salguero-Gómez, R.; Jones, O.R.; Jongejans, E.; Blomberg, S.P.; Hodgson, D.J.; Mbeau-Ache, C.; Zuidema, P.A.; de Kroon, H.; Buckley, Y.M. Fast–slow continuum and reproductive strategies structure plant life-history variation worldwide. Proc. Natl. Acad. Sci. USA 2016, 113, 230–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westoby, M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant Soil 1998, 199, 213–227. [Google Scholar] [CrossRef]
- Gianoli, E.; Palacio-López, K. Phenotypic integration may constrain phenotypic plasticity in plants. Oikos 2009, 118, 1924–1928. [Google Scholar] [CrossRef]
- Matesanz, S.; Blanco-Sánchez, M.; Ramos-Muñoz, M.; Cruz, M.; Benavides, R.; Escudero, A. Phenotypic integration does not constrain phenotypic plasticity: Differential plasticity of traits is associated to their integration across environments. New Phytol. 2021, 231, 2359–2370. [Google Scholar] [CrossRef]
- Russo, S.E.; McMahon, S.M.; Detto, M.; Ledder, G.; Wright, S.J.; Condit, R.S.; Davies, S.J.; Ashton, P.S.; Bunyavejchewin, S.; Chang-Yang, C.-H.; et al. The interspecific growth–mortality trade-off is not a general framework for tropical forest community structure. Nat. Ecol. Evol. 2021, 5, 174–183. [Google Scholar] [CrossRef]
- Russo, S.E.; Ledder, G.; Muller, E.B.; Nisbet, R.M. Dynamic Energy Budget models: Fertile ground for understanding resource allocation in plants in a changing world. Conserv. Physiol. 2022, 10, coac061. [Google Scholar] [CrossRef]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.C.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Colin Prentice, I.; et al. The global spectrum of plant form and function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Russo, S.E.; Davies, S.J.; King, D.A.; Tan, S. Soil-related performance variation and distributions of tree species in a Bornean rain forest. J. Ecol. 2005, 93, 879–889. [Google Scholar] [CrossRef] [Green Version]
- Greene, D.F.; Johnson, E.A. Estimating the Mean Annual Seed Production of Trees. Ecology 1994, 75, 642–647. [Google Scholar] [CrossRef] [Green Version]
- Henery, M.L.; Westoby, M. Seed mass and seed nutrient content as predictors of seed output variation between species. Oikos 2001, 92, 479–490. [Google Scholar] [CrossRef]
- Moles, A.T.; Westoby, M. Seedling survival and seed size: A synthesis of the literature. J. Ecol. 2004, 92, 372–383. [Google Scholar] [CrossRef]
- Shipley, B.; Dion, J. The Allometry of Seed Production in Herbaceous Angiosperms. Am. Nat. 1992, 139, 467–483. [Google Scholar] [CrossRef]
- Wenny, D.G. Advantages of seed dispersal: A re-evaluation of directed dispersal. Evol. Ecol. Res. 2001, 3, 24. [Google Scholar]
- Chapman, C.A.; Russo, S.E. Primate seed dispersal: Linking behavioral ecology with forest community structure. In Primates in Perspective; Oxford University Press: London, UK, 2006; pp. 510–525. [Google Scholar]
- Russo, S.E.; Portnoy, S.; Augspurger, C.K. Incorporating animal behavior into seed dispersal models: Implications for seed shadows. Ecology 2006, 87, 3160–3174. [Google Scholar] [CrossRef] [Green Version]
- Butler, D.W.; Green, R.J.; Lamb, D.; McDonald, W.J.F.; Forster, P.I. Biogeography of seed-dispersal syndromes, life-forms and seed sizes among woody rain-forest plants in Australia’s subtropics. J. Biogeogr. 2007, 34, 1736–1750. [Google Scholar] [CrossRef]
- Howe, H.F. Making dispersal syndromes and networks useful in tropical conservation and restoration. Glob. Ecol. Conserv. 2016, 6, 152–178. [Google Scholar] [CrossRef] [Green Version]
- Albert, C.H.; Thuiller, W.; Yoccoz, N.G.; Douzet, R.; Aubert, S.; Lavorel, S. A multi-trait approach reveals the structure and the relative importance of intra- vs. interspecific variability in plant traits: Intra- vs. interspecific variability in plant traits. Funct. Ecol. 2010, 24, 1192–1201. [Google Scholar] [CrossRef]
- Bin, Y.; Lin, G.; Russo, S.E.; Huang, Z.; Shen, Y.; Cao, H.; Lian, J.; Ye, W. Testing the competition-colonization trade-off and its correlations with functional trait variations among subtropical tree species. Sci. Rep. 2019, 9, 14942. [Google Scholar] [CrossRef] [Green Version]
- Muller-Landau, H.C.; Wright, S.J.; Calderón, O.; Hubbell, S.P.; Foster, R.B. Assessing recruitment limitation: Concepts, methods and case-studies from a tropical forest. In Proceedings of the Seed Dispersal and Frugivory: Ecology, Evolution and Conservation. Third International Symposium-Workshop on Frugivores and Seed Dispersal, São Pedro, Brazil, 6–11 August 2000; Levey, D.J., Silva, W.R., Galetti, M., Eds.; CABI: Wallingford, UK, 2002; pp. 35–53, ISBN 978-0-85199-525-0. [Google Scholar]
- Schupp, E.W.; Milleron, T.; Russo, S.E. Dissemination limitation and the origin and maintenance of species-rich tropical forests. In Proceedings of the Seed Dispersal and Frugivory: Ecology, Evolution and Conservation. Third International Symposium-Workshop on Frugivores and Seed Dispersal, São Pedro, Brazil, 6–11 August 2000; Levey, D.J., Silva, W.R., Galetti, M., Eds.; CABI Publishing: Wallingford, UK, 2002; pp. 19–33, ISBN 978-0-85199-525-0. [Google Scholar]
- Svenning, J.-C.; Normand, S.; Skov, F. Postglacial dispersal limitation of widespread forest plant species in nemoral Europe. Ecography 2008, 31, 316–326. [Google Scholar] [CrossRef]
- Brodie, J.F.; Lieberman, S.; Moehrenschlager, A.; Redford, K.H.; Rodríguez, J.P.; Schwartz, M.; Seddon, P.J.; Watson, J.E.M. Global policy for assisted colonization of species. Science 2021, 372, 456–458. [Google Scholar] [CrossRef] [PubMed]
- McLACHLAN, J.S.; Hellmann, J.J.; Schwartz, M.W. A Framework for Debate of Assisted Migration in an Era of Climate Change. Conserv. Biol. 2007, 21, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Oldfather, M.F.; Van Den Elzen, C.L.; Heffernan, P.M.; Emery, N.C. Dispersal evolution in temporally variable environments: Implications for plant range dynamics. Am. J. Bot. 2021, 108, 1584–1594. [Google Scholar] [CrossRef]
- Clark, J.S. Why Trees Migrate So Fast: Confronting Theory with Dispersal Biology and the Paleorecord. Am. Nat. 1998, 152, 21. [Google Scholar] [CrossRef]
- Dick, C.W.; Bermingham, E.; Lemes, M.R.; Gribel, R. Extreme long-distance dispersal of the lowland tropical rainforest tree Ceiba pentandra L. (Malvaceae) in Africa and the Neotropics. Mol. Ecol. 2007, 16, 3039–3049. [Google Scholar] [CrossRef]
- Kremer, A.; Ronce, O.; Robledo-Arnuncio, J.J.; Guillaume, F.; Bohrer, G.; Nathan, R.; Bridle, J.R.; Gomulkiewicz, R.; Klein, E.K.; Ritland, K.; et al. Long-distance gene flow and adaptation of forest trees to rapid climate change. Ecol. Lett. 2012, 15, 378–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soons, M.B.; Bullock, J.M. Non-random seed abscission, long-distance wind dispersal and plant migration rates. J. Ecol. 2008, 96, 581–590. [Google Scholar] [CrossRef]
- Cain, M.L.; Damman, H.; Muir, A. Seed Dispersal and the Holocene Migration of Woodland Herbs. Ecol. Monogr. 1998, 68, 325–347. [Google Scholar] [CrossRef]
