

Preventive and Curative Effects of Salicylic and Methyl Salicylic Acid Having Antifungal Potential against Monilinia laxa and the Development of Phenolic Response in Apple Peel




Abstract
:1. Introduction
2. Results
2.1. Effects of Preventive and Curative Use of SA and MeSA on Brown Rot
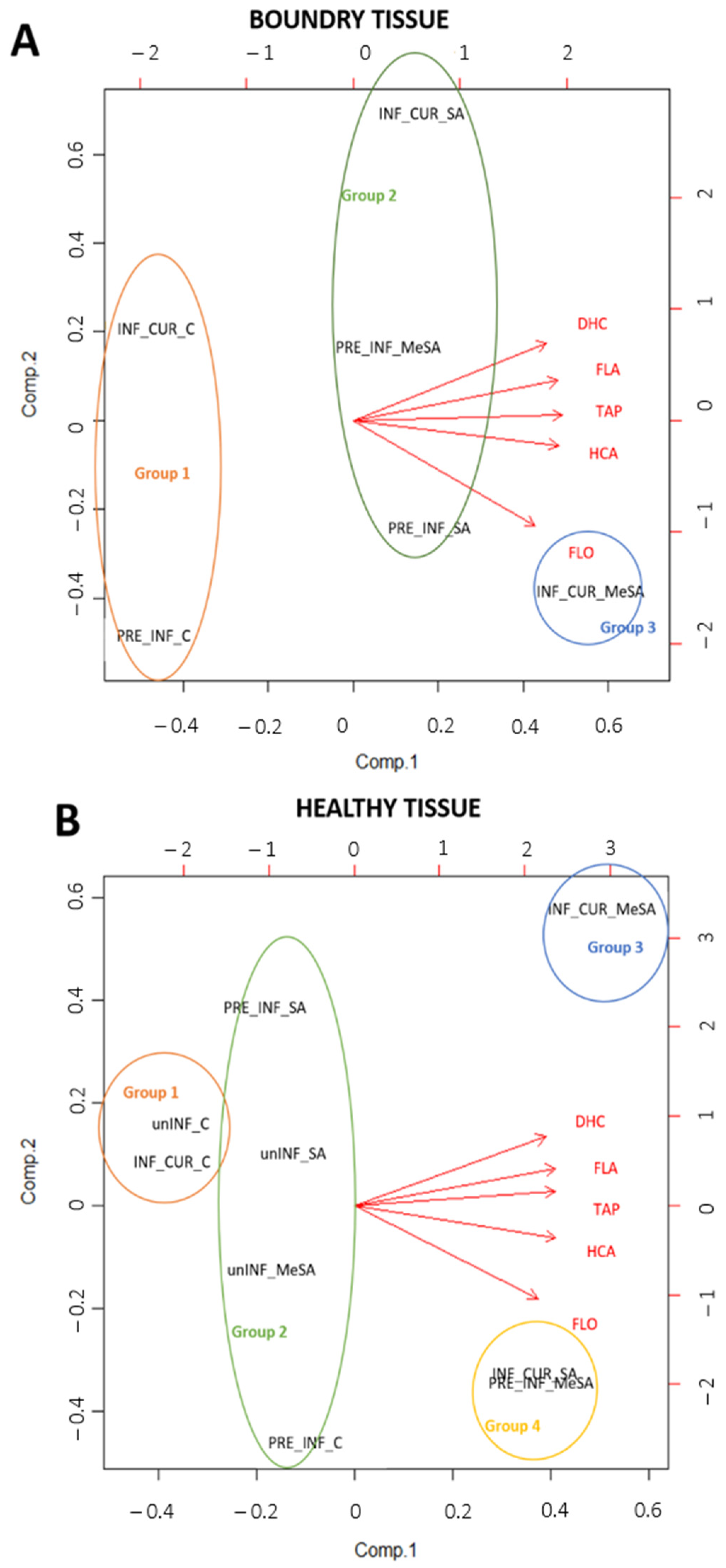
2.2. Effects of Preventive and Curative Use of SA and MeSA and Infection with M. laxa on Phenolic Composition
3. Discussion
4. Materials and Methods
4.1. Plant Material, Pathogen and Preparation of SA and MeSA Solutions
4.2. Treatments, Infection and Sampling
4.3. Individual Phenolic Compounds HPLC–MS Analysis of Apple Peel
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Babalar, M.; Asghari, M.; Talaei, A.; Khosroshahi, A. Effect of pre- and postharvest salicylic acid treatment on ethylene production, fungal decay and overall quality of Selva strawberry fruit. Food Chem. 2007, 105, 449–453. [Google Scholar] [CrossRef]
- Calzadilla, A.; Rehdanz, K.; Betts, R.; Falloon, P.; Wiltshire, A.; Tol, R.S.J. Climate change impacts on global agriculture. Clim. Chang. 2013, 120, 357–374. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M.; Macarisin, D.; Wilson, C. Twenty years of postharvest biocontrol research: Is it time for a new paradigm? Postharvest Biol. Technol. 2009, 52, 137–145. [Google Scholar] [CrossRef]
- Bremmer, J.; Gonzalez-Martinez, A.; Jongeneel, R.; Huiting, H.; Stokkers, R. Impact Assessment Study on EC 2030 Green Deal Targets for Sustainable Food Production: Effects of Farm to Fork and Biodiversity Strategy 2030 at Farm, National and EU Level; Report 2021-150; Wageningen Economic Research: Wageningen, The Netherlands, 2021. [Google Scholar]
- Fayet, C.M.J.; Reilly, K.H.; Van Ham, C.; Verburg, P.H. What is the future of abandoned agricultural lands? A systematic review of alternative trajectories in Europe. Land Use Policy 2022, 112, 105833. [Google Scholar] [CrossRef]
- Moschitz, H.; Muller, A.; Kretzschmar, U.; Haller, L.; Porras, M.; Pfeifer, C.; Oehen, B.; Willer, H.; Stolz, H. How can the EU Farm to Fork strategy deliver on its organic promises? Some critical reflections. EuroChoices 2021, 20, 30–36. [Google Scholar] [CrossRef]
- Tyagi, P.; Singh, A.; Gupta, A.; Prasad, M.; Ranjan, R. Chapter 4—Mechanism and function of salicylate in plant toward biotic stress tolerance. In Emerging Plant Growth Regulators in Agriculture; Aftab, T., Naeem, M., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 131–164. [Google Scholar] [CrossRef]
- Hayat, S.; Ahmad, A.; Alyemeni, M.N. Salicylic Acid: Plant Growth and Development; Springer: Dordrecht, The Netherlands, 2013; 681p. [Google Scholar]
- Hayat, S.; Ahmad, A. Salicylic Acid: A Plant Hormone; Springer: Dordrecht, The Netherlands, 2007; 401p. [Google Scholar]
- Klessig, D.F.; Malamy, J. The salicylic acid signal in plants. Plant Mol. Biol. 1994, 26, 1439–1458. [Google Scholar] [CrossRef]
- Raskin, I. Role of salicylic acid in plants. Annu. Rev. Plant Biol. 1992, 43, 439–463. [Google Scholar] [CrossRef]
- Stella de Freitas, T.F.; Stout, M.J.; Sant’Ana, J. Effects of exogenous methyl jasmonate and salicylic acid on rice resistance to Oebalus pugnax. Pest Manag. Sci. 2019, 75, 744–752. [Google Scholar] [CrossRef]
