Abstract
A new species of Marsupella sect. Ustulatae Müll. Frib. ex R.M. Schust. is described following an integrated morphological and molecular–phylogenetic study which examined the recently found dioicous plants growing epilithically on acidic substrates in several mountain ranges of Portugal between Peneda-Gerês in the north and Serra da Monchique in the extreme south. Employed molecular markers (plastid trnF–trnT region and nuclear ribosomal ITS) confirmed the distinctness of the lineage from other currently recognized species in the section, and furthermore, previously neglected diversity within M. sprucei (Limpr.) Bernet was signaled. Although not yet confirmed outside Portugal, the newly reported species is probably not rare in the region and has likely been overlooked as M. funckii (F. Weber & D. Mohr) Dumort. or M. profunda Lindb. in the past.
1. Introduction
The predominately Northern Hemisphere liverwort family Gymnomitriaceae currently includes nine genera [1], of which Marsupella Dumort. is the most speciose, with thirty-three species worldwide [1,2,3,4], seven of which are recently described and two representing infraspecific taxa elevated to species rank [2,3,4]. In Europe, there are currently sixteen accepted species [5], and in Portugal, six species are reported [6]. Many species have been synonymized over time, though it is likely some will be resurrected as further molecular data inform taxonomic recircumscriptions, as in the case of Marsupella aquatica (Lindenb.) Schiffn. [7] or, more recently, M. patens (N. Kitag.) Bakalin & Fedosov and M. vermiformis (R.M. Schust.) Bakalin & Fedosov [3].
During a decade of bryological exploration by R.D. Porley of Serra de Monchique, Algarve, Portugal, the most south-westerly located mountain range in continental Europe, numerous Marsupella collections were made. Most of these were determined as M. emarginata (Ehrh.) Dumort., but a few collections, mostly from the summit of Fóia, the highest peak at 902 m a.s.l., but also from the north flank, clearly belonged to a different plant characterized by closely appressed flat glossy brownish-black patches and leaves with plane leaf margins and mostly subacute to obtuse leaf lobes. These plants initially called to mind M. profunda Lindb., a European endemic known from further north in mainland Portugal, England and Macaronesia [8]. Subsequent microscopic study, however, indicated a subtle difference in the angle between the leaf lobes, mostly 60–90° in the Monchique plants, opposed to 40–60° in M. profunda [9], but more remarkable was that the Monchique plants appeared to be dioicous, whereas M. profunda is paroicous. Material was then sent to D.G. Long (RBG Edinburgh, Edinburgh, UK) and N.G. Hodgetts (Cuillin Views, Isle of Skye, UK), who both agreed that the Portuguese plants did not accord with M. profunda and that the plants were ostensibly dioicous. To resolve the identity of the Portuguese plants, R.D. Porley sent material to J. Kučera, who sequenced them to reveal the molecular affinities using the sequence data publicly accessible in GenBank and unpublished data from a DNA barcoding study on rare British bryophytes [10]. The comparison of sequence data for nuclear ribosomal ITS region and chloroplast trnL–trnF and trnH–psbA showed that the plants did not match M. profunda or any other previously sequenced species, except for two at that time unreleased molecular accessions (included in analyses here), retrieved from specimens collected by D. Bell and D.G. Long during a visit to northern Portugal (Bell 251 and Bell 247, respectively), and tentatively named Marsupella sprucei (Limpr.) Bernet [10]. Later examination of the specimens confirmed that the plants were apparently dioicous (M. sprucei is paroicous), with separate male and female plants observed in both collections. Three additional specimens of the unknown plant, confirmed later by molecular data, were found among Portuguese collections of Marsupella funckii (F. Weber & D. Mohr) Dumort. by J. Kučera, held in herbarium CBFS. Indeed, the prominent Czech hepaticologist Jiří Váňa (1940–2018), who revised the three specimens collected by J.K., confirmed his tentative identifications of the plants as M. funckii, having noted the dioicy in two of the specimens which bore gametangia.
With respect to the distinctive morphology of the plants and molecular differences from all other previously molecularly investigated species of Marsupella, we describe in this paper the species as new to science. The discovery on Serra de Monchique of a novel species, employing integrative taxonomic techniques [11], is not unprecedented; the mosses Coscinodon monchiquensis R.D. Porley, Ochyra & Ignatova and Neodicranella hamulosa R.D. Porley, Fedosov & Plášek were also recently described from the region [12,13], underlining the bryologically underexplored nature of the territory. By contrast, the more northern localities are considered bryologically much better worked, particularly in the case of Serra da Estrela [14] and Peneda-Gerês National Park [6,15,16,17]; Serra do Caramulo is, however, bryologically less well known [18].
2. Results
2.1. Molecular Data
Based on both ITS and trnF–trnT data, accessions of the Portuguese plants which are described below as a new species, Marsupella lusitanica, show little variation in ITS (two A/G substitutions in ITS1 in one of four analyzed accessions, two ITS1 substitutions further downstream in another accession, two one-base indels in two different poly-C, resp. poly-G regions of ITS2 in two of six accessions) and no variation in trnF–trnL region (trnL–trnT region was retrieved for only one accession). On the contrary, the sequences differ in eleven-point substitutions and three 2–8 bp indels from M. profunda only in ITS2, the only common region analyzed. Multiple substitutions and indels in both trnF–trnT and ITS regions differentiate the newly discovered species from other lineages of sect. Ustulatae Müll. Frib. ex R.M. Schust. as well.
Molecular synapomorphies of M. lusitanica are translated to the distinct pattern in tree topology summarizing the results of the phylogenetic analyses of both the concatenated data matrix (Figure 1) and the data from the analyses of individual markers, ITS and trnL–trnT (Supplementary Data, Figures S1 and S2, respectively). The well-supported lineage containing accessions of M. lusitanica is nested in a fully supported clade consisting of members of Marsupella sect. Ustulatae as understood by Bakalin et al. [3]: M. disticha Steph., M. funckii, M. bolanderi (Austin) Underw., M. sphacelata (Giesecke ex Lindenb.) Dumort., M. profunda and accessions of multiple lineages which are labeled as M. sprucei and M. neglecta (Limpr.) Lindb., a taxon generally treated as a variety or synonym of M. sprucei by recent authors [5,19,20,21]. This clade is unresolved in a polytomy with two other lineages, one corresponding to section Hyalacme (Lindb.) Bakalin & Fedosov (M. apiculata Schiffn., M. condensata (Ångstr. ex C. Hartm.) Lindb. ex Kaal.), and the other containing the clades corresponding to sections Stolonicaulon (N. Kitag.) Váňa, Boeckiorum Müll. Frib. ex R.M. Schust. and Marsupella. The latter section is the largest lineage containing the type of the genus, M. emarginata, and related species. A single accession of Poeltia campylata Grolle clusters with weak support with species of Marsupella sect. Hyalacme, while Marsupella including Poeltia appears very well supported, and Prasanthus Lindb., Gymnomitrion Corda and Nardia Gray branch off at successively deeper nodes. The affinities among lineages within sect. Ustulatae are poorly resolved except for the weakly supported sister relationship between M. disticha and the rest of the species, but accessions from most individual species are mostly well supported and show little variability. An interesting exception to this appears to be M. sprucei, accessions of which arise in multiple lineages, differently so in ITS and trnF–T-based datasets. ITS data (Figure S1) suggest differentiation into three lineages, the largest of them containing seven accessions from montane to alpine habitats of Central Europe and morphologically corresponding to M. sprucei in the strict sense, the second containing three accessions of plants labeled as M. sprucei from southwestern England and Wales, and the third containing accessions morphologically corresponding to M. neglecta from Central European alpine habitats (growing on soil in contrast to most occurrences of M. sprucei s.str. which were collected from epilithic habitats), and two molecularly somewhat divergent accessions from southern Siberia and the Russian Far East, for which morphological and ecological data are not known; the lineage including M. neglecta and two M. sprucei s.lat. accessions is nevertheless strongly supported. Chloroplast data (Figure S2) suggest a finer division into four lineages with unresolved mutual relationships (data for the UK specimens are unfortunately missing, but it is plausible to assume that they would form another distinct lineage); three of the lineages are formed by accessions of what appeared to form the nearly invariable ‘M. sprucei s.str.’ ITS lineage. Moreover, the two Russian accessions do not appear to be closely related based on their published trnL–trnF data, only the accession from Magadan region showing a moderately supported affinity with M. neglecta. The analysis of concatenated data (Figure 1) retains the ITS-based and morphology-supported clustering of M. sprucei s.str. (only weakly supported, PP 0.95/BS 69, given the chloroplast-based differences) and the clustering of M. neglecta with the Russian M. sprucei s.lat. accessions (with only a slightly weakened BS support) and resolves the British lineage of M. sprucei s.lat. distinct from both other M. sprucei s.lat. lineages.