- Howe, H.F.; Smallwood, J. Ecology of Seed Dispersal. Annu. Rev. Ecol. Syst. 1982, 13, 201–228. [Google Scholar] [CrossRef]
- Detto, M.; Levine, J.M.; Pacala, S.W. Maintenance of high diversity in mechanistic forest dynamics models of competition for light. Ecol. Monogr. 2022, 92, e1500. [Google Scholar] [CrossRef]
- Loehle, C. Tree life history strategies: The role of defenses. Can. J. For. Res. 1988, 18, 209–222. [Google Scholar] [CrossRef]
- Russo, S.E.; Brown, P.; Tan, S.; Davies, S.J. Interspecific demographic trade-offs and soil-related habitat associations of tree species along resource gradients: Demographic trade-offs on habitat gradients. J. Ecol. 2008, 96, 192–203. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.J.; Kitajima, K.; Kraft, N.J.B.; Reich, P.B.; Wright, I.J.; Bunker, D.E.; Condit, R.; Dalling, J.W.; Davies, S.J.; Engelbrecht, B.M.J.; et al. Functional traits and the growth–mortality trade-off in tropical trees. Ecology 2010, 91, 3664–3674. [Google Scholar] [CrossRef] [PubMed]
- Coley, P.D.; Bryant, J.P.; Chapin, F.S. Resource Availability and Plant Antiherbivore Defense. Science 1985, 230, 895–899. [Google Scholar] [CrossRef] [Green Version]
- Herms, D.A.; Mattson, W.J. The Dilemma of Plants: To Grow or Defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar] [CrossRef] [Green Version]
- Huot, B.; Yao, J.; Montgomery, B.L.; He, S.Y. Growth–Defense Tradeoffs in Plants: A Balancing Act to Optimize Fitness. Mol. Plant 2014, 7, 1267–1287. [Google Scholar] [CrossRef] [Green Version]
- Kohyama, T. Size-Structured Tree Populations in Gap-Dynamic Forest--The Forest Architecture Hypothesis for the Stable Coexistence of Species. J. Ecol. 1993, 81, 131. [Google Scholar] [CrossRef]
- Rüger, N.; Comita, L.S.; Condit, R.; Purves, D.; Rosenbaum, B.; Visser, M.D.; Wright, S.J.; Wirth, C. Beyond the fast–slow continuum: Demographic dimensions structuring a tropical tree community. Ecol. Lett. 2018, 21, 1075–1084. [Google Scholar] [CrossRef] [Green Version]
- Moles, A.T.; Westoby, M. Seed size and plant strategy across the whole life cycle. Oikos 2006, 113, 91–105. [Google Scholar] [CrossRef]
- Calcagno, V.; Mouquet, N.; Jarne, P.; David, P. Coexistence in a metacommunity: The competition–colonization trade-off is not dead. Ecol. Lett. 2006, 9, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D. Competition and Biodiversity in Spatially Structured Habitats. Ecology 1994, 75, 2–16. [Google Scholar] [CrossRef]
- Hara, T. Effects of variation in individual growth on plant species coexistence. J. Veg. Sci. 1993, 4, 409–416. [Google Scholar] [CrossRef]
- D’Andrea, R.; O’Dwyer, J.P. Competition for space in a structured landscape: The effect of seed limitation on coexistence under a tolerance-fecundity trade-off. J. Ecol. 2021, 109, 1886–1897. [Google Scholar] [CrossRef]
- Lebrija-Trejos, E.; Reich, P.B.; Hernández, A.; Wright, S.J. Species with greater seed mass are more tolerant of conspecific neighbours: A key driver of early survival and future abundances in a tropical forest. Ecol. Lett. 2016, 19, 1071–1080. [Google Scholar] [CrossRef]
- Muller-Landau, H.C. The tolerance–fecundity trade-off and the maintenance of diversity in seed size. Proc. Natl. Acad. Sci. USA 2010, 107, 4242–4247. [Google Scholar] [CrossRef] [Green Version]
- Villellas, J.; García, M.B. The role of the tolerance-fecundity trade-off in maintaining intraspecific seed trait variation in a widespread dimorphic herb. Plant Biol. 2013, 15, 899–909. [Google Scholar] [CrossRef]
- Sexton, J.P.; Montiel, J.; Shay, J.E.; Stephens, M.R.; Slatyer, R.A. Evolution of Ecological Niche Breadth. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 183–206. [Google Scholar] [CrossRef] [Green Version]
- Futuyma, D.J.; Moreno, G. The evolution of ecological specialization. Annu. Rev. Ecol. Syst. 1988, 19, 207–233. [Google Scholar] [CrossRef]
- Peay, K.G. The Mutualistic Niche: Mycorrhizal Symbiosis and Community Dynamics. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 143–164. [Google Scholar] [CrossRef]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef]
- Kitajima, K.; Myers, J.A. Seedling ecophysiology: Strategies toward achievement of positive net carbon balance. In Seedling Ecology and Evolution; Leck, M.A., Parker, V.T., Simpson, R.L., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 172–188. ISBN 978-0-511-81513-3. [Google Scholar]
- Pierce, S.; Brusa, G.; Vagge, I.; Cerabolini, B.E.L. Allocating CSR plant functional types: The use of leaf economics and size traits to classify woody and herbaceous vascular plants. Funct. Ecol. 2013, 27, 1002–1010. [Google Scholar] [CrossRef] [Green Version]
- Donohue, K. Setting the Stage: Phenotypic Plasticity as Habitat Selection. Int. J. Plant Sci. 2003, 164, S79–S92. [Google Scholar] [CrossRef] [Green Version]
- Lande, R. Evolution of phenotypic plasticity in colonizing species. Mol. Ecol. 2015, 24, 2038–2045. [Google Scholar] [CrossRef]
- Murren, C.J.; Auld, J.R.; Callahan, H.; Ghalambor, C.K.; Handelsman, C.A.; Heskel, M.A.; Kingsolver, J.G.; Maclean, H.J.; Masel, J.; Maughan, H.; et al. Constraints on the evolution of phenotypic plasticity: Limits and costs of phenotype and plasticity. Heredity 2015, 115, 293–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zettlemoyer, M.A.; Peterson, M.L. Does Phenological Plasticity Help or Hinder Range Shifts Under Climate Change? Front. Ecol. Evol. 2021, 9, 689192. [Google Scholar] [CrossRef]
- Nathan, R.; Schurr, F.M.; Spiegel, O.; Steinitz, O.; Trakhtenbrot, A.; Tsoar, A. Mechanisms of long-distance seed dispersal. Trends Ecol. Evol. 2008, 23, 638–647. [Google Scholar] [CrossRef]
- Chen, S.; Poschlod, P.; Antonelli, A.; Liu, U.; Dickie, J.B. Trade-off between seed dispersal in space and time. Ecol. Lett. 2020, 23, 1635–1642. [Google Scholar] [CrossRef] [PubMed]
- Westoby, M.; Leishman, M.; Lord, J. Comparative ecology of seed size and dispersal. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1996, 351, 1309–1318. [Google Scholar]
- Leishman, M.R.; Westoby, M.; Jurado, E. Correlates of Seed Size Variation: A Comparison Among Five Temperate Floras. J. Ecol. 1995, 83, 517. [Google Scholar] [CrossRef]
- Moles, A.T.; Ackerly, D.D.; Webb, C.O.; Tweddle, J.C.; Dickie, J.B.; Westoby, M. A Brief History of Seed Size. Science 2005, 307, 576–580. [Google Scholar] [CrossRef] [Green Version]
- Westoby, M.; Jurado, E.; Leishman, M. Comparative evolutionary ecology of seed size. Trends Ecol. Evol. 1992, 7, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Leishman, M.R.; Wright, I.J.; Moles, A.T.; Westoby, M. The evolutionary ecology of seed size. In Seeds: The Ecology of Regeneration in Plant Communities; Fenner, M., Ed.; CABI Publishing: Wallingford, UK, 2000; pp. 31–57. ISBN 978-0-85199-947-0. [Google Scholar]
- Michaels, H.J.; Benner, B.; Hartgerink, A.P.; Lee, T.D.; Rice, S.; Willson, M.F.; Bertin, R.I. Seed size variation: Magnitude, distribution, and ecological correlates. Evol. Ecol. 1988, 2, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.S.; Macklin, E.; Wood, L. Stages and spatial scales of recruitment limitation in Southern Appalachian forests. Ecol. Monogr. 1998, 68, 213–235. [Google Scholar] [CrossRef]
