
Shrub Cover and Soil Moisture Affect Taxus baccata L. Regeneration at Its Southern Range

 , ,
, ,  ,
,  ,
, 
Abstract
1. Introduction
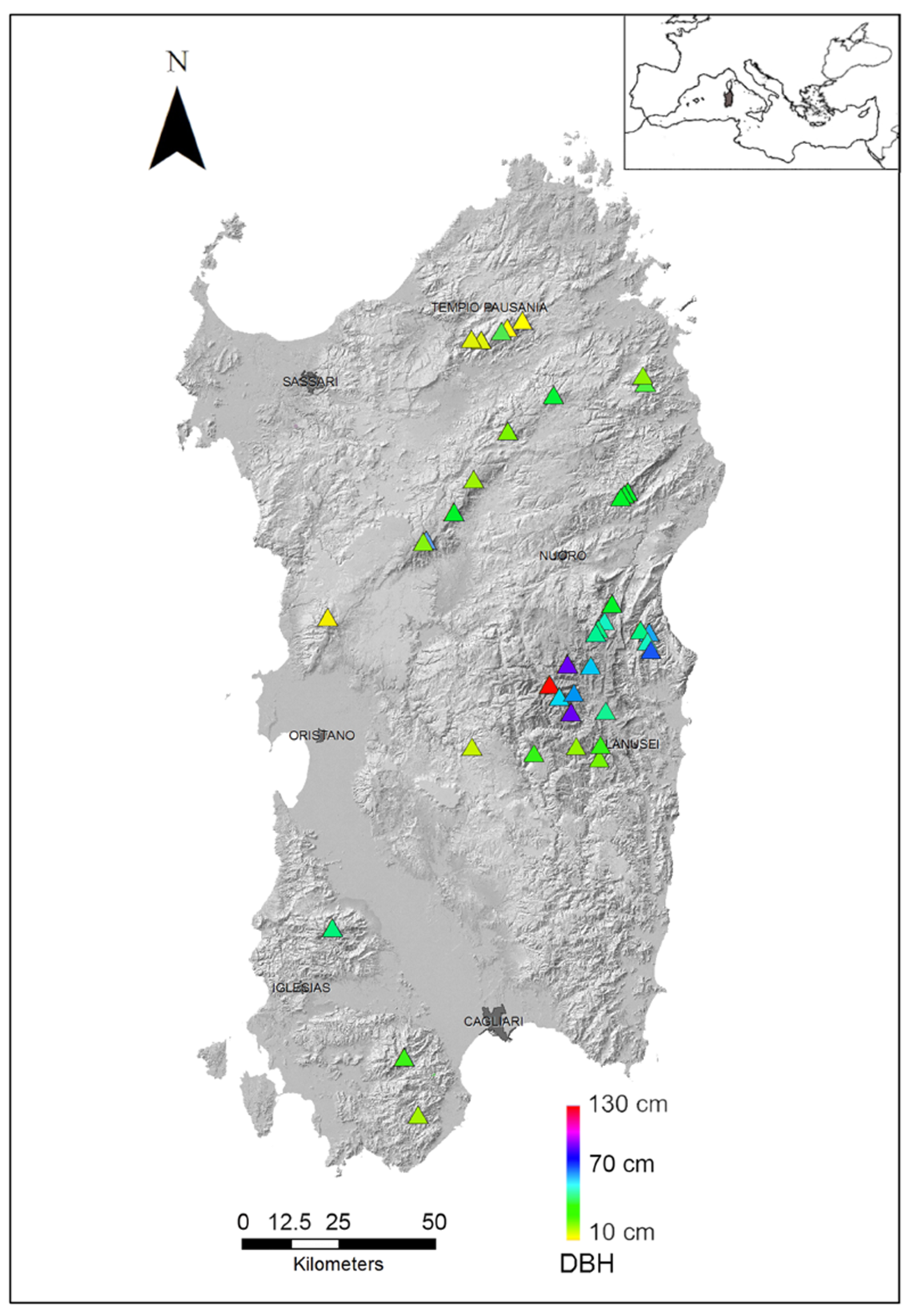
2. Results
3. Discussion
3.1. Main Factors Affecting Yew Recruitment
3.2. Implications for Conservation
4. Materials and Methods
4.1. Study System
4.2. Data Collection
4.3. Factors Affecting Yew Recruitment
- (1)
- Wood age; according to previous studies, yews start reproducing at ages comprised between 35 and 70 years [11]; consequently, the youngest formations are not capable of an efficient system of sexual reproduction. On the contrary, older trees can produce abundant quantities of pollen and seeds, allowing easier dispersal through frugivorous animals. Wood age was approximated by average diameter at breast height (aDBH), measured on as many as possible (often all) individuals counted per site. We selected sites with average DBH ranging from 13 to 130 cm, to infer possible differences between young formations and older stands.