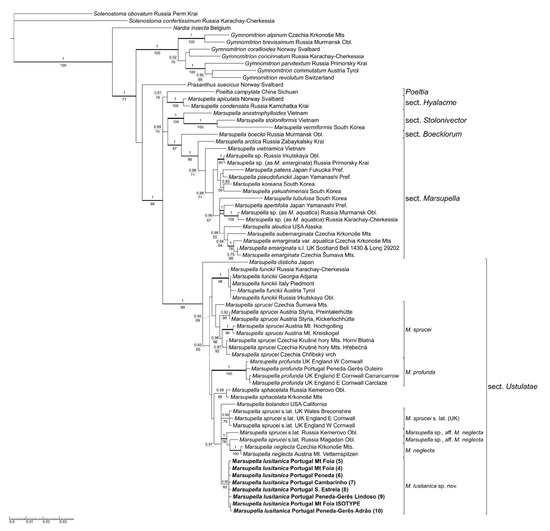
Figure 1.
Consensus tree from the Bayesian inference of concatenated ITS and trnF–trnT data, partitioned between ITS and trnF–trnT nucleotide data and standard data from indels coded by a simple coding method [19]. Accessions of Marsupella lusitanica are printed in bold. Support values representing posterior probabilities from Bayesian analysis are plotted above branches, while bootstrap support values from Maximum Likelihood analysis of the equally partitioned dataset appear below branches. Support values are shown only on branches receiving either PP > 0.9 or BS > 70. Branches which received maximum support from at least one of the analyses and accessions of Marsupella lusitanica appear in bold.
2.2. Description of the New Species
Marsupella lusitanica R.D. Porley & Jan Kučera, sp. nov. Figure 2, Figure 3 and Figure 4 and Figure S3.
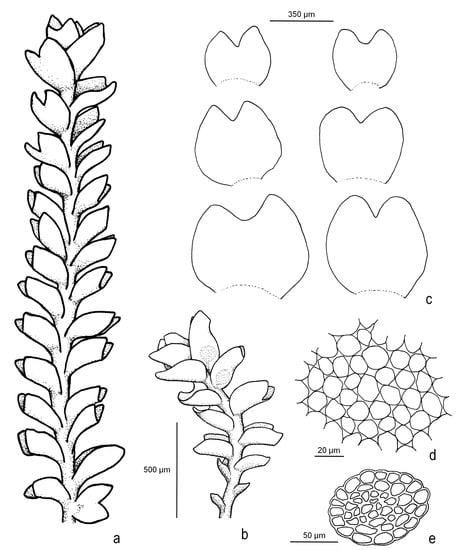
Figure 2.
Marsupella lusitanica R.D. Porley & Jan Kučera (all from Porley s.n., paratype 5): (a) sterile shoot, wet; (b) male plant, wet; (c) leaves; (d) cells in mid-lobe region; (e) cross-section of stem. Line drawing by Nick Hodgetts.

Figure 3.
Marsupella lusitanica R.D. Porley & Jan Kučera, habit microphotographs: (a) female plant with perianth (arrow) and sporophyte; (b) two male shoots; (c) section of gynoecium showing the perigynium, enlarged bracts, and a smaller perianth (arrow) inside; (d) two sterile shoots. (a,c) from Kučera 10623, paratype 7, (b) from Kučera 10685, paratype 8, (d) from holotype, Porley s.n.
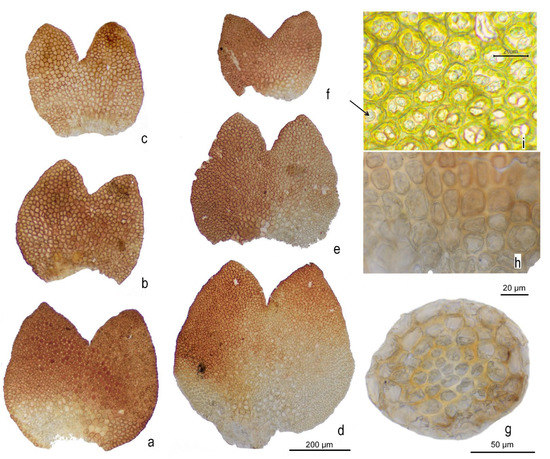
Figure 4.
Marsupella lusitanica R.D. Porley & Jan Kučera, detail microphotographs: (a–f) vegetative leaves; (g) stem cross-section; (h) basal cells; (i) mid-lobe cells showing the oil bodies; arrow to biconcentric oil body. (a–c,g) from holotype, Porley s.n., (d–e) from Kučera 10685, paratype 8, (f,h) from Kučera 10623, paratype 7, (i) from Porley s.n., paratype 3.
Etymology. The specific epithet refers to the geographical provenance (from Latin lusitanicus, Lusitanian, Portuguese) from which the new species is currently only known.
Diagnosis. The species differs from other species of Marsupella sect. Ustulatae Müll. Frib. ex R.M. Schust. in the combination of dioicous distribution of gametangia, slightly wider than long, semiovate leaves mostly with a relatively open sinus (angle between lobes at base of sinus mostly 40–80°, angle at lobe tips mostly 90–120°), descending to 22–30% of leaf length, and subacute to obtuse leaf lobes with relatively small cells, 12–18 µm wide at the base of lobes and 8–13 µm wide at lobe margins.
Type: Portugal: Algarve, Faro District, Serra de Monchique, Mt. Fóia, 37.317894° N 8.592727° W, altitude 876 m, on inclined face of syenite rock on north-facing rocky outcrop on summit, with Scapania compacta (Roth) Dumort., Grimmia trichophylla Grev. and Cephaloziella divaricata (Spruce) Schiffn., 13 September 2015, with sporophytes, leg. R.D. Porley, LISU. Isotype CBFS (20843).
Additional specimens examined (paratypes): (1) Algarve, Faro District, Fóia, Serra de Monchique, 37.317894° N 8.592727° W, altitude 876 m, syenite rock crevice on north-facing rocky outcrop on summit, 1 March 2019, with female gametangia, leg. R.D. Porley, herb. Porley; (2) ibidem, 13 March 2019, with female gametangia, leg. R.D. Porley, herb. Porley; (3) ibidem, in fissure of syenite block, N-facing, with crustose lichens, 19 January 2023, with archegonia, leg. R.D. Porley, herb. Porley; (4) ibidem, 37.319063° N 8.580492° W, 792 m, on syenite rock of terrace wall in crevice, N-facing aspect, with Hypnum cupressiforme, 3 December 2015, leg. R.D. Porley, herb. Porley, dupl. CBFS (20845); (5) ibidem, on syenite rock on terrace wall on sheltered north-facing slope, 21 November 2018, with antheridia, leg. R.D. Porley, herb. Porley, dupl. CBFS (20844); (6) Minho, Viana do Castelo District, Parque Nacional da Peneda-Gerês, Serra da Peneda: path Nossa Sra. da Peneda–Bouça dos Homens, at a brook 610 m WNW of the Na. Sra. da Peneda church, 41.977432° N 8.229002° W, ca. 920 m, shaded vertical face of a granitic boulder at the brook bank, N-facing, 4 July 2002, leg. J. Kučera 10,535 (CBFS); (7) Beira Alta, Viseu District, Serra do Caramulo, Cambarinho, on the edge of the laurel botanical reserve (ca. 0.6 km SE of the village), at a junction of tracks, 40.672565° N 8.201744° W, ca. 500 m, granitic stone by the track, light shade from the Pinus pinaster–Eucalyptus forest, 7 July 2002, leg. J. Kučera 10,623 (CBFS); (8) Beira Alta, Guarda District, Serra da Estrela, Loriga, path along the left riverbank of Ribeiro de Loriga, 130 m SW of the bridge 0.5 km SSW of the village center, 40.319168° N 7.697196° W, altitude 660 m, wall of granite stones, vertically, NW-or., not shaded, 9 July 2002, leg. J. Kučera 10,685 (CBFS); (9) Minho, Viana do Castelo District, Peneda-Gerês National Park, Lindoso Village, 41.867389° N 8.199222° W, castle wall, on granite rock, altitude 462 m, 11 June 2010, leg. D. Bell 251, with D.G. Long (E); (10) Minho, Viana do Castelo, Peneda-Gerês National Park, Branda de Bordenca, SW of Adrão, 41.909333° N 8.264611° W, river valley, on granite wall by path, altitude 640 m, 11 June 2010, leg. D. Bell 247, with D.G. Long (E).