- Clark, J.S.; Ji, Y. Fecundity and Dispersal in Plant Populations: Implications for Structure and Diversity. Am. Nat. 1995, 146, 72–111. [Google Scholar] [CrossRef]
- Leishman, M.R. Does the seed size/number trade-off model determine plant community structure? An assessment of the model mechanisms and their generality. Oikos 2001, 93, 294–302. [Google Scholar] [CrossRef]
- Venable, D.L. Size-Number Trade-Offs and the Variation of Seed Size with Plant Resource Status. Am. Nat. 1992, 140, 287–304. [Google Scholar] [CrossRef]
- Coomes, D.A.; Grubb, P.J. Colonization, tolerance, competition and seed-size variation within functional groups. Trends Ecol. Evol. 2003, 18, 283–291. [Google Scholar] [CrossRef]
- Jansen, P.A.; Bartholomeus, M.; Bongers, F.; Elzinga, J.A.; den Ouden, J.; van Wieren, S.E. The role of seed size in dispersal by a scatter-hoarding rodent. In Proceedings of the Seed Dispersal and Frugivory: Ecology, Evolution and Conservation. Third International Symposium-Workshop on Frugivores and Seed Dispersal, São Pedro, Brazil, 6–11 August 2000; Levey, D.J., Silva, W.R., Galetti, M., Eds.; CABI Publishing: Wallingford, UK, 2002; pp. 209–225, ISBN 978-0-85199-525-0. [Google Scholar]
- Dylewski, Ł.; Ortega, Y.K.; Bogdziewicz, M.; Pearson, D.E. Seed size predicts global effects of small mammal seed predation on plant recruitment. Ecol. Lett. 2020, 23, 1024–1033. [Google Scholar] [CrossRef]
- Moles, A.T.; Falster, D.S.; Leishman, M.R.; Westoby, M. Small-seeded species produce more seeds per square metre of canopy per year, but not per individual per lifetime: Seed mass and lifetime seed production. J. Ecol. 2004, 92, 384–396. [Google Scholar] [CrossRef]
- Calvo, R.N.; Horvitz, C.C. Pollinator Limitation, Cost of Reproduction, and Fitness in Plants: A Transition-Matrix Demographic Approach. Am. Nat. 1990, 136, 499–516. [Google Scholar] [CrossRef]
- Price, M.V.; Campbell, D.R.; Waser, N.M.; Brody, A.K. Bridging the generation gap in plants: Pollination, parental fecundity, and offspring demography. Ecology 2008, 89, 1596–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardee, G.L.; Jensen, I.O.; Inouye, D.W.; Irwin, R.E. The individual and combined effects of snowmelt timing and frost exposure on the reproductive success of montane forbs. J. Ecol. 2019, 107, 1970–1981. [Google Scholar] [CrossRef]
- Schiffer, A.; Loy, X.; Morozumi, C.; Brosi, B.J. Differences in individual flowering time change pollen limitation and seed set in three montane wildflowers. Am. J. Bot. 2022, e16123. [Google Scholar] [CrossRef] [PubMed]
- Ghazoul, J. Pollen and seed dispersal among dispersed plants. Biol. Rev. 2005, 80, 413. [Google Scholar] [CrossRef]
- Clark, J.S.; Silman, M.; Kern, R.; Macklin, E.; Hillerislambersi, J. Seed dispersal near and far: Patterns across temperate and tropical forests. Ecology 1999, 80, 1475–1494. [Google Scholar] [CrossRef]
- Howe, H.F.; Miriti, M.N. When Seed Dispersal Matters. BioScience 2004, 54, 651. [Google Scholar] [CrossRef]
- Russo, S.E. Responses of dispersal agents to tree and fruit traits in Virola calophylla (Myristicaceae): Implications for selection. Oecologia 2003, 136, 80–87. [Google Scholar] [CrossRef]
- Holbrook, K.M.; Smith, T.B. Seed dispersal and movement patterns in two species of Ceratogymna hornbills in a West African tropical lowland forest. Oecologia 2000, 125, 249–257. [Google Scholar] [CrossRef]
- Trakhtenbrot, A.; Nathan, R.; Perry, G.; Richardson, D.M. The importance of long-distance dispersal in biodiversity conservation: LDD in conservation. Divers. Distrib. 2005, 11, 173–181. [Google Scholar] [CrossRef]
- Willis, C.G.; Baskin, C.C.; Baskin, J.M.; Auld, J.R.; Venable, D.L.; Cavender-Bares, J.; Donohue, K.; de Casas, R.R.; The NESCent Germination Working Group. The evolution of seed dormancy: Environmental cues, evolutionary hubs, and diversification of the seed plants. New Phytol. 2014, 203, 300–309. [Google Scholar] [CrossRef]
- Baskin, C.C.; Chesson, P.L.; Baskin, J.M. Annual Seed Dormancy Cycles in Two Desert Winter Annuals. J. Ecol. 1993, 81, 551. [Google Scholar] [CrossRef]
- Pake, C.E.; Venable, D.L. Seed Banks in Desert Annuals: Implications for Persistence and Coexistence in Variable Environments. Ecology 1996, 77, 1427–1435. [Google Scholar] [CrossRef]
- Dalling, J.W.; Brown, T.A. Long-Term Persistence of Pioneer Species in Tropical Rain Forest Soil Seed Banks. Am. Nat. 2009, 173, 531–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalamea, P.; Dalling, J.W.; Sarmiento, C.; Arnold, A.E.; Delevich, C.; Berhow, M.A.; Ndobegang, A.; Gripenberg, S.; Davis, A.S. Dormancy-defense syndromes and tradeoffs between physical and chemical defenses in seeds of pioneer species. Ecology 2018, 99, 1988–1998. [Google Scholar] [CrossRef] [Green Version]
- Walck, J.L.; Hidayati, S.N.; Dixon, K.W.; Thompson, K.; Poschlod, P. Climate change and plant regeneration from seed: Climate change and plant regeneration. Glob. Chang. Biol. 2011, 17, 2145–2161. [Google Scholar] [CrossRef]
- Jankowska-Blaszczuk, M.; Daws, M.I. Impact of red: Far red ratios on germination of temperate forest herbs in relation to shade tolerance, seed mass and persistence in the soil. Funct. Ecol. 2007, 21, 1055–1062. [Google Scholar] [CrossRef]
- Rees, M. Evolutionary ecology of seed dormancy and seed size. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1996, 351, 1299–1308. [Google Scholar]
- Penfield, S. Seed dormancy and germination. Curr. Biol. 2017, 27, R874–R878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, E.S.; Lischke, H.; Schmatz, D.R.; Zimmermann, N.E. Climate, competition and connectivity affect future migration and ranges of European trees: Future migration and ranges of European trees. Glob. Ecol. Biogeogr. 2012, 21, 164–178. [Google Scholar] [CrossRef]
- Bonte, D.; Van Dyck, H.; Bullock, J.M.; Coulon, A.; Delgado, M.; Gibbs, M.; Lehouck, V.; Matthysen, E.; Mustin, K.; Saastamoinen, M.; et al. Costs of dispersal. Biol. Rev. 2012, 87, 290–312. [Google Scholar] [CrossRef]
- Thompson, K.; Hodgson, J.G.; Rich, T.C.G. Native and alien invasive plants: More of the same? Ecography 1995, 18, 390–402. [Google Scholar] [CrossRef]
- Hawkes, C.V. Are Invaders Moving Targets? The Generality and Persistence of Advantages in Size, Reproduction, and Enemy Release in Invasive Plant Species with Time since Introduction. Am. Nat. 2007, 170, 832–843. [Google Scholar] [CrossRef]
- Holthuijzen, A.M.A.; Sharik, T.L.; Fraser, J.D. Dispersal of eastern red cedar (Juniperus virginiana) into pastures: An overview. Can. J. Bot. 1987, 65, 1092–1095. [Google Scholar] [CrossRef]
- Grotkopp, E.; Rejmánek, M.; Rost, T.L. Toward a Causal Explanation of Plant Invasiveness: Seedling Growth and Life-History Strategies of 29 Pine (Pinus) Species. Am. Nat. 2002, 159, 396–419. [Google Scholar] [CrossRef] [PubMed]