- Hayat, S.; Ali, B.; Ahmad, A. Salicylic acid: Biosynthesis, metabolism and physiological role in plants. In Salicylic Acid: A Plant Hormone; Springer: Berlin/Heidelberg, Germany, 2007; pp. 1–14. [Google Scholar]
- Valverde, J.M.; Giménez, M.J.; Guillen, F.; Valero, D.; Martinez-Romero, D.; Serrano, M. Methyl salicylate treatments of sweet cherry trees increase antioxidant systems in fruit at harvest and during storage. Postharvest Biol. Technol. 2015, 109, 106–113. [Google Scholar] [CrossRef]
- Forouhar, F.; Yang, Y.; Kumar, D.; Chen, Y.; Fridman, E.; Park, S.W.; Tong, L. Structural and biochemical studies identify tobacco SABP2 as a methyl salicylate esterase and implicate it in plant innate immunity. Proc. Natl. Acad. Sci. USA 2005, 102, 1773–1778. [Google Scholar] [CrossRef] [Green Version]
- Park, S.W.; Kaimoyo, E.; Kumar, D.; Mosher, S.; Klessig, D.F. Methyl salicylate is a critical mobile signal for plant systemic acquired resistance. Science 2007, 318, 113–116. [Google Scholar] [CrossRef]
- Gačnik, S.; Veberič, R.; Hudina, M.; Koron, D.; Mikulič-Petkovšek, M. Salicylate treatment affects fruit quality and also alters the composition of metabolites in strawberries. Horticulturae 2021, 7, 400. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Jaafar, H.; Karimi, E. Involvement of salicylic acid on antioxidant and anticancer properties, anthocyanin production and chalcone synthase activity in ginger (Zingiber officinale Roscoe) varieties. Int. J. Mol. Sci. 2012, 13, 14828–14844. [Google Scholar] [CrossRef] [PubMed]
- Giménez, M.J.; Valverde, J.M.; Valero, D.; Guillén, F.; Martínez-Romero, D.; Serrano, M.; Castillo, S. Quality and antioxidant properties on sweet cherries as affected by preharvest salicylic and acetylsalicylic acids treatments. Food Chem. 2014, 160, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Allan, A.C.; Wang, W.; Yin, X. The effects of salicylic acid on quality control of horticultural commodities. N. Z. J. Crop Hortic. Sci. 2022, 50, 99–117. [Google Scholar] [CrossRef]
- Gacnik, S.; Veberič, R.; Hudina, M.; Marinovic, S.; Halbwirth, H.; Mikulič-Petkovšek, M. Salicylic and methyl salicylic acid affect quality and phenolic profile of apple fruits three weeks before the harvest. Plants 2021, 10, 1807. [Google Scholar] [CrossRef]
- Gacnik, S.; Veberic, R.; Marinovic, S.; Halbwirth, H.; Mikulic-Petkovsek, M. Effect of pre-harvest treatments with salicylic and methyl salicylic acid on the chemical profile and activity of some phenylpropanoid pathway related enzymes in apple leaves. Sci. Hortic. 2021, 277, 109794. [Google Scholar] [CrossRef]
- Filgueiras, C.C.; Martins, A.D.; Pereira, R.V.; Willett, D.S. The Ecology of salicylic acid signaling: Primary, secondary and tertiary effects with applications in agriculture. Int. J. Mol. Sci. 2019, 20, 5851. [Google Scholar] [CrossRef] [Green Version]
- Joyce, D.C.; Wearing, H.; Coates, L.; Terry, L. Effects of phosphonate and salicylic acid treatments on anthracnose disease development and ripening of ’Kensington Pride’ mango fruit. Aust. J. Exp. Agric. 2001, 41, 805–813. [Google Scholar]