Description.Plants small, in low mats or patches closely attached to rock, brown to brownish-ochre, young growth yellowish-green, black to purplish-black when dry, lobes often more heavily pigmented than base of leaf giving a scorched appearance, somewhat glossy when dry, shoots erect (3–)4–5(–7) mm long and 0.4–1.0 mm wide, with intercalary branches and stolons with reduced leaves. Rhizoids few, scattered, smooth to slightly roughened, arising from stem only, colorless on younger stems and stolons, brownish or vinaceous. Stem cross-section ±0.14–0.22 mm wide, hyalodermis apparently lacking but outermost cortical cells enlarged, with less thickened walls, chlorophyllose only in young shoots, hyaline and eroded in older shoots, thick-walled medullary cells distinctly smaller. Leaves distichously arranged, channeled, transversely inserted, semiovate, ca. 0.30–0.52 mm long and 0.35–0.58 mm wide, widest below middle, 1.0–1.25× wider than long, imbricate distally, diminishing in size and distant proximally, margins plane, antical margin not or barely decurrent, patent to spreading, keel straight to slightly arched, lobes ± equal in size, apices subacute, occasionally obtuse, sometimes one lobe obtuse the other subacute on same leaf, sinus descending along mostly convex lobe margins to ca. 22–31(–40)% of leaf length, angle between lobes ca. 40–80° at base of sinus, 90–120(–140)° at lobe tips. Leaf cells isodiametric to somewhat oblong, (10–)12–18(–20) μm wide and (12–)14–21(–24) µm long, trigones large and bulging, marginal cells of lobes 10–18 μm long 8–13 μm wide, cuticle smooth. Oil bodies 2–3 per cell, spherical 4.5–7.5 μm to ovoid 10(–12) μm long, nearly filling cell lumen, containing 4–8 smaller oil droplets bound by an outer, only slightly irregular membrane; smaller part of the oil bodies biconcentric. Dioicous. Androecia in the upper part of stems mostly followed by vegetative leaves towards apex (intercalary), 0.7–0.75 mm wide, with 3–5 paired bracts with ventricose base, bracts ±530 μm wide with recurved antical margin, antheridial body ovoid–spherical, to 125 μm in diameter, antheridial stalks biseriate, to 95 μm long. Gynoecia terminal, to 1.0 mm wide, with subgynoecial innovations, bracts strongly concave, up to 850 μm wide, margins weakly recurved, perigynia well developed, perianths ovoid–conical, leptodermous, 0.2–0.5 mm high, over-topped by elongated upper bracts to 0.9 mm long, cells at mouth of perianth 35–40 μm long. Capsule spherical, spores almost smooth, (10–)11–12(–13) μm, elaters bispiral, 5–8 μm wide.
Differentiation.Marsupella lusitanica is characterized by a combination of relatively small size, dioicous sexuality, semi-ovate leaves with subacute to occasionally obtuse leaf lobes and moderately open sinus descending less than one-third of leaf length (Figure 4a–f), the glossy dark brown to purplish black color in the dry state (Figure 5; note, however the brownish appearance in the wet state, Figure S3), and the epilithic occurrence on acidic substrates in oceanic-montane but not high montane or alpine environments of westernmost Iberian Peninsula. With respect to the dioicy, which is a relatively rare condition in species of the sect. Ustulatae, shared by the wide-Holarctic-distributed M. funckii and M. sphacelata and the western North American M. bolanderi, M. lusitanica shares the general aspect of the smaller forms of the exceedingly variable (and possibly taxonomically heterogeneous) M. funckii. However, the typical form of M. funckii differs in leaves having acute widely triangular leaf lobes with wide (60–90°) and more deeply (1/3–1/2 of leaf length) descending sinus. The taxa described as M. badensis Schiffn. and M. pygmaea (Limpr.) Steph. are more similar to M. lusitanica in leaf shape, particularly the less deep sinus and somewhat ovate leaf lobes, but M. badensis is described as more robust (stems 1–2 cm tall as opposed to ca. 0.5(–0.7) cm tall stems of M. lusitanica), with the leaf sinus still more open and lobes with a mostly straight rather than convex line [22,23,24], and M. pygmaea being even smaller than M. lusitanica, with densely appressed leaves with acute lobes but larger leaf cells, 18–21(–25) µm [22,23,25]. The taxon described as M. ramosa Müll. Frib. shares the leaf shape of M. badensis but has substantially larger cells, 20–25 × 25–30 µm [22,23]. Obtuse lobes, the distribution in Portugal and apparently very similar ecology suggest the most likely confusion between M. lusitanica and M. profunda, but that species can safely be ruled out by its paroicous sexuality if gametangia are at hand, which fortunately is mostly the case in M. profunda. Gametophytic characters alone provide a less reliable differentiation; M. profunda usually has a different leaf shape, with leaves longer than wide (in M. lusitanica the leaves are typically 1.1–1.2× wide as long, Figure 4a–f), and broader, often lingulate lobes [26]. The sinus in M. profunda is often deeper and more distinctly U-shaped, and leaf cell dimension is marginally greater, although there is overlap. Perianth mouth cell length may be useful; for M. lusitanica, they were found to range between 25 and 40 μm, while measurements made in M. profunda indicate they are somewhat longer, ranging between 30 and 70 μm. However, only a small number (3–4) of perianths of M. lusitanica were available for study, whereas sporophytes with perianths are frequent in the paroicous M. profunda. Another dioicous species of the section, M. sphacelata, is usually substantially more robust, 1–5(–8) cm tall, with suborbicular to 2 mm long and wide leaves with obtuse lobes and very narrow sinus, and a distinct stem hyalodermis. The western North American M. bolanderi has a somewhat similar appearance, being a small reddish-brown dioicous species with suborbicular to semiovate leaves somewhat wider than long and a relatively deep (30–40%), wide sinus. Apart from the generally more open sinus, the median cells in M. bolanderi are larger (mostly 20–25 µm wide), and stem cortex cells thin-walled [27]. The small size, semi-ovate leaves and moderately wide sinus mostly between 20% and 30% of leaf length is seen in M. sprucei s.lat., which probably accommodates several taxa, as indicated by our molecular data. All taxa recognized within M. sprucei are nevertheless monoicous (mostly paroicous, although synoicous gametangia have been reported by Limpricht [25] and disputed in a follow-up account by Schiffner [28]). In M. sprucei s.str., fertile plants are distinctly clavate, with densely imbricate, julaceous leaves when dry, and the median leaf cells described as substantially larger than in M. lusitanica, about 20–25 µm, which is true also for our accessions from Czechia and a part of the Austrian collections (Köckinger 15,429, 15,430), which form two of the trnF–T-based lineages; plants from the other two Austrian collections which constitute another trnF–T-based lineage have variable cells, with most of them nevertheless rather significantly smaller, between 12 and 16 µm. In the taxon described as M. neglecta, which appears to be a distinct species based on our data, both the habit of plants (not particularly densely foliated, pectinate on sterile shoots, never clavate on fertile shoots) and leaf form and cell sizes are similar to M. lusitanica, but the lobes of vegetative leaves are sharply acute and the plants are clearly paroicous and nearly always fertile [25]. The differences among species of the sect. Ustulatae which are most likely to be confused with M. lusitanica at its localities are summarized in Table 1.

Figure 5.
Dry habit of Marsupella lusitanica (paratype 1), showing the characteristic purplish black color, with Scapania compacta, Racomitrium heterostichum and Grimmia trichophylla. Photo by R.D. Porley, 1 March 2019.

Table 1.
Comparison of selected character traits among Marsupella sprucei, M. funckii, M. profunda and M. lusitanica.