- Divíšek, J.; Chytrý, M.; Beckage, B.; Gotelli, N.J.; Lososová, Z.; Pyšek, P.; Richardson, D.M.; Molofsky, J. Similarity of introduced plant species to native ones facilitates naturalization, but differences enhance invasion success. Nat. Commun. 2018, 9, 4631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, S.; Cabido, M.; Casanoves, F. Plant functional traits and environmental filters at a regional scale. J. Veg. Sci. 1998, 9, 113–122. [Google Scholar] [CrossRef]
- Laliberté, E.; Zemunik, G.; Turner, B.L. Environmental filtering explains variation in plant diversity along resource gradients. Science 2014, 345, 1602–1605. [Google Scholar] [CrossRef]
- Maire, V.; Gross, N.; Börger, L.; Proulx, R.; Wirth, C.; da Silveira Pontes, L.; Soussana, J.-F.; Louault, F. Habitat filtering and niche differentiation jointly explain species relative abundance within grassland communities along fertility and disturbance gradients. New Phytol. 2012, 196, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Sargent, R.D.; Ackerly, D.D. Plant–pollinator interactions and the assembly of plant communities. Trends Ecol. Evol. 2008, 23, 123–130. [Google Scholar] [CrossRef]
- Bazzaz, F.A. Habitat Selection in Plants. Am. Nat. 1991, 137, S116–S130. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Schwilk, D.W.; Ackerly, D.D. A trait-based test for habitat filtering: Convex hull volume. Ecology 2006, 87, 1465–1471. [Google Scholar] [CrossRef] [PubMed]
- Kraft, N.J.B.; Valencia, R.; Ackerly, D.D. Functional Traits and Niche-Based Tree Community Assembly in an Amazonian Forest. Science 2008, 322, 580–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraft, N.J.B.; Adler, P.B.; Godoy, O.; James, E.C.; Fuller, S.; Levine, J.M. Community assembly, coexistence and the environmental filtering metaphor. Funct. Ecol. 2015, 29, 592–599. [Google Scholar] [CrossRef]
- Weiher, E.; Clarke, G.D.P.; Keddy, P.A. Community Assembly Rules, Morphological Dispersion, and the Coexistence of Plant Species. Oikos 1998, 81, 309. [Google Scholar] [CrossRef]
- Sexton, J.P.; McIntyre, P.J.; Angert, A.L.; Rice, K.J. Evolution and Ecology of Species Range Limits. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 415–436. [Google Scholar] [CrossRef] [Green Version]
- Darwin, C. On the Origin of Species by Means of Natural Selection, or, the Preservation of Favoured Races in the Struggle for Life; J. Murray: London, UK, 1859. [Google Scholar]
- Morris, W.F.; Ehrlén, J.; Dahlgren, J.P.; Loomis, A.K.; Louthan, A.M. Biotic and anthropogenic forces rival climatic/abiotic factors in determining global plant population growth and fitness. Proc. Natl. Acad. Sci. USA 2020, 117, 1107–1112. [Google Scholar] [CrossRef] [Green Version]
- Funk, J.L.; Larson, J.E.; Ames, G.M.; Butterfield, B.J.; Cavender-Bares, J.; Firn, J.; Laughlin, D.C.; Sutton-Grier, A.E.; Williams, L.; Wright, J. Revisiting the Holy Grail: Using plant functional traits to understand ecological processes. Biol. Rev. 2017, 92, 1156–1173. [Google Scholar] [CrossRef]
- Lavorel, S.; Garnier, E. Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the Holy Grail: Plant response and effect groups. Funct. Ecol. 2002, 16, 545–556. [Google Scholar] [CrossRef]
- Baltzer, J.L.; Thomas, S.C. Leaf optical responses to light and soil nutrient availability in temperate deciduous trees. Am. J. Bot. 2005, 92, 214–223. [Google Scholar] [CrossRef]
- Libalah, M.B.; Droissart, V.; Sonké, B.; Hardy, O.J.; Drouet, T.; Pescador, D.S.; Kenfack, D.; Thomas, D.W.; Chuyong, G.B.; Couteron, P. Shift in functional traits along soil fertility gradient reflects non-random community assembly in a tropical African rainforest. Plant Ecol. Evol. 2017, 150, 265–278. [Google Scholar] [CrossRef] [Green Version]
- Russo, S.E.; Zhang, L.; Tan, S. Covariation between understorey light environments and soil resources in Bornean mixed dipterocarp rain forest. J. Trop. Ecol. 2012, 28, 33–44. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.D.; Vellend, M. Non-climatic constraints on upper elevational plant range expansion under climate change. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141779. [Google Scholar] [CrossRef] [Green Version]
- Graae, B.J.; Vandvik, V.; Armbruster, W.S.; Eiserhardt, W.L.; Svenning, J.-C.; Hylander, K.; Ehrlén, J.; Speed, J.D.M.; Klanderud, K.; Bråthen, K.A.; et al. Stay or go—How topographic complexity influences alpine plant population and community responses to climate change. Perspect. Plant Ecol. Evol. Syst. 2018, 30, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Solarik, K.A.; Messier, C.; Ouimet, R.; Bergeron, Y.; Gravel, D. Local adaptation of trees at the range margins impacts range shifts in the face of climate change. Glob. Ecol. Biogeogr. 2018, 27, 1507–1519. [Google Scholar] [CrossRef]
- Clark, D.B.; Palmer, M.W.; Clark, D.A. Edaphic factors and the landscape-scale distributions of tropical rain forest trees. Ecology 1999, 80, 2662–2675. [Google Scholar] [CrossRef]
- Thomson, D.M.; Baythavong, B.S.; Rice, K.J. Invasions and the Evolution of Range Limits. In Serpentine: The Evolution and Ecology of a Model System; University of California Press: Berkeley, CA, USA, 2011; Chapter 9; p. 20. [Google Scholar]
- Harrison, S.; Safford, H.D.; Grace, J.B.; Viers, J.H.; Davies, K.F. Regional and local species richness in an insular environment: Serpentine plants in California. Ecol. Monogr. 2006, 76, 41–56. [Google Scholar] [CrossRef] [Green Version]
- Davies, S.J.; Tan, S.; LaFrankie, J.V.; Potts, M.D. Soil-Related Floristic Variation in a Hyperdiverse Dipterocarp Forest. In Pollination Ecology and the Rain Forest; Roubik, D.W., Sakai, S., Hamid Karim, A.A., Eds.; Ecological Studies; Springer: New York, NY, USA, 2005; Volume 174, pp. 22–34. ISBN 978-0-387-21309-5. [Google Scholar]
- Zuleta, D.; Russo, S.E.; Barona, A.; Barreto-Silva, J.S.; Cardenas, D.; Castaño, N.; Davies, S.J.; Detto, M.; Sua, S.; Turner, B.L.; et al. Importance of topography for tree species habitat distributions in a terra firme forest in the Colombian Amazon. Plant Soil 2020, 450, 133–149. [Google Scholar] [CrossRef]
- Harms, K.E.; Condit, R.; Hubbell, S.P.; Foster, R.B. Habitat associations of trees and shrubs in a 50-ha neotropical forest plot: Habitat associations of trees and shrubs. J. Ecol. 2001, 89, 947–959. [Google Scholar] [CrossRef]
- Whitman, M.; Beaman, R.S.; Repin, R.; Kitayama, K.; Aiba, S.; Russo, S.E. Edaphic specialization and vegetation zones define elevational range-sizes for Mt Kinabalu regional flora. Ecography 2021, 44, 1698–1709. [Google Scholar] [CrossRef]
- Kenfack, D.; Chuyong, G.B.; Condit, R.; Russo, S.E.; Thomas, D.W. Demographic variation and habitat specialization of tree species in a diverse tropical forest of Cameroon. For. Ecosyst. 2014, 1, 22. [Google Scholar] [CrossRef] [Green Version]
- Foster, A.C.; Martin, P.H.; Redmond, M.D. Soil moisture strongly limits Douglas-fir seedling establishment near its upper elevational limit in the southern Rocky Mountains. Can. J. For. Res. 2020, 50, 837–842. [Google Scholar] [CrossRef]