- Yao, H.; Tian, S. Effects of pre- and post-harvest application of salicylic acid or methyl jasmonate on inducing disease resistance of sweet cherry fruit in storage. Postharvest Biol. Technol. 2005, 35, 253–262. [Google Scholar] [CrossRef]
- Abbasi, P.A.; Ali, S.; Braun, G.; Bevis, E.; Fillmore, S. Reducing apple scab and frogeye or black rot infections with salicylic acid or its analogue on field-established apple trees. Can. J. Plant Pathol. 2019, 41, 345–354. [Google Scholar] [CrossRef]
- Halim, V.A.; Eschen-Lippold, L.; Altmann, S.; Birschwilks, M.; Scheel, D.; Rosahl, S. Salicylic acid is important for basal defense of Solanum tuberosum against Phytophthora infestans. Mol. Plant-Microbe Interact. 2007, 20, 1346–1352. [Google Scholar] [CrossRef] [Green Version]
- Da Rocha Neto, A.C.; Luiz, C.; Maraschin, M.; Di Piero, R.M. Efficacy of salicylic acid to reduce Penicillium expansum inoculum and preserve apple fruits. Int. J. Food Microbiol. 2016, 221, 54–60. [Google Scholar] [CrossRef]
- Singh, D.; Sharma, R.R. Postharvest Diseases of Fruits and Vegetables and Their Management in Postharvest Disinfection of Fruits and Vegetables; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1–52. [Google Scholar] [CrossRef]
- Martini, C.; Mari, M. Monilinia fructicola, Monilinia laxa (Monilinia Rot, Brown Rot). In Postharvest Decay; Elsevier: Amsterdam, The Netherlands, 2014; pp. 233–265. [Google Scholar] [CrossRef]
- Gačnik, S.; Munda, A.; Mikulič-Petkovšek, M. Effect of Salicylic and Methyl-Salicylic Acid on Mycelial Growth of Different Fungi and on Infection of Apple Fruits with Monilinia laxa. In Proceedings of the 14th Slovenian Conference on Plant Protection with International Participation, Maribor, Slovenia, 5–6 March 2019; pp. 513–518. [Google Scholar]
- Zhang, H.; Ma, L.; Wang, L.; Jiang, S.; Dong, Y.; Zheng, X. Biocontrol of gray mold decay in peach fruit by integration of antagonistic yeast with salicylic acid and their effects on postharvest quality parameters. Biol. Control 2008, 47, 60–65. [Google Scholar] [CrossRef]
- Wang, Y.-Y.; Li, B.-Q.; Qin, G.-Z.; Li, L.; Tian, S.-P. Defense response of tomato fruit at different maturity stages to salicylic acid and ethephon. Sci. Hortic. 2011, 129, 183–188. [Google Scholar] [CrossRef]
- Kapulnik, Y.; Yalpani, N.; Raskin, I. Salicylic acid induces cyanide-resistant respiration in tobacco cell-suspension cultures. Plant Physiol. 1992, 100, 1921–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Silva, H.; Klessig, D.F. Active oxygen species in the induction of plant systemic acquired resistance by salicylic acid. Science 1993, 262, 1883–1886. [Google Scholar] [CrossRef] [PubMed]
- Scaramuzza, N.; Cigarini, M.; Mutti, P.; Berni, E. Sanitization of packaging and machineries in the food industry: Effect of hydrogen peroxide on ascospores and conidia of filamentous fungi. Int. J. Food Microbiol. 2020, 316, 108421. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha Neto, A.C.; Maraschin, M.; Di Piero, R.M. Antifungal activity of salicylic acid against Penicillium expansum and its possible mechanisms of action. Int. J. Food Microbiol. 2015, 215, 64–70. [Google Scholar] [CrossRef]