Marsupella lusitanica may on occasion also be confused with the dioicous M. emarginata growing in the same places; indeed, M. emarginata is frequently on the same rock outcrop (type locality) and terrace wall as M. lusitanica. However, normally it grows in more sheltered niches such as in hollows and under overhanging rocks, where it is usually more robust, but it also extends onto more exposed surfaces and onto protosoils surrounding outcropping rock. A study of these forms confirmed that when in the dry state, they never develop the intense shiny black coloration or the scorched appearance typical of M. lusitanica (Figure 5), and careful microscopic observation usually reveals at least traces of recurved antical leaf margins, which is generally the most useful discriminating character; note, however, that perigonial, perigynial and subperigynial bracts in M. lusitanica have distinctly recurved margins as well (Figure 3a,b). The leaf shape and sinus also differ. In M. emarginata, the leaves are orbicular or transversely elliptical (rarely very slightly longitudinally elliptical) in outline, with the widest place at about the middle of the leaf, with a markedly narrowed, at times cordate base, while the leaves of M. lusitanica are generally semi-ovate in outline, with the widest place below the middle (Figure 4a–f). The sinus is generally wider in M. emarginata, variously deep and the lobe shape is generally quite variable, but small forms of the species that are likely to be confused with M. lusitanica have the sinus mostly rectangular, and about one-third of the leaf length, with acute lobes.
3. Discussion
3.1. Molecular Affinities
Based on the molecular synapomorphies of the unidentified Marsupella plants from Serra de Monchique, Peneda-Gerês, Serra da Estrela and Serra do Caramulo, the plants should either be recognized as a distinct species within the sect. Ustulatae, or all species of the sect. Ustulatae, as understood by Bakalin et al. [3], should be synonymized as one species. Given the morphological differences with other currently recognized species of the section Ustulatae and the dioicy which otherwise only occurs in morphologically and molecularly different M. bolanderi, M. funckii and M. sphacelata, the description of the Portuguese plants as a new species is clearly the preferable option. At the same time, the discovery of the molecular heterogeneity of plants referrable to M. sprucei as understood in recent floras and checklists calls for a detailed study of these non-monophyletic units, the current understanding of which arose as a result of numerous synonymizations in earlier treatments as compared to the work of K. Müller [23]. The latter (the last comprehensive European liverwort flora) recognized at the species level M. ustulata Spruce and M. sprucei, differing from the former in substantially larger leaf cells; M. neglecta and M. gracilis (C. Massal. & Carestia) Pearson were recognized as identical at the varietal level under M. ustulata. The varietal status of M. ustulata and M. neglecta has been retained by Damsholt [29,30], and our molecular data, coupled with morphological and ecological features which match the original description [25], confirm that at least M. neglecta should clearly be recognized as a distinct species. The different signals from nuclear and plastid markers, along with sometimes overlapping morphological traits, do not allow for a clear separation of M. sprucei and M. ustulata; however, the number of studied accessions is so far too low for safely founded conclusions. A similar situation can be expected in taxa synonymized under M. funckii, where morphological variation may in the future be shown to be supported by molecular traits. The past synonymizations often occurred without much discussion. Thus, the synonymy of M. ustulata with M. sprucei was justified with the existence of intermediate cell sizes [20], the merger of M. ramosa with M. funckii was justified with the suggestion of the former being ‘a large-celled modification (probably polyploid)’ of the latter [31], and the synonymy of M. pygmaea and M. badensis received no justification at all [32,33]. On the other hand, the molecular support for the recognition of M. aquatica, as advocated by Vilnet et al. [7] and later authors, might be based on the neglect of cryptic diversity within the morphs corresponding to those taxa. European plants corresponding to M. emarginata and M. aquatica (both those selected by us and those deposited in GenBank by previous authors other than Vilnet et al. [7] and Bakalin et al. [3]) show either no differences at all or a different signal from ITS and plastid data (cf. Figure 1, Figures S1 and S2), and at the same time, they are molecularly clearly different from the plants (of mostly Russian provenance) sequenced under that name, despite us employing the most characteristic morphs corresponding to both taxa. This suggests the existence of two additional, morphologically probably nearly cryptic species in the M. emarginata complex, in addition to those revealed by Bakalin et al. [3], and simultaneously, the arguable specific distinctness of M. aquatica.
3.2. Habitat and Conservation
Serra de Monchique, situated at 37° latitude on the extreme southwest point of mainland Europe, Portugal, in Algarve province, lies within the Mediterranean macrobioclimate region of the Iberian Peninsula, defined by warm, wet winters with markedly dry summers. The highest point, Fóia, at 902 m, receives over 1000 mm of rainfall per year [34]; however, the E–W orientation of the massif and the close proximity to the North Atlantic Ocean results in warm, moist air currents bringing summer fog to N- and NW-facing aspects. The central part of the massif is known as the Monchique alkaline complex [35] and comprises rocks of alkaline feldspar and nepheline syenite, a silica under-saturated rock rich in alumina, alkalis, rare earths (e.g., Nd) and trace elements (e.g., Rb, Ba, Nb, Th and Pb), low in magnesium and iron [36,37], differentiating it from granite. Parque Nacional Peneda-Gerês is located in the north of Portugal in the provinces of Minho and Trás-os-Montes e Alto Douro. It is a mountainous area, dominated by granite rocks, near to the transition between the Mediterranean and Atlantic macrobioclimatic regions with a Euoceanic Continentality, and the diverse topography results in various microclimates that support vegetation characteristic of Mediterranean, Eurosiberian and Alpine phytogeographic environments. Of the summits, Peneda is at 1340 m and Gerês is at 1545 m, and receives precipitation levels in excess of 3000 mm per year at higher altitudes [15,16]. Serra da Estrela is located in central Portugal, in Beira Alta province, and is the highest point in mainland Portugal, at 1993 m, comprising a massive granite ridge. It experiences mainly a Mediterranean and Atlantic bioclimate, but also with Continental, Alpine and Boreal elements with a mean annual precipitation of about 2500 mm in its higher parts [14]. Serra do Caramulo, in Beira Alta, comprising granitic rocks, also experiences a Mediterranean and Atlantic bioclimate, with a mean annual precipitation of just over 2300 mm, but due to its lower altitude (the highest elevation Pico do Caramulinho at 1074 m), lacks alpine and continental elements [18].
The habitat of the newly described species has been explored in detail only in Serra de Monchique. Here, Marsupella lusitanica occurs on a natural dome-shaped outcrop of syenite blocks and on an old terrace stone wall constructed from syenite rock on the north-facing flank of the mountain. It grows closely appressed to rock, typical of a pioneer species and characteristic of the genus [21]. At the type locality, only female plants have been detected. It occurs at several points, albeit in a small quantity, around the more humid base and lower levels of a north-facing ‘tor’ or outcrop at 876 m altitude (Figure 6), on exposed crustose and foliose lichen-covered rock surfaces and in shallow fissures (Figure 6 and Figure S4) typically mixed with Grimmia trichophylla Grev., Hypnum cupressiforme Hedw., Scapania compacta (Roth) Dumort., Racomitrium heterostichum (Hedw.) Brid., Cephaloziella divaricata (Sm.) Schiffn., Frullania tamarisci (L.) Dumort., Cynodontium bruntonii (Sm.) Bruch & Schimp. and, more rarely, Frullania fragilifolia (Taylor) Gottsche, Lindenb. & Nees. Marsupella emarginata is also present. The hyperoceanic microclimate of this outcrop is evidenced by the presence of the Cisto crispi–Ulicetum minoris gorse formation in which the lower woody stems of Cistus salvifolius L. support a rich epiphytic bryophyte community including Antitrichia curtipendula (Hedw.) Brid., Ulota calvescens Wilson and several Orthotrichum species [38]. However, significantly, also on the same outcrop is Racomitrium lanuginosum (Hedw.) Brid., a species in the circumpolar boreo-arctic montane element [39]. The other known locality (sites of paratypes 4, 5) on Serra de Monchique for M. lusitanica is ca. 1 km to the east on the north side of Fóia, at 784–792 m a.s.l., where it grows in at least two places on old neglected terraced north-facing moss-covered walls of syenite (Figure S5), partially overgrown by vegetation (Crataegus monogyna Jacq., Pteridium aquilinum (L.) Kuhn, Rhododendron ponticum subsp. baeticum (Boiss. & Reut.) Hand.-Mazz., Rubus ulmifolius Schott). This is the only site where male plants were detected; the majority of plants examined are female. Marsupella emarginata also occurs on the same wall. Much less detail is known about other sites in Portugal where the species was not collected intentionally, either supposed to be M. sprucei (DB’s collections from Peneda-Gerês) or M. funckii (J.K.’s collections from Peneda-Gerês, Serra da Estrela and Serra do Caramulo). In all three sites from the more northern Portuguese localities, the species was noted on vertical faces of granitic stones (either natural or from stone walls) in sheltered, moderately humid situations.

Figure 6.
North-facing syenite rock outcrops on the summit of Fóia, Serra de Monchique, type locality of Marsupella lusitanica, one of the micropopulations (paratype 3) arrowed. Photo by R.D. Porley, 19 January 2023.