- Moyes, A.B.; Castanha, C.; Germino, M.J.; Kueppers, L.M. Warming and the dependence of limber pine (Pinus flexilis) establishment on summer soil moisture within and above its current elevation range. Oecologia 2013, 171, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Cramer, M.D.; Hedding, D.W.; Greve, M.; Midgley, G.F.; Ripley, B.S. Plant specialisation may limit climate-induced vegetation change to within topographic and edaphic niches on a sub-Antarctic island. Funct. Ecol. 2022, 36, 2636–2648. [Google Scholar] [CrossRef]
- Opedal, Ø.H.; Armbruster, W.S.; Graae, B.J. Linking small-scale topography with microclimate, plant species diversity and intra-specific trait variation in an alpine landscape. Plant Ecol. Divers. 2015, 8, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Méndez-Toribio, M.; Meave, J.A.; Zermeño-Hernández, I.; Ibarra-Manríquez, G. Effects of slope aspect and topographic position on environmental variables, disturbance regime and tree community attributes in a seasonal tropical dry forest. J. Veg. Sci. 2016, 27, 1094–1103. [Google Scholar] [CrossRef]
- Muscarella, R.; Kolyaie, S.; Morton, D.C.; Zimmerman, J.K.; Uriarte, M. Effects of topography on tropical forest structure depend on climate context. J. Ecol. 2020, 108, 145–159. [Google Scholar] [CrossRef]
- Homeier, J.; Breckle, S.-W.; Günter, S.; Rollenbeck, R.T.; Leuschner, C. Tree Diversity, Forest Structure and Productivity along Altitudinal and Topographical Gradients in a Species-Rich Ecuadorian Montane Rain Forest: Ecuadorian Montane Forest Diversity and Structure. Biotropica 2010, 42, 140–148. [Google Scholar] [CrossRef]
- Pierick, K.; Leuschner, C.; Homeier, J. Topography as a factor driving small-scale variation in tree fine root traits and root functional diversity in a species-rich tropical montane forest. New Phytol. 2021, 230, 129–138. [Google Scholar] [CrossRef]
- Parmesan, C.; Hanley, M.E. Plants and climate change: Complexities and surprises. Ann. Bot. 2015, 116, 849–864. [Google Scholar] [CrossRef]
- Dobrowski, S.Z. A climatic basis for microrefugia: The influence of terrain on climate. Glob. Chang. Biol. 2011, 17, 1022–1035. [Google Scholar] [CrossRef]
- Bennett, K.D.; Tzedakis, P.C.; Willis, K.J. Quaternary Refugia of North European Trees. J. Biogeogr. 1991, 18, 103. [Google Scholar] [CrossRef]
- Vellend, M.; Béhé, M.; Carteron, A.; Crofts, A.L.; Danneyrolles, V.; Gamhewa, H.T.; Ni, M.; Rinas, C.L.; Watts, D.A. Plant Responses to Climate Change and an Elevational Gradient in Mont Mégantic National Park, Québec, Canada. Northeast. Nat. 2021, 28, 4–28. [Google Scholar] [CrossRef]
- Duque, A.; Stevenson, P.R.; Feeley, K.J. Thermophilization of adult and juvenile tree communities in the northern tropical Andes. Proc. Natl. Acad. Sci. USA 2015, 112, 10744–10749. [Google Scholar] [CrossRef] [Green Version]
- Feeley, K.J.; Silman, M.R.; Bush, M.B.; Farfan, W.; Cabrera, K.G.; Malhi, Y.; Meir, P.; Revilla, N.S.; Quisiyupanqui, M.N.R.; Saatchi, S. Upslope migration of Andean trees: Andean trees migrate upslope. J. Biogeogr. 2011, 38, 783–791. [Google Scholar] [CrossRef]
- Morin, X.; Chuine, I. Niche breadth, competitive strength and range size of tree species: A trade-off based framework to understand species distribution: Species range patterns and processes. Ecol. Lett. 2006, 9, 185–195. [Google Scholar] [CrossRef]
- Ibáñez, I.; Clark, J.S.; LaDeau, S.; Lambers, J.H.R. Exploiting temporal variability to understand tree recruitment response to climate change. Ecol. Monogr. 2007, 77, 163–177. [Google Scholar] [CrossRef]
- Anderson-Teixeira, K.J.; Miller, A.D.; Mohan, J.E.; Hudiburg, T.W.; Duval, B.D.; DeLucia, E.H. Altered dynamics of forest recovery under a changing climate. Glob. Chang. Biol. 2013, 19, 2001–2021. [Google Scholar] [CrossRef]
- De Frenne, P.; Rodriguez-Sanchez, F.; Coomes, D.A.; Baeten, L.; Verstraeten, G.; Vellend, M.; Bernhardt-Romermann, M.; Brown, C.D.; Brunet, J.; Cornelis, J.; et al. Microclimate moderates plant responses to macroclimate warming. Proc. Natl. Acad. Sci. USA 2013, 110, 18561–18565. [Google Scholar] [CrossRef] [Green Version]
- Scherrer, D.; Körner, C. Infra-red thermometry of alpine landscapes challenges climatic warming projections. Glob. Chang. Biol. 2010, 16, 2602–2613. [Google Scholar] [CrossRef]
- Willi, Y.; Van Buskirk, J. A review on trade-offs at the warm and cold ends of geographical distributions. Philos. Trans. R. Soc. B Biol. Sci. 2022, 377, 20210022. [Google Scholar] [CrossRef]
- Bin, Y.; Li, Y.; Russo, S.E.; Cao, H.; Ni, Y.; Ye, W.; Lian, J. Leaf trait expression varies with tree size and ecological strategy in a subtropical forest. Funct. Ecol. 2022, 36, 1010–1022. [Google Scholar] [CrossRef]
- Valladares, F.; Matesanz, S.; Guilhaumon, F.; Araújo, M.B.; Balaguer, L.; Benito-Garzón, M.; Cornwell, W.; Gianoli, E.; Kleunen, M.; Naya, D.E.; et al. The effects of phenotypic plasticity and local adaptation on forecasts of species range shifts under climate change. Ecol. Lett. 2014, 17, 1351–1364. [Google Scholar] [CrossRef] [Green Version]
- Loehle, C. Height growth rate tradeoffs determine northern and southern range limits for trees. J. Biogeogr. 1998, 25, 735–742. [Google Scholar] [CrossRef]
- Doak, D.F.; Morris, W.F. Demographic compensation and tipping points in climate-induced range shifts. Nature 2010, 467, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Taccoen, A.; Piedallu, C.; Seynave, I.; Gégout-Petit, A.; Gégout, J.-C. Climate change-induced background tree mortality is exacerbated towards the warm limits of the species ranges. Ann. For. Sci. 2022, 79, 23. [Google Scholar] [CrossRef]
- Kawecki, T.J. Adaptation to Marginal Habitats. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 321–342. [Google Scholar] [CrossRef] [Green Version]
- Bogdziewicz, M.; Szymkowiak, J.; Fernández-Martínez, M.; Peñuelas, J.; Espelta, J.M. The effects of local climate on the correlation between weather and seed production differ in two species with contrasting masting habit. Agric. For. Meteorol. 2019, 268, 109–115. [Google Scholar] [CrossRef]
- Kelly, D.; Geldenhuis, A.; James, A.; Penelope Holland, E.; Plank, M.J.; Brockie, R.E.; Cowan, P.E.; Harper, G.A.; Lee, W.G.; Maitland, M.J.; et al. Of mast and mean: Differential-temperature cue makes mast seeding insensitive to climate change. Ecol. Lett. 2013, 16, 90–98. [Google Scholar] [CrossRef]
- Koenig, W.D.; Knops, J.M.H. Patterns of Annual Seed Production by Northern Hemisphere Trees: A Global Perspective. Am. Nat. 2000, 155, 59–69. [Google Scholar] [CrossRef]
- Villellas, J.; Ehrlén, J.; Olesen, J.M.; Braza, R.; García, M.B. Plant performance in central and northern peripheral populations of the widespread Plantago coronopus. Ecography 2013, 36, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Kaarlejärvi, E.; Olofsson, J. Concurrent biotic interactions influence plant performance at their altitudinal distribution margins. Oikos 2014, 123, 943–952. [Google Scholar] [CrossRef]