- Lattanzio, V.; Lattanzino, V.M.T.; Cardinali, A. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. Phytochem. Adv. Res. 2006, 661, 23–67. [Google Scholar]
- Nicholson, R.L.; Hammerschmidt, R. Phenolic compounds and their role in disease resistance. Annu. Rev. Phytopathol. 1992, 30, 369–389. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Slatnar, A.; Stampar, F.; Veberic, R. Phenolic compounds in apple leaves after infection with apple scab. Biol. Plant. 2011, 55, 725. [Google Scholar] [CrossRef]
- Rusjan, D.; Veberič, R.; Mikulič-Petkovšek, M. The response of phenolic compounds in grapes of the variety ‘Chardonnay’ (Vitis vinifera L.) to the infection by phytoplasma Bois noir. Eur. J. Plant Pathol. 2012, 133, 965–974. [Google Scholar] [CrossRef]
- Gutha, L.R.; Casassa, L.F.; Harbertson, J.F.; Naidu, R.A. Modulation of flavonoid biosynthetic pathway genes and anthocyanins due to virus infection in grapevine (Vitis vinifera L.) leaves. BMC Plant Biol. 2010, 10, 187. [Google Scholar] [CrossRef] [Green Version]
- Kouam, J.C.D.; Ndjaga, J.M.; Akoa, S.P.; Ondobo, M.L.; Onomo, P.E.; Djocgoue, P.F.; Niemenak, N.; Collin, S. Flavan-3-ol and flavonol analysis in healthy and infected parents and progenies of cocoa leaves (Theobroma cacao L.) with Phytophthora megakarya Bras. and Grif. Trop. Plant Pathol. 2022, 47, 646–658. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Schmitzer, V.; Jakopic, J.; Cunja, V.; Veberic, R.; Munda, A.; Stampar, F. Phenolic compounds as defence response of pepper fruits to Colletotrichum coccodes. Physiol. Mol. Plant Pathol. 2013, 84, 138–145. [Google Scholar] [CrossRef]
- Slatnar, A.; Mikulic-Petkovsek, M.; Halbwirth, H.; Stampar, F.; Stich, K.; Veberic, R. Enzyme activity of the phenylpropanoid pathway as a response to apple scab infection. Ann. Appl. Biol. 2010, 156, 449–456. [Google Scholar] [CrossRef]
- Shirley, W.B. Flavonoid biosynthesis: ‘new’ functions for an ‘old’ pathway. Trends Plant Sci. 1996, 1, 377–382. [Google Scholar] [CrossRef]
- Feucht, W.; Treutter, D.; Schwalb, P. Principles of barrier formation of scab-infected apple fruits. Z. Pflanzenkrankh. Pflanzenschutz. 1998, 105, 394–403. [Google Scholar]
- Fang, F.; Huang, W.-D. Salicylic acid modulated flavonol biosynthesis in three key phases during grape berry development. Eur. Food Res. Technol. 2013, 237, 441–448. [Google Scholar] [CrossRef]
- Lee, M.H.; Bostock, R.M. Fruit exocarp phenols in relation to quiescence and development of Monilinia fructicola infections in Prunus spp.: A role for cellular redox? Phytopathology 2007, 97, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Villarino, M.; Sandin-Espana, P.; Melgarejo, P.; De Cal, A. High chlorogenic and neochlorogenic acid levels in immature peaches reduce Monilinia laxa infection by interfering with fungal melanin biosynthesis. J. Agric. Food Chem. 2011, 59, 3205–3213. [Google Scholar] [CrossRef]
- Gununu, P.R.; Munhuweyi, K.; Obianom, P.C.; Sivakumar, D. Assessment of eleven South African peach cultivars for susceptibility to brown rot and blue mould. Sci. Hortic. 2019, 254, 1–6. [Google Scholar] [CrossRef]