Of the 16 European species of Marsupella, most show a northern distribution, in either the boreo-arctic montane or arctic-montane floristic element [39]. Marsupella emarginata shows a wider distribution, in the European Boreo-temperate element, and only one shows a much narrower range, M. profunda, classified as oceanic southern-temperate. In Portugal, M. profunda is reported from Parque Nacional Peneda-Gerês, Parque Natural do Alvão, Parque Natural da Serra de São Mamede and Parque Natural da Serra da Estrela; the latter site also supports M. sprucei, a taxon classified as Endangered [6] in Portugal. All these localities are in the high mountains of mid to northern Portugal. The currently known distribution of M. lusitanica (Figure 7) suggests a similar distribution pattern to M. profunda in Portugal, although it has not yet been recorded in Alvão and São Mamede natural parks, and, conversely, M. profunda has not been recorded in the Monchique mountains. These differences, however, rather suggest insufficient sampling effort in most Portuguese regions outside Peneda-Gerês and Serra da Estrela; it is most likely that further field sampling and revision of herbarium material will reveal additional populations. Given the broader distribution range of M. profunda that nevertheless has been revealed long after the initial discovery of the species, the possibility of endemicity of M. lusitanica to either Portugal or the Iberian Peninsula would be speculative and premature.
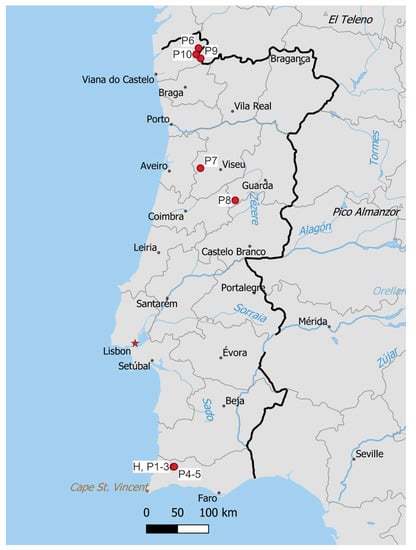
Figure 7.
Known distribution of Marsupella lusitanica. H denotes the locality of holotype, P1–10 correspond to localities of paratypes 1–10. Made with Natural Earth.
The population of Marsupella lusitanica in southern Portugal occurs within the 76,000 ha Monchique Natura 2000 Special Area of Conservation (SAC), yet it is debatable just how effective this designation is in protecting the biodiversity and natural communities and indeed the bryophytes, an issue highlighted in a recent horizon-scanning study to address fundamental questions in bryology [40]. The SAC has been extensively planted with Eucalyptus and pine and more ominously, wildfires are an ever-increasing threat compounded by rapid climate change and global warming [41]. A particularly devasting wildfire in 2018 covered over 27,000 ha on the northern and eastern flanks of the mountain, yet the impact on M. lusitanica and other bryophytes remains largely unknown. Rock outcrops are listed on Annex 1 of the European Community Habitats Directive (Siliceous rocky slopes with chasmophytic vegetation) yet are known to be particularly vulnerable to fire [42]. Parque Nacional Peneda-Gerês and Parque Natural da Serra da Estrela are both designated as SAC, and both areas have been impacted by extreme wildfire events, a situation that has been suggested as becoming the ‘new normal’ in Portugal [43]. Marsupella profunda, a species with an oceanic European and Macaronesian distribution, was assessed as ‘Vulnerable’ under criteria D1 in the last IUCN Red List of Threatened Species in Europe [44], meaning that while its population is not considered significantly declining, its size is small, estimated to be less than 1000 individual-equivalents. For M. lusitanica, the known extent of occurrence and area of occupation, as well as the documented population size, is apparently less, yet its actual distribution is obscured by greater uncertainty. The same inconclusiveness applies to possible trends in population dynamics. Even so, M. lusitanica arguably qualifies at a minimum for the ‘Vulnerable’ category worldwide (currently under both D1 and D2 criteria), prone to both effects of human activities and stochastic events within a short time period in an uncertain future. However, as the population is also possibly declining, it might potentially be assessed under B or C criteria, under which scenario it may be categorized as ‘Endangered’, even if the area of occupancy was between 10 and 500 km2 and the population size was estimated at between 250 and 2500 individual-equivalents, which may conceivably be a realistic approximation.
4. Materials and Methods
4.1. Molecular Investigation
Publicly accessible sequence data for most European Marsupella species and other genera of Gymnomitriaceae exist for the chloroplast region trnL–trnF and the nuclear ribosomal ITS. As for M. profunda, sequence data from ITS2, trnH–psbA, rbcL and matK were available from the unpublished DNA barcoding study on rare British bryophytes [10]. As there are limited publicly accessible rbcL or trnH-psbA data for Marsupella, no publicly accessible data for matK, and both rbcL and trnH-psbA are relatively uninformative for the genus, we focused our study on plastid trnF–trnL and nuclear ribosomal ITS to place the sequences of the new species into phylogenetic context. The dataset for the phylogenetic analysis was based on the available sequences in the GenBank database, which were retrieved using BLAST searches, and on the results of previously published molecular-phylogenetic studies in Marsupella by Bakalin et al. [3,4]. Sequencing of the trnF–trnL and ITS loci was carried out in the laboratories of the Faculty of Science, University of South Bohemia in České Budějovice, following the protocols for DNA isolation, amplification and sequencing described by Kučera et al. [45]. Where material allowed for the amplification of longer regions, we preferred to retrieve the chloroplast trnF–trnL including the adjacent variable trnL–trnT and trnT–trnL spacers. In these cases, amplification and sequencing employed the TabF [46] or our newly designed trnF-1F primer (5′– TGC CAG AAA CCA GAT TTG AAC TG –3′) as forward primers, and either the trnT-Bryo-R (5′– GGA GTC GAA CCG ATG ACC AT –3′) or rps4-22R (5′– GAG GTC CTC GAT AAC GNG ACA TAA –3′) as reverse primers (also designed by us). Newly generated sequences and others used for the context of the study are listed in the Appendix A.
Raw sequence reads were checked for reading errors and edited (primer complements and low-quality ends trimmed) and aligned with sequences downloaded from GenBank using the online interface of Mafft ver. 7 (https://mafft.cbrc.jp/alignment/server/, accessed on 18 March 2023 [47]) using the E-INS-i strategy with otherwise default options; the resulting alignment was checked and manually edited where necessary in Geneious Prime software (www.geneious.com, accessed on 18 March 2023). ITS and trnF–trnT matrices were evaluated independently, and as no topological conflict was discovered at supported nodes, a concatenated matrix was built and evaluated in phylogenetic context. The dataset was partitioned into ITS and trnF–trnT DNA data partitions and a standard data partition with indels scored using the simple-coding method [19]. Phylogenetic affinities were assessed using the Bayesian inference in MrBayes v. 3.2.7a [48] and Maximum Likelihood analysis using RAxML v. 8.2.12 [49], run at the cluster facilities of Metacentrum Virtual Organization (see Acknowledgements). The parameters for the analysis were set and trees from the analysis were summarized as described in Kučera et al. [45]. The trees were visualized using the TreeGraph2 software [50] and further edited graphically in Inkscape 1.2 (https://inkscape.org/, www.geneious.com) under the GPL license.
4.2. Morphological Investigation
The new species and specimens of species used for comparison were examined by standard morphological methods employing light microscopy and consultation of relevant morphological treatments and floras [9,22,23,29,51]. Emphasis was placed on characters most likely to provide distinction from other species in section Ustulatae and Marsupella emarginata from the same biogeographic region. The material collected and investigated by R.D. Porley is preserved in his personal herbarium (herb. Porley), the material investigated by J. Kučera is filed in herbarium CBFS and the material collected by D. Bell is kept in herbarium E. Abbreviations of herbaria follow the Index Herbariorum database (https://sweetgum.nybg.org/science/ih/, accessed on 18 March 2023).
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12071468/s1, Figure S1: Consensus tree from the Bayesian inference of ITS data; Figure S2: Consensus tree from the Bayesian inference of plastid trnF–trnT data; Figure S3: Wet habit of Marsupella lusitanica; Figure S4: Detail of a site with a patch of M. lusitanica; Figure S5: Locality of paratypes 4 and 5.