- Michalet, R.; Brooker, R.W.; Cavieres, L.A.; Kikvidze, Z.; Lortie, C.J.; Pugnaire, F.I.; Valiente-Banuet, A.; Callaway, R.M. Do biotic interactions shape both sides of the humped-back model of species richness in plant communities? Ecol. Lett. 2006, 9, 767–773. [Google Scholar] [CrossRef]
- Toll, K. An evolutionary framework for understanding habitat partitioning in plants. Am. J. Bot. 2023, 110, e16119. [Google Scholar] [CrossRef] [PubMed]
- Afkhami, M.E.; McIntyre, P.J.; Strauss, S.Y. Mutualist-mediated effects on species’ range limits across large geographic scales. Ecol. Lett. 2014, 17, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Van der Putten, W.H. Climate Change, Aboveground-Belowground Interactions, and Species’ Range Shifts. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 365–383. [Google Scholar] [CrossRef]
- Louthan, A.M.; Doak, D.F.; Angert, A.L. Where and When do Species Interactions Set Range Limits? Trends Ecol. Evol. 2015, 30, 780–792. [Google Scholar] [CrossRef]
- HilleRisLambers, J.; Harsch, M.A.; Ettinger, A.K.; Ford, K.R.; Theobald, E.J. How will biotic interactions influence climate change-induced range shifts?: Biotic interactions and range shifts. Ann. N. Y. Acad. Sci. 2013, 1297, 112–125. [Google Scholar] [CrossRef]
- Lankau, R.A.; Keymer, D.P. Ectomycorrhizal fungal richness declines towards the host species’ range edge. Mol. Ecol. 2016, 25, 3224–3241. [Google Scholar] [CrossRef]
- Visser, M.E.; Gienapp, P. Evolutionary and demographic consequences of phenological mismatches. Nat. Ecol. Evol. 2019, 3, 879–885. [Google Scholar] [CrossRef]
- Kharouba, H.M.; Wolkovich, E.M. Disconnects between ecological theory and data in phenological mismatch research. Nat. Clim. Chang. 2020, 10, 406–415. [Google Scholar] [CrossRef]
- Miller-Struttmann, N.E.; Geib, J.C.; Franklin, J.D.; Kevan, P.G.; Holdo, R.M.; Ebert-May, D.; Lynn, A.M.; Kettenbach, J.A.; Hedrick, E.; Galen, C. Functional mismatch in a bumble bee pollination mutualism under climate change. Science 2015, 349, 1541–1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traveset, A.; Richardson, D.M. Mutualistic Interactions and Biological Invasions. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Van Nuland, M.E.; Peay, K.G. Symbiotic niche mapping reveals functional specialization by two ectomycorrhizal fungi that expands the host plant niche. Fungal Ecol. 2020, 46, 100960. [Google Scholar] [CrossRef]
- Van Grunsven, R.H.A.; Van Der Putten, W.H.; Bezemer, T.M.; Tamis, W.L.M.; Berendse, F.; Veenendaal, E.M. Reduced plant-soil feedback of plant species expanding their range as compared to natives. J. Ecol. 2007, 95, 1050–1057. [Google Scholar] [CrossRef]
- Van GRUNSVEN, R.H.A.; Van Der PUTTEN, W.H.; Martijn Bezemer, T.; Berendse, F.; Veenendaal, E.M. Plant–soil interactions in the expansion and native range of a poleward shifting plant species: Plant–soil feedback in a poleward shifting plant. Glob. Chang. Biol. 2010, 16, 380–385. [Google Scholar] [CrossRef]
- Novoplansky, A. Picking battles wisely: Plant behaviour under competition. Plant Cell Environ. 2009, 32, 726–741. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D. On the Meaning of Competition and the Mechanisms of Competitive Superiority. Funct. Ecol. 1987, 1, 304. [Google Scholar] [CrossRef]
- Case, T.J.; Holt, R.D.; McPeek, M.A.; Keitt, T.H. The community context of species’ borders: Ecological and evolutionary perspectives. Oikos 2005, 108, 28–46. [Google Scholar] [CrossRef]
- Hargreaves, A.L.; Samis, K.E.; Eckert, C.G. Are Species’ Range Limits Simply Niche Limits Writ Large? A Review of Transplant Experiments beyond the Range. Am. Nat. 2014, 183, 157–173. [Google Scholar] [CrossRef] [Green Version]
- Gaston, K.J. The Structure and Dynamics of Geographic Ranges; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Booth, M.G.; Hoeksema, J.D. Mycorrhizal networks counteract competitive effects of canopy trees on seedling survival. Ecology 2010, 91, 2294–2302. [Google Scholar] [CrossRef] [PubMed]
- Eckhart, V.M.; Geber, M.A.; Morris, W.F.; Fabio, E.S.; Tiffin, P.; Moeller, D.A. The Geography of Demography: Long-Term Demographic Studies and Species Distribution Models Reveal a Species Border Limited by Adaptation. Am. Nat. 2011, 178, S26–S43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkpatrick, B.L.; Bazzaz, F.A. Influence of Certain Fungi on Seed Germination and Seedling Survival of Four Colonizing Annuals. J. Appl. Ecol. 1979, 16, 515. [Google Scholar] [CrossRef]
- Louda, S.M. Distribution Ecology: Variation in Plant Recruitment over a Gradient in Relation to Insect Seed Predation. Ecol. Monogr. 1982, 52, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Janzen, D.H. Seed Predation by Animals. Annu. Rev. Ecol. Syst. 1971, 2, 465–492. [Google Scholar] [CrossRef]
- Russo, S.E. Linking seed fate to natural dispersal patterns: Factors affecting predation and scatter-hoarding of Virola calophylla seeds in Peru. J. Trop. Ecol. 2005, 21, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Russo, S.E.; Augspurger, C.K. Aggregated seed dispersal by spider monkeys limits recruitment to clumped patterns in Virola calophylla: Seed dispersal and plant recruitment. Ecol. Lett. 2004, 7, 1058–1067. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, M. Mean crowding. J. Anim. Ecol. 1967, 36, 1–30. [Google Scholar] [CrossRef]
- Pyke, G.H.; Pulliam, H.R.; Charnov, E.L. Optimal Foraging: A Selective Review of Theory and Tests. Q. Rev. Biol. 1977, 52, 137–154. [Google Scholar] [CrossRef] [Green Version]
- Connell, J.H. On the role of natural enemies in preventing competitive excusion in some marine animals and in rain forest trees. In Dynamics of Populations; Advanced Study Institute on Dynamics of Numbers in Populations: Wageningen, The Netherlands, 1971; pp. 298–312. [Google Scholar]
- Janzen, D.H. Herbivores and the Number of Tree Species in Tropical Forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Augspurger, C.K. Light Requirements of Neotropical Tree Seedlings: A Comparative Study of Growth and Survival. J. Ecol. 1984, 72, 777. [Google Scholar] [CrossRef]
- Condit, R.; Hubbell, S.P.; Foster, R.B. Density Dependence in Two Understory Tree Species in a Neotropical Forest. Ecology 1994, 75, 671–680. [Google Scholar] [CrossRef]
- Beckstead, J.; Meyer, S.E.; Molder, C.J.; Smith, C. A Race for Survival: Can Bromus tectorum Seeds Escape Pyrenophora semeniperda-caused Mortality by Germinating Quickly? Ann. Bot. 2007, 99, 907–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bever, J.D.; Platt, T.G.; Morton, E.R. Microbial Population and Community Dynamics on Plant Roots and Their Feedbacks on Plant Communities. Annu. Rev. Microbiol. 2012, 66, 265–283. [Google Scholar] [CrossRef] [Green Version]
- Chew, F.S.; Courtney, S.P. Plant Apparency and Evolutionary Escape from Insect Herbivory. Am. Nat. 1991, 138, 729–750. [Google Scholar] [CrossRef]
- Feeny, P. Plant Apparency and Chemical Defense. In Biochemical Interaction between Plants and Insects; Recent Advances in Phytochemistry; Springer: Boston, MA, USA, 1976; Volume 10. [Google Scholar]