- Obi, V.I.; Montenegro, J.; Barriuso, J.J.; Saidani, F.; Aubert, C.; Gogorcena, Y. Is the tolerance of commercial peach cultivars to brown rot caused by Monilinia laxa modulated by its antioxidant content? Plants 2020, 9, 589. [Google Scholar] [CrossRef] [PubMed]
- Desikan, R.; Reynolds, A.; Hancock, J.; Neill, S. Harpin and hydrogen peroxide both initiate programmed cell death but have differential effects on defence gene expression in Arabidopsis suspension cultures. Biochem. J. 1998, 330, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorey, S.; Baillieul, F.; Pierrel, M.-A.; Saindrenan, P.; Fritig, B.; Kauffmann, S. Spatial and temporal induction of cell death, defense genes, and accumulation of salicylic acid in tobacco leaves reacting hypersensitively to a fungal glycoprotein elicitor. Mol. Plant-Microbe Interact. 1997, 10, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Dempsey, D.A.; Vlot, A.C.; Wildermuth, M.C.; Klessig, D.F. Salicylic acid biosynthesis and metabolism. Arab. Book 2011, 9, e0156. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Used Standards | Obtained by | |
|---|---|---|
| Flavanols | procyanidin B1 | Fluka Chemie |
| catechin | Fluka Chemie | |
| epicatechin | Fluka Chemie | |
| Hydroxycinnamic acids | p-coumaric acid | Fluka Chemie |
| sinapic acid | Fluka Chemie | |
| chlorogenic acid | Sigma-Aldrich | |
| 4-caffeoylquinic acid | Sigma-Aldrich | |
| Flavonols | quercetin-3-rutinoside | Fluka Chemie |
| quercetin-3-galactoside | Fluka Chemie | |
| quercetin-3-glucoside | Fluka Chemie | |
| quercetin-3-xyloside | Fluka Chemie | |
| kaempferol-3-glucoside | Fluka Chemie | |
| quercetin-3-arabinopyranoside | Apin Chemicals LTD | |
| quercetin-3-arabinofuranoside | Apin Chemicals LTD | |
| isorhamnetin-3-glucoside | Extrasynthèse | |
| quercetin-3-rhamnoside | Sigma-Aldrich | |
| Didydrochalcones | phloretin | Fluka Chemie |
| phloridzin | Fluka Chemie |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gacnik, S.; Munda, A.; Veberic, R.; Hudina, M.; Mikulic-Petkovsek, M. Preventive and Curative Effects of Salicylic and Methyl Salicylic Acid Having Antifungal Potential against Monilinia laxa and the Development of Phenolic Response in Apple Peel. Plants 2023, 12, 1584. https://doi.org/10.3390/plants12081584
Gacnik S, Munda A, Veberic R, Hudina M, Mikulic-Petkovsek M. Preventive and Curative Effects of Salicylic and Methyl Salicylic Acid Having Antifungal Potential against Monilinia laxa and the Development of Phenolic Response in Apple Peel. Plants. 2023; 12(8):1584. https://doi.org/10.3390/plants12081584
Chicago/Turabian StyleGacnik, Sasa, Alenka Munda, Robert Veberic, Metka Hudina, and Maja Mikulic-Petkovsek. 2023. "Preventive and Curative Effects of Salicylic and Methyl Salicylic Acid Having Antifungal Potential against Monilinia laxa and the Development of Phenolic Response in Apple Peel" Plants 12, no. 8: 1584. https://doi.org/10.3390/plants12081584
APA StyleGacnik, S., Munda, A., Veberic, R., Hudina, M., & Mikulic-Petkovsek, M. (2023). Preventive and Curative Effects of Salicylic and Methyl Salicylic Acid Having Antifungal Potential against Monilinia laxa and the Development of Phenolic Response in Apple Peel. Plants, 12(8), 1584. https://doi.org/10.3390/plants12081584