Author Contributions
Conceptualization, R.D.P. and J.K.; sampling and fieldwork, R.D.P., D.B. and J.K; retrieval and analysis of molecular data, J.K. and D.B.; retrieval and analysis of morphological data, R.D.P. and J.K.; writing—original draft preparation, R.D.P.; writing—review and editing, J.K., R.D.P. and D.B.; figures, R.D.P. and J.K.; funding, J.K. All authors have read and agreed to the published version of the manuscript.
Funding
Molecular analyses performed by Jan Kučera at University of South Bohemia were funded by the institutional sources of the Faculty of Science. Computational resources (‘Metacentrum VO’) were supplied by the Ministry of Education, Youth and Sports of the Czech Republic under the Projects CESNET (project no. LM2015042).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
DNA sequences are available on the GenBank database and all authors agree with MDPI Research Data Policies.
Acknowledgments
Many thanks to Nick Hodgetts, who kindly produced the line drawings and who accompanied one of us (R.D.P.) to see the plant in the field. We also acknowledge the expert opinion of David Long, to whom we sent specimens. Finally, a personal thanks to Cecilia Sérgio for her constant encouragement and support over the years of R.D.P.’s pursuit of bryophytes on Serra de Monchique. J.K. acknowledges the effective laboratory work by Alžběta Manukjanová (University of South Bohemia) that enabled the generation of molecular data for this study.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Table A1.
Voucher information and GenBank accession numbers (ordered trnL–trnF(–trnT), ITS) of the specimens used in the molecular study. Newly obtained sequences are printed in bold. Accessions used in the study of Bell are identified using BOLD Process IDs which appear in the final BOLD and GenBank records upon their release.
Table A1.
Voucher information and GenBank accession numbers (ordered trnL–trnF(–trnT), ITS) of the specimens used in the molecular study. Newly obtained sequences are printed in bold. Accessions used in the study of Bell are identified using BOLD Process IDs which appear in the final BOLD and GenBank records upon their release.
| Species | Voucher Information | GenBank Accession Number | |
| ITS | trnF–trnL (trnT) | ||
| Gymnomitrion alpinum | Czech Republic: Krkonoše Mts., Úpská jáma, Kučera 21057 (CBFS) | OQ474588 | OQ507748 |
| Gymnomitrion brevissimum | Russia: Murmansk Obl., N. Konstantinova G 8171 (KPABG) | EU791833 | EU791711 |
| Gymnomitrion commutatum | Austria: Tyrol, Mt Kreuzjoch, Kučera 18862 (CBFS) | OQ474596 | OQ507758 |
| Gymnomitrion concinnatum | Russia: Caucasus, Karachay-Cherkessia, N. Konstantinova K 465a-05 (KPABG) | EU791831 | EU791710 |
| Gymnomitrion corallioides | Norway: Svalbard, N. Konstantinova 155-04 (KPABG) | EU791826 | EU791705 |
| Gymnomitrion parvitextum | Russia: Primorsky Krai, Mamontov 170-1-10 (KPABG) | MF521472 | MF521482 |
| Gymnomitrion revolutum | Switzerland, Kiebacher 2235 (LE) | OQ024235 | OQ029671 |
| Marsupella aleutica | USA: Alaska, Schofield 103958 (MO) | MH826408 | MH822632 |
| Marsupella anastrophylloides | Vietnam: Hà Giang Prov., Bakalin & Klimova, V-15-6-20 (VBGI) | OM480746 | OM489480 |
| Marsupella apertifolia | Japan: Yamanashi Pref., Bakalin J-87-25-15 (VBGI) | MH539835 | MH539892 |
| Marsupella apiculata | Norway: Svalbard, N. Konstantinova K 93-1-06 (KPABG) | EU791819 | EU791699 |
| Marsupella arctica | Russia: Zabaykalsky Krai, Kodar Range, Mamontov 2-July-2013 (CBFS) | OQ474591 | OQ507752 |
| Marsupella boeckii | Russia: Murmansk Obl., Revda, Kučera 11525 (CBFS) | OQ474590 | OQ507751 |
| Marsupella bolanderi | USA: California, Santa Barbara, Santa Yen Mts., 38802 (KPABG) | MF521463 | MF521475 |
| Marsupella condensata | Russia: Kamchatka Krai, Bakalin K-60-30-15 (VGBI) | MH539844 | MH539901 |
| Marsupella disticha | Japan: Deguchi & Yamaguchi, Bryophytes of Asia 170 (KPABG) | EU791824 | EU791703 |
| Marsupella emarginata | Czech Rep.: Šumava Mts., Mt. Plechý, Kučera 23914 (CBFS) | OQ474584 | OQ507743 |
| Marsupella emarginata | UK: Scotland, Bell 1430 | OL434465 | – |
| Marsupella emarginata var. aquatica | UK: Scotland, Long 29202 (E) | – | KF942959 |
| Marsupella emarginata var. aquatica | Czech Rep.: Krkonoše Mts., Úpská jáma, Kučera 20264 (CBFS) | OQ474583 | OQ507742 |
| Marsupella funckii | Russia: Caucasus, Karachay-Cherkessia, N. Konstantinova K 516-1-05 (KPABG) | EU791820 | EU791700 |
| Marsupella funckii | Georgia: Adjaria, Bakalin G-17-2-13 (VGBI) | MH539854 | MH539911 |
| Marsupella funckii | Italy: Piedmont, Ghiffa, Kučera 15227 (CBFS) | OQ474581 | OQ507740 |
| Marsupella funckii | Austria: Tyrol, Zillergrund, Kučera 18904 (CBFS) | OQ474587 | OQ507747 |
| Marsupella funckii | Russia: Irkutskaya Obl., Pik Taltsinskyi, Kučera 20642 (CBFS) | OQ474593 | OQ507754 |
| Marsupella koreana | South Korea: Gyeongsang Prov., Bakalin kor-23-18-15 (VGBI) | MH539850 | MH539907 |
| Marsupella lusitanica, isotype | Portugal: Algarve, Mt. Fóia, Porley s.n. 13-Sep-2018 (CBFS:20843) | OQ474573 | OQ507732 |
| Marsupella lusitanica | Portugal: Algarve, Mt. Fóia, Porley s.n. 21-Nov-2018 (CBFS:20844) | OQ474574 | OQ507733 |
| Marsupella lusitanica | Portugal: Algarve, Mt. Fóia, Porley s.n. 3-Dec-2015 (CBFS:20844) | – | OQ507746 |
| Marsupella lusitanica | Portugal: Minho, Peneda-Gerês NP, Peneda, Kučera 10535 (CBFS) | OQ474592 | OQ507753 |
| Marsupella lusitanica | Portugal: Beira Alta, Campia, Cambarinho, Kučera 10623 (CBFS) | – | OQ507756 |
| Marsupella lusitanica | Portugal: Beira Alta, Loriga, Kučera 10685 (CBFS) | OQ474595 | OQ507757 |
| Marsupella lusitanica | Portugal: Minho, Peneda-Gerês NP, Lindoso, Bell 257 (E) | LWT1248-22 | – |
| Marsupella lusitanica | Portugal: Minho, Peneda-Gerês NP, Adrão, Bell 241 (E) | LWT1269-22 | – |
| Marsupella neglecta | Czech Rep.: Krkonoše Mts., Mt. Sněžka, Kučera 18476 (CBFS) | OQ474575 | OQ507734 |
| Marsupella neglecta | Austria: Styria, Vetternspitzen, Kučera 14668 (CBFS) | OQ474594 | OQ507755 |
| Marsupella patens | Japan: Fukuoka Pref., Bakalin J-7-26a-14 (VBGI) | MH539846 | MH539903 |
| Marsupella profunda | Portugal: Minho, Peneda-Gerês, Outeiro, Bell 154 (E) | LWT1242-22 | – |
| Marsupella profunda | UK: England, vc2, Carrancarrow, Callaghan 1069 (E) | LWT1249-22 | – |
| Marsupella profunda | UK: England, vc2, Carclaze, Holyoak 09-47 | LWT1245-22 | – |
| Marsupella profunda | UK: England, vc1, Lower Bostraze, Holyoak 09-40 | LWT1246-22 | – |