- Mitchell, C.E.; Power, A.G. Release of invasive plants from fungal and viral pathogens. Nature 2003, 421, 625–627. [Google Scholar] [CrossRef]
- Colautti, R.I.; Ricciardi, A.; Grigorovich, I.A.; MacIsaac, H.J. Is invasion success explained by the enemy release hypothesis? Ecol. Lett. 2004, 7, 721–733. [Google Scholar] [CrossRef]
- Diez, J.M.; Dickie, I.; Edwards, G.; Hulme, P.E.; Sullivan, J.J.; Duncan, R.P. Negative soil feedbacks accumulate over time for non-native plant species: Plant-soil feedbacks change over time. Ecol. Lett. 2010, 13, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Russo, S.E.; Kitajima, K. The Ecophysiology of Leaf Lifespan in Tropical Forests: Adaptive and Plastic Responses to Environmental Heterogeneity. In Tropical Tree Physiology; Goldstein, G., Santiago, L.S., Eds.; Tree Physiology; Springer International Publishing: Cham, Switzerland, 2016; Volume 6, pp. 357–383. ISBN 978-3-319-27420-1. [Google Scholar]
- Matsuki, S.; Koike, T. Comparison of Leaf Life Span, Photosynthesis and Defensive Traits Across Seven Species of Deciduous Broad-leaf Tree Seedlings. Ann. Bot. 2006, 97, 813–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makino, T.T.; Sakai, S. Experience changes pollinator responses to floral display size: From size-based to reward-based foraging. Funct. Ecol. 2007, 21, 854–863. [Google Scholar] [CrossRef]
- Dauber, J.; Biesmeijer, J.C.; Gabriel, D.; Kunin, W.E.; Lamborn, E.; Meyer, B.; Nielsen, A.; Potts, S.G.; Roberts, S.P.M.; Sõber, V.; et al. Effects of patch size and density on flower visitation and seed set of wild plants: A pan-European approach: Patch size, flower visitation and seed set. J. Ecol. 2010, 98, 188–196. [Google Scholar] [CrossRef] [Green Version]
- Courchamp, F.; Clutton-Brock, T.; Grenfell, B. Inverse density dependence and the Allee effect. Trends Ecol. Evol. 1999, 14, 405–410. [Google Scholar] [CrossRef]
- Moeller, D.A.; Geber, M.A.; Eckhart, V.M.; Tiffin, P. Reduced pollinator service and elevated pollen limitation at the geographic range limit of an annual plant. Ecology 2012, 93, 1036–1048. [Google Scholar] [CrossRef] [PubMed]
- Groom, M.J. Allee Effects Limit Population Viability of an Annual Plant. Am. Nat. 1998, 151, 487–496. [Google Scholar] [CrossRef]
- Bennett, A.E.; Classen, A.T. Climate change influences mycorrhizal fungal–plant interactions, but conclusions are limited by geographical study bias. Ecology 2020, 101, e02978. [Google Scholar] [CrossRef]
- Wolfe, B.E.; Pringle, A. Geographically structured host specificity is caused by the range expansions and host shifts of a symbiotic fungus. ISME J. 2012, 6, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Mueller, T.L.; Karlsen-Ayala, E.; Moeller, D.A.; Bellemare, J. Of mutualism and migration: Will interactions with novel ericoid mycorrhizal communities help or hinder northward Rhododendron range shifts? Oecologia 2022, 198, 839–852. [Google Scholar] [CrossRef]
- Vogelsang, K.M.; Bever, J.D. Mycorrhizal densities decline in association with nonnative plants and contribute to plant invasion. Ecology 2009, 90, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Pigliucci, M. Phenotypic integration: Studying the ecology and evolution of complex phenotypes. Ecol. Lett. 2003, 6, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Schlichting, C.D. Phenotypic Integration and Environmental Change. BioScience 1989, 39, 460–464. [Google Scholar] [CrossRef]
- Ghalambor, C.K.; McKAY, J.K.; Carroll, S.P.; Reznick, D.N. Adaptive versus non-adaptive phenotypic plasticity and the potential for contemporary adaptation in new environments. Funct. Ecol. 2007, 21, 394–407. [Google Scholar] [CrossRef]
- Drenovsky, R.E.; Grewell, B.J.; D’Antonio, C.M.; Funk, J.L.; James, J.J.; Molinari, N.; Parker, I.M.; Richards, C.L. A functional trait perspective on plant invasion. Ann. Bot. 2012, 110, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Richards, C.L.; Bossdorf, O.; Muth, N.Z.; Gurevitch, J.; Pigliucci, M. Jack of all trades, master of some? On the role of phenotypic plasticity in plant invasions. Ecol. Lett. 2006, 9, 981–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Richards, C.L.; Valladares, F.; et al. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Matesanz, S.; Gianoli, E.; Valladares, F. Global change and the evolution of phenotypic plasticity in plants: Global change and plasticity. Ann. N. Y. Acad. Sci. 2010, 1206, 35–55. [Google Scholar] [CrossRef] [PubMed]
- Arnold, P.A.; Nicotra, A.B.; Kruuk, L.E.B. Sparse evidence for selection on phenotypic plasticity in response to temperature. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180185. [Google Scholar] [CrossRef] [Green Version]
- Levis, N.A.; Pfennig, D.W. Evaluating ‘Plasticity-First’ Evolution in Nature: Key Criteria and Empirical Approaches. Trends Ecol. Evol. 2016, 31, 563–574. [Google Scholar] [CrossRef]
- Schwander, T.; Leimar, O. Genes as leaders and followers in evolution. Trends Ecol. Evol. 2011, 26, 143–151. [Google Scholar] [CrossRef]
- West-Eberhard, M.J. Developmental Plasticity and Evolution; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Gupta, A.P.; Lewontin, R.C. A Study of Reaction Norms in Natural Populations of Drosophila pseudoobscura. Evolution 1982, 36, 934–948. [Google Scholar] [CrossRef]
- Price, T.D.; Qvarnström, A.; Irwin, D.E. The role of phenotypic plasticity in driving genetic evolution. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 1433–1440. [Google Scholar] [CrossRef] [Green Version]
- Fox, R.J.; Donelson, J.M.; Schunter, C.; Ravasi, T.; Gaitán-Espitia, J.D. Beyond buying time: The role of plasticity in phenotypic adaptation to rapid environmental change. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180174. [Google Scholar] [CrossRef] [PubMed]
- Velikova, V.; Arena, C.; Izzo, L.G.; Tsonev, T.; Koleva, D.; Tattini, M.; Roeva, O.; De Maio, A.; Loreto, F. Functional and Structural Leaf Plasticity Determine Photosynthetic Performances during Drought Stress and Recovery in Two Platanus orientalis Populations from Contrasting Habitats. Int. J. Mol. Sci. 2020, 21, 3912. [Google Scholar] [CrossRef] [PubMed]
- Chevin, L.-M.; Collins, S.; Lefèvre, F. Phenotypic plasticity and evolutionary demographic responses to climate change: Taking theory out to the field. Funct. Ecol. 2013, 27, 967–979. [Google Scholar] [CrossRef] [Green Version]
- Andersen, K.M.; Turner, B.L.; Dalling, J.W. Seedling performance trade-offs influencing habitat filtering along a soil nutrient gradient in a tropical forest. Ecology 2014, 95, 3399–3413. [Google Scholar] [CrossRef]
- Svenning, J.-C.; Gravel, D.; Holt, R.D.; Schurr, F.M.; Thuiller, W.; Münkemüller, T.; Schiffers, K.H.; Dullinger, S.; Edwards, T.C.; Hickler, T.; et al. The influence of interspecific interactions on species range expansion rates. Ecography 2014, 37, 1198–1209. [Google Scholar] [CrossRef] [PubMed]
- Benito Garzón, M.; Robson, T.M.; Hampe, A. ΔTraitSDMs: Species distribution models that account for local adaptation and phenotypic plasticity. New Phytol. 2019, 222, 1757–1765. [Google Scholar] [CrossRef]
- Pichancourt, J.-B.; van Klinken, R.D. Phenotypic Plasticity Influences the Size, Shape and Dynamics of the Geographic Distribution of an Invasive Plant. PLoS ONE 2012, 7, e32323. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, S.A. Effects of Habitat Fragmentation on the Reproductive Ecology of Four Plant Species in Mallee Woodland. Conserv. Biol. 2000, 14, 758–768. [Google Scholar] [CrossRef]
- Masson-Delmotte, V.; Intergovernmental Panel on Climate Change; WMO; United Nations Environment Programme. Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems: Summary for Policymakers; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2019; ISBN 978-92-9169-154-8. [Google Scholar]
- Albert, J.S.; Carnaval, A.C.; Flantua, S.G.A.; Lohmann, L.G.; Ribas, C.C.; Riff, D.; Carrillo, J.D.; Fan, Y.; Figueiredo, J.J.P.; Guayasamin, J.M.; et al. Human impacts outpace natural processes in the Amazon. Science 2023, 379, eabo5003. [Google Scholar] [CrossRef]
- Franklin, J. Mapping Species Distributions: Spatial Inference and Prediction; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Kearney, M.; Porter, W. Mechanistic niche modelling: Combining physiological and spatial data to predict species’ ranges. Ecol. Lett. 2009, 12, 334–350. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Charney, N.D.; Record, S.; Gerstner, B.E.; Merow, C.; Zarnetske, P.L.; Enquist, B.J. A Test of Species Distribution Model Transferability Across Environmental and Geographic Space for 108 Western North American Tree Species. Front. Ecol. Evol. 2021, 9, 689295. [Google Scholar] [CrossRef]
- Beaumont, L.J.; Hughes, L.; Poulsen, M. Predicting species distributions: Use of climatic parameters in BIOCLIM and its impact on predictions of species’ current and future distributions. Ecol. Model. 2005, 186, 251–270. [Google Scholar] [CrossRef]
- Kearney, M.; Porter, W.P. Mapping the fundamental niche: Physiology, climate, and the distribution of a nocturnal lizard. Ecology 2004, 85, 3119–3131. [Google Scholar] [CrossRef] [Green Version]
- Kearney, M.R.; Porter, W.P. NicheMapR—An R package for biophysical modelling: The ectotherm and Dynamic Energy Budget models. Ecography 2020, 43, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Moorcroft, P.R.; Hurtt, G.C.; Pacala, S.W. A method for scaling vegetation dynamics: The Ecosystem Demography Model (ED). Ecol. Monogr. 2001, 71, 557–586. [Google Scholar] [CrossRef]
- Medvigy, D.; Wofsy, S.C.; Munger, J.W.; Hollinger, D.Y.; Moorcroft, P.R. Mechanistic scaling of ecosystem function and dynamics in space and time: Ecosystem Demography model version 2. J. Geophys. Res. 2009, 114, G01002. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, S.; Lewandrowski, W.; Elliott, C.P.; Miller, B.P.; Turner, S.R. High-resolution distribution modeling of a threatened short-range endemic plant informed by edaphic factors. Ecol. Evol. 2020, 10, 763–777. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Yao, L.; Meng, J.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef]
- Zhang, L.; Zhu, L.; Li, Y.; Zhu, W.; Chen, Y. Maxent Modelling Predicts a Shift in Suitable Habitats of a Subtropical Evergreen Tree (Cyclobalanopsis glauca (Thunberg) Oersted) under Climate Change Scenarios in China. Forests 2022, 13, 126. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Graham, C.H. The ability of climate envelope models to predict the effect of climate change on species distributions. Glob. Chang. Biol. 2006, 12, 2272–2281. [Google Scholar] [CrossRef]
- Kearney, M.R.; Wintle, B.A.; Porter, W.P. Correlative and mechanistic models of species distribution provide congruent forecasts under climate change: Congruence of correlative and mechanistic distribution models. Conserv. Lett. 2010, 3, 203–213. [Google Scholar] [CrossRef]
- Araújo, M.B.; Peterson, A.T. Uses and misuses of bioclimatic envelope modeling. Ecology 2012, 93, 1527–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaac, N.J.B.; Jarzyna, M.A.; Keil, P.; Dambly, L.I.; Boersch-Supan, P.H.; Browning, E.; Freeman, S.N.; Golding, N.; Guillera-Arroita, G.; Henrys, P.A.; et al. Data Integration for Large-Scale Models of Species Distributions. Trends Ecol. Evol. 2020, 35, 56–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heberling, J.M.; Miller, J.T.; Noesgaard, D.; Weingart, S.B.; Schigel, D. Data integration enables global biodiversity synthesis. Proc. Natl. Acad. Sci. USA 2021, 118, e2018093118. [Google Scholar] [CrossRef]
- Lee-Yaw, J.A.; McCune, J.L.; Pironon, S.; Sheth, S.N. Species distribution models rarely predict the biology of real populations. Ecography 2022, 2022, e05877. [Google Scholar] [CrossRef]
- Rubenstein, M.A.; Weiskopf, S.R.; Carter, S.L.; Eaton, M.J.; Johnson, C.; Lynch, A.J.; Miller, B.W.; Morelli, T.L.; Rodriguez, M.A.; Terando, A.; et al. Do empirical observations support commonly-held climate change range shift hypotheses? A systematic review protocol. Environ. Evid. 2020, 9, 10. [Google Scholar] [CrossRef]
- Feng, X.; Park, D.S.; Walker, C.; Peterson, A.T.; Merow, C.; Papeş, M. A checklist for maximizing reproducibility of ecological niche models. Nat. Ecol. Evol. 2019, 3, 1382–1395. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.-Z.; Wang, C.-J.; Yu, F.-H. Effects of occurrence record number, environmental variable number, and spatial scales on MaxEnt distribution modelling for invasive plants. Biologia 2019, 74, 757–766. [Google Scholar] [CrossRef]
- Berec, L.; Angulo, E.; Courchamp, F. Multiple Allee effects and population management. Trends Ecol. Evol. 2007, 22, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Ehrlen, J.; Eriksson, O. Dispersal Limitation and Patch Occupancy in Forest Herbs. Ecology 2000, 81, 1667–1674. [Google Scholar] [CrossRef]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant Ecological Strategies: Some Leading Dimensions of Variation Between Species. Annu. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef] [Green Version]
- Stearns, S.C. The Evolution of Life Histories; Oxford University Press: London, UK, 1992. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McNichol, B.H.; Russo, S.E. Plant Species’ Capacity for Range Shifts at the Habitat and Geographic Scales: A Trade-Off-Based Framework. Plants 2023, 12, 1248. https://doi.org/10.3390/plants12061248
McNichol BH, Russo SE. Plant Species’ Capacity for Range Shifts at the Habitat and Geographic Scales: A Trade-Off-Based Framework. Plants. 2023; 12(6):1248. https://doi.org/10.3390/plants12061248
Chicago/Turabian StyleMcNichol, Bailey H., and Sabrina E. Russo. 2023. "Plant Species’ Capacity for Range Shifts at the Habitat and Geographic Scales: A Trade-Off-Based Framework" Plants 12, no. 6: 1248. https://doi.org/10.3390/plants12061248
APA StyleMcNichol, B. H., & Russo, S. E. (2023). Plant Species’ Capacity for Range Shifts at the Habitat and Geographic Scales: A Trade-Off-Based Framework. Plants, 12(6), 1248. https://doi.org/10.3390/plants12061248