| Marsupella pseudofunckii | Japan: Yamanashi Pref., Bakalin J-88-23-15 (VBGI) | MH539852 | MH539909 |
| Marsupella sp. (as M. aquatica) | Russia: Murmansk Obl., Konstantinova 152/5-87 (KPABG) | EU791813 | AF519201 |
| Marsupella sp. (as M. aquatica) | Russia: Caucasus, Karachay-Cherkessia, N. Konstantinova K 517-4-05 (KPABG) | EU791814 | EU791694 |
| Marsupella sp. (as M. emarginata) | Russia: Primorsky Krai, Bakalin & Klimova, Prim-16-14-16 (VBGI) | MH539848 | MH539905 |
| Marsupella sp. | Russia: Irkutskaya Obl., Snezhnaya River valley, Kučera 20641 (CBFS) | OQ474582 | OQ507741 |
| Marsupella sphacelata | Russia: Kemerovo Obl., N. Konstantinova 65/1-00 (KPABG) | EU791821 | AF519200 |
| Marsupella sphacelata | Czech Rep.: Krkonoše Mts., Úpská jáma, Kučera 21077 (CBFS) | OQ474597 | OQ507759 |
| Marsupella sprucei | Czech Rep.: Šumava Mts., Mt. Plechý, F. Müller s.n. 3-October-2007 (DR, dupl. CBFS:20918) | OQ474576 | OQ507735 |
| Marsupella sprucei | Czech Rep.: Krušné hory Mts., Horní Blatná, F. Müller s.n. 18-September-2018 (DR, dupl. CBFS:20919) | OQ474577 | OQ507736 |
| Marsupella sprucei | Czech Rep.: Krušné hory Mts., Hřebečná, S. Biedermann s.n. 18-September-2018 (DR, dupl. CBFS:20920) | OQ474578 | OQ507737 |
| Marsupella sprucei | Czech Rep.: Děčín distr., Chřibský vrch, F. Müller s.n. 17-July-2019 (DR, dupl. CBFS:20965) | OQ474579 | OQ507738 |
| Marsupella sprucei | Austria: Styria, Preintalerhütte, Köckinger 15430 (CBFS) | OQ474585 | OQ507744 |
| Marsupella sprucei | Austria: Styria, Kickerlochhütte, Köckinger 15429 (CBFS) | OQ474586 | OQ507745 |
| Marsupella sprucei | Austria: Salzburg, Mt. Hochgolling, Kučera 9363 (CBFS) | OQ474589 | OQ507749 |
| Marsupella sprucei | Austria: Styria, Mt. Kreiskogel, Kučera 6391 (CBFS) | – | OQ507750 |
| Marsupella sprucei s.lat. | Russia: Kemerovo Obl., N. Konstantinova 54-1-00 (KPABG) | EU791823 | HQ833031 |
| Marsupella sprucei s.lat. | Russia: Magadan Obl., Bakalin mag-38-39-11 (VGBI) | MH539856 | MH539913 |
| Marsupella sprucei s.lat. | UK: England, vc1, Bakers’ Pit, Bell 531 (E) | LWT1261-22 | – |
| Marsupella sprucei s.lat. | UK: England, vc2, St. Neot, Holyoak 99-465 | LWT1259-22 | – |
| Marsupella sprucei s.lat. | UK: Wales, vc42, Lower Neuadd Res., Bell 566 (E) | LWT1262-22 | – |
| Marsupella stoloniformis | Vietnam: Lao Cai Prov., Bakalin & Klimova, V-11-11-17 (VBGI) | MH539859 | MH539916 |
| Marsupella subemarginata | Czech Rep.: Krkonoše Mts., Mt. Kotel, Kučera 23513 (CBFS) | OQ474580 | OQ507739 |
| Marsupella vermiformis | South Korea: Jeju Prov., Choi 120911 (VBGI) | MH539857 | MH539914 |
| Marsupella vietnamica | Vietnam: Lao Cai Prov., Bakalin, V-2-101-16 (VBGI) | MH539862 | MH539919 |
| Marsupella tubulosa | South Korea, South Gyeongsang Prov., Bakalin Kor-23-15-15 (VBGI) | MH539861 | MH539918 |
| Marsupella yakushimensis | South Korea, Gangwon Prov., Choi 8347 (VBGI) | MH539864 | MH539921 |
| Nardia insecta | Belgium, N. Konstantinova 102077 (KPABG) | EU791836 | EU791714 |
| Poeltia campylata | China: Sichuan, Bakalin 48-2-17, 37210 (VGBI) | MH580596 | MH580593 |
| Prasanthus suecicus | Norway: Svalbard, N. Konstantinova K 121-5-06 (KPABG) | EU791825 | EU791704 |
| Solenostoma confertissimum | Russia: Caucasus, Karachay-Cherkessia, N. Konstantinova K 459-8a-05 (KPABG) | GQ220774 | GQ220758 |
| Solenostoma obovatum | Russia: Perm Krai, N. Konstantinova K 324-1-04 (KPABG) | GQ220771 | GQ220755 |
References
- Söderström, L.; Hagborg, A.; von Konrat, M.; Bartholomew-Began, S.; Bell, D.; Briscoe, L.; Brown, E.; Cargill, D.C.; da Costa, D.P.; Crandall-Stotler, B.J.; et al. World checklist of hornworts and liverworts. PhytoKeys 2016, 59, 1–828. [Google Scholar] [CrossRef] [PubMed]
- Mamontov, Y.S.; Vilnet, A.A.; Konstantinova, N.A.; Bakalin, V.A. Two new species of Gymnomitriaceae (Marchantiophyta) in the North Pacific. Bot. Pac. 2019, 8, 67–80. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Fedosov, V.E.; Fedorova, A.V.; Nguyen, V.S. Integrative Taxonomic Revision of Marsupella (Gymnomitriaceae, Hepaticae) Reveals Neglected Diversity in Pacific Asia. Cryptogam. Bryol. 2019, 40, 59–85. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Vilnet, A.A.; Mamontov, Y.S.; Schäfer-Verwimp, A.; Maltseva, Y.D.; Klimova, K.G.; Nguyen, V.S.; Choi, S.S. Stolonicaulon: A Section-Puzzle within Marsupella (Gymnomitriaceae, Marchantiophyta). Plants 2022, 11, 1596. [Google Scholar] [CrossRef] [PubMed]
- Hodgetts, N.G.; Söderström, L.; Blockeel, T.L.; Caspari, S.; Ignatov, M.S.; Konstantinova, N.A.; Lockhart, N.; Papp, B.; Schröck, C.; Sim-Sim, M.; et al. An annotated checklist of bryophytes of Europe, Macaronesia and Cyprus. J. Bryol. 2020, 42, 1–116. [Google Scholar] [CrossRef]
- Sérgio, C.; Garcia, C.; Sim-Sim, M.; Vieira, C.; Hespanhol, H.; Stow, S. Atlas e Livro Vermelho dos Briófitos Ameaçados de Portugal; MUHNAC, Documenta: Lisboa, Spain, 2013. [Google Scholar]
- Vilnet, A.A.; Konstantinova, N.A.; Troitsky, A.V. Molecular insight on phylogeny and systematics of the Lophoziaceae, Scapaniaceae, Gymnomitriaceae and Jungermanniaceae. Arctoa 2010, 19, 31–50. [Google Scholar] [CrossRef]
- Porley, R.; Papp, B.; Söderström, L.; Hallingbäck, T. European Bryophyte Conservation in the New Millennium; University of Malaya: Kuala Lumpur, Malaysia, 2008. [Google Scholar]
- Paton, J.A. The Liverwort Flora of the British Isles; Harley Books: Colchester, UK, 1999; ISBN 978-0-946589-60-9. [Google Scholar]
- Bell, D.; Long, D.; Hollingsworth, P. The Use of DNA Barcoding to Address Major Taxonomic Problems for Rare British Bryophytes; Final Revised Report; Royal Botanic Garden Edinburgh: Edinburgh, UK, 2013. [Google Scholar]
- Dayrat, B. Towards integrative taxonomy. Biol. J. Linn. Soc. 2005, 85, 407–415. [Google Scholar] [CrossRef]
- Ignatov, M.S.; Porley, R.D.; Ochyra, R.; Kuznetsova, O.I.; Ignatova, E.A. Coscinodon monchiquensis R.D. Porley, Ochyra & Ignatova (Grimmiaceae), a new species from the Algarve, southern Portugal. J. Bryol. 2018, 40, 125–136. [Google Scholar] [CrossRef]
- Porley, R.D.; Fedosov, V.; Plášek, V.; Fedorova, A. Undiscovered Biodiversity of the European Moss Flora: Neodicranella hamulosa (Aongstroemiaceae), a New Genus and Species from SW Portugal. Plants 2021, 10, 2289. [Google Scholar] [CrossRef]
- Garcia, C.; Sérgio, C.; Jansen, J. The bryophyte flora of the natural park of Serra da Estrela (Portugal): Conservation and Biogeographical approaches. Cryptogam. Bryol. 2008, 29, 49–73. [Google Scholar]
- Sérgio, C.; Garcia, C.; Hespanhol, H.; Vieira, C.; Stow, S.; Long, D. Diversidad de briófitos en el Parque Nacional Peneda-Gerês (Portugal): Seleccionando áreas importantes de plantas “Important Plant Areas (IPA)” basado en nuevos datos y anteriores referencias. Bot. Complut. 2012, 36, 39–50. [Google Scholar] [CrossRef]
- Sérgio, C.; Schumacker, R. Contribuiçao para o estudo da flora briológica do Parque Nacional da Peneda-Gerês. Port. Acta Biol. 1992, 16, 107–137. [Google Scholar]
- Hespanhol, H.; Vieira, C.; Lomba, Â.; Séneca, A. New data on bryophyte distribution in the Peneda-Gerês National Park (NW Portugal): The use of GIS for conservation remarks. Bol. Soc. Esp. Briol. 2005, 26–27, 59–74. [Google Scholar]
- Sérgio, C.; Silva, I. The bryophyte flora of the Vouga region (Aveiro, Beira Litoral). Port. Acta Biol. 2007, 22, 159–188. [Google Scholar]
- Simmons, M.P.; Ochoterena, H. Gaps as Characters in Sequence-Based Phylogenetic Analyses. Syst. Biol. 2000, 49, 369–381. [Google Scholar] [CrossRef]
- Grolle, R. Miscellanea Hepaticologica 151–160. Lindbergia 1975, 3, 47–56. [Google Scholar]
- Váňa, J.; Söderström, L.; Hagborg, A.; Von Konrat, M.; Engel, J.J. Early Land Plants Today: Taxonomy, systematics and nomenclature of Gymnomitriaceae. Phytotaxa 2010, 11, 1. [Google Scholar] [CrossRef]
- Müller, K. Die Lebermoose Deutschlands, Oesterreichs und der Schweiz, 2nd ed.; Dr. L. Rabenhorst’s Kryptogamen-Flora von Deutschland, Oesterreich und der Schweiz; E. Kummer: Leipzig, Germany, 1906–1911; Volume 6. [Google Scholar]
- Müller, K. Die Lebermoose Europas (Musci hepatici), 3rd ed.; Dr. L. Rabenhorst’s Kryptogamen-Flora von Deutschland, Oesterreich und der Schweiz; Geest & Portig: Leipzig, Germany, 1956; Volume 6. [Google Scholar]
- Schiffner, V.F. Kritische Bemerkungen über die europäische Lebermoose mit Bezug auf die Exemplare des Exsiccatenwerkes: Hepaticae europaeae exsiccatae. I. Serie. Sitzungsber. Deutsch. Naturwiss. Med. Ver. Böhmen “Lotos” Prag 1901, 49, 75–130. [Google Scholar]
- Limpricht, K.G. Über neue Arten und Formen der Gattung Sarcoscyphus Corda. Jahresber. Schles. Ges. Vaterländische Cult. 1880, 58, 179–184. [Google Scholar]
- Paton, J.A. Marsupella profunda Lindb in Cornwall, new to the British Isles. J. Bryol. 1990, 16, 1–4. [Google Scholar] [CrossRef]
- Konstantinova, N.A. Marsupella. In FNA Editors, Gymnomitriaceae, Bryophyte Flora of North America; Provisional Publication, 2020. Available online: http://www.mobot.org/plantscience/bfna/V3/Gymnomitriaceae.htm (accessed on 18 March 2023).
- Schiffner, V. Studien über kritische Arten der Gattungen Gymnomitrium und Marsupella. (Fortsetzung.). Oesterr. Bot. Z. 1903, 53, 166–172. [Google Scholar] [CrossRef]
- Damsholt, K. Illustrated Flora of Nordic Liverworts and Hornworts; Nordic Bryological Society: Lund, Sweden, 2002; ISBN 87-986781-2-4. [Google Scholar]
- Damsholt, K. The Liverworts of Greenland; Nordic Bryological Society: Lund, Sweden, 2013; ISBN 978-91-637-2514-2. [Google Scholar]
- Váňa, J. Notes on the genus Marsupella s. lat. (Gymnomitriaceae, Hepaticae) 1–10. Infrageneric taxa. Bryobrothera 1999, 5, 221–229. [Google Scholar]
- Grolle, R. Verzeichnis der Lebermoose Europas und benachbarter Gebiete. Feddes Repert. 1976, 76, 171–279. [Google Scholar] [CrossRef]
- Duda, J.; Váňa, J. Rozšíření játrovek v Československu—XXXVII [Die Verbreitung der Lebermoose in der Tschechoslowakei—XXXVII]. Časopis Slez. Muz., Sér. A, Vědy Přír. 1983, 32, 97–110. [Google Scholar]
- Malato Beliz, J. A Serra de Monchique: FLORA e Vegetação; Parques Naturais; Serviço Nacional de Parques Reservas e Património Paisagístico: Lisboa, Portugal, 1982. [Google Scholar]
- González-Clavijo, E.J.; Valadares, V. A Estrutura do Complexo de Monchique. In Proceedings of the Actas do VI Congresso Nacional de Geologia Ciências da Terra (UNL), Lisboa, Portugal, 4–6 June 2003; Volume No. Especial 5, pp. 37–40. [Google Scholar]
- Rock, N.M.S. Petrology and Petrogenesis of the Monchique Alkaline Complex, Southern Portugal. J. Petrol. 1978, 19, 171–214. [Google Scholar] [CrossRef]
- Bernard-Griffiths, J.; Gruau, G.; Cornen, G.; Azambre, B.; Mace, J. Continental Lithospheric Contribution to Alkaline Magmatism: Isotopic (Nd, Sr, Pb) and Geochemical (REE) Evidence from Serra de Monchique and Mount Ormonde Complexes. J. Petrol. 1997, 38, 115–132. [Google Scholar] [CrossRef]
- Lara, F.; Porley, R.D.; Draper, I.; Aleffi, M.; Garcia, C.; Garilleti, R. Ulota s.l. (Orthotrichaceae, Bryidae) at southernmost Mediterranean localities: Not a simple matter. Pl. Biosyst. 2022, 156, 1448–1455. [Google Scholar] [CrossRef]
- Hill, M.O.; Preston, C.D. The geographical relationships of British and Irish bryophytes. J. Bryol. 1998, 20, 127–226. [Google Scholar] [CrossRef]
- Patiño, J.; Bisang, I.; Goffinet, B.; Hedenäs, L.; McDaniel, S.; Pressel, S.; Stech, M.; Ah-Peng, C.; Bergamini, A.; Caners, R.T.; et al. Unveiling the nature of a miniature world: A horizon scan of fundamental questions in bryology. J. Bryol. 2022, 44, 1–34. [Google Scholar] [CrossRef]
- Pörtner, D.C.; Roberts, M.; Tignor, E.S.; Poloczanska, K.; Mintenbeck, A.; Alegría, M.; Craig, S.; Langsdorf, S.; Löschke, V.; Möller, A.; et al. IPCC Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022. [Google Scholar]
- Mitchell, R.J.; Simonson, W.; Flegg, L.A.; Santos, P.; Hall, J. A comparison of the resilience of four habitats to fire, and the implications of changes in community composition for conservation: A case study from the Serra de Monchique, Portugal. Plant Ecol. Divers. 2009, 2, 45–56. [Google Scholar] [CrossRef]
- Tedim, F.; Leone, V.; Amraoui, M.; Bouillon, C.; Coughlan, M.; Delogu, G.; Fernandes, P.; Ferreira, C.; McCaffrey, S.; McGee, T.; et al. Defining Extreme Wildfire Events: Difficulties, Challenges, and Impacts. Fire 2018, 1, 9. [Google Scholar] [CrossRef]
- Sérgio, C.; Bergamini, A.; Garcia, C.; Garilleti, R.; Infante, M.; Porley, R.D. Marsupella profunda. The IUCN Red List of Threatened Species 2019: E.T39195A87726839 2019. Available online: https://www.gbif.org/fr/species/176752393 (accessed on 18 March 2023). [CrossRef]
- Kučera, J.; Kuznetsova, O.I.; Manukjanová, A.; Ignatov, M.S. A phylogenetic revision of the genus Hypnum: Towards completion. Taxon 2019, 68, 628–660. [Google Scholar] [CrossRef]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant. Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef]
- Schuster, R.M. The Hepaticae and Anthocerotae of North America; Columbia University Press: New York, NY, USA; London, UK, 1974; Volume III. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).