- (2)
- Sex ratio [SR = Females/(Females + Males)]; the reproductive ecology of dioecious species is important in the understanding of different dynamics related to the spread of such species [17,66]. The importance of sex ratio in the analysis of population evolution is related to the fact that in many dioecious species, male prevalence was highlighted, especially under stressful conditions [82]. Females are subject to higher stresses, due to the major effort put on the reproductive phases, resulting in a diminished structural increase and higher mortality when under stress [83,84,85]. Plant populations characterized by wind pollination and abiotic dispersal were found to have more often male-biased sex ratios [86]. Recent studies showed how the growth rate of T. baccata females was lower than that of males, together with a higher water request for females [66,85,87]. A Mediterranean climate regime would favor male prevalence more than other European regions [17]. Moreover, it was supposed that older populations were male-sex-biased [17,60,66]. Consequently, we supposed that sex-biased populations were less capable of reproduction. The sex ratio was detected by analyzing all possible individuals in the field, through the observation of pollen strobiles on male trees and arils on females. Although pollen and arils occur in different periods of the year, the contemporary observation of sexual elements in the same populations was possible by searching their remnants both on the branches and on the ground, under the trees’ canopies. In the case of larger populations, more checks were carried out.
- (3)
- Wood closure (Clos); shading could affect seed germination and seedling establishment [17,32]. The structure of older yew stands could affect yew regeneration by impeding or reducing the growth of seedlings, mostly where canopies of conspecific trees do not allow a viable light quantity [17,22,32,88]. The closure was estimated as % ground cover of canopy projections.
- (4)
- Browsing (Bro = yes or no); the action of herbivores is well known to negatively affect the survival and establishment of seedlings, and thus the regeneration of yews [39,64]. Browsing was assessed by recording any evidence related to current impacts attributable to livestock and wild animals and/or their signs (e.g., physical presence observed in the field, excrements, tracks, fur/wool on the bark and on branches, decortication by deers’ antlers, etc.).
- (5)
- Shrub cover (Shrub + Spi); shrubs could protect seedlings from drought and browsing, thus promoting the regeneration of trees [38,39]. Here, total shrub cover was measured as the sum of abundance indexes of spiny shrubs (Spi, ranging from 0 to 5) and other shrubs (Shr, 0–5), meaning 0 = absent; 1 = very rare; 2 = rare; 3 = sporadic; 4 = frequent; and 5 = abundant.
- (6)
- Soil summer moisture (Ssmo = Yes or No); Mediterranean habitats are characterized by more or less long dry periods that could negatively affect yew recruitment [11]. Water availability has been considered a limiting factor for yew regeneration in the southern range of the species [53]. Moreover, it has been demonstrated that drought stress reduces leaf size and chlorophyll content, whereas it increases secondary metabolites and antioxidants, and their negative effect on plant morphology (stem length, leaf size, vegetative growth) and physiology (Pn, gs) [89]. Ssmo was assessed by observing in the field the soil moisture of the sites at the peak of the dry season.
- (7)
- Site morphology (Morph = Slope or Watercourses); rockiness and exposure to climatic factors (sun irradiation, winds, precipitations, frost) could affect microhabitat features, particularly in relation to microclimate (dryness and moisture). In particular, watercourses have higher moisture, and it is supposed that they allow a better regeneration than slopes [11]. Half of the selected sites (20) were chosen at dry conditions (slopes) and 20 further sites at moist localities (streams, gorges, springs).
- (8)
- Site slope (Pend); declivity was supposed to be important for the persistence of more natural conditions of the wooded formations [79] and for longer-lasting moisture [52]. Slopes were also found to be potentially relevant for seed dispersal in combination with water in some yew populations [90]. Site slope was quantified as average declivity and was calculated as the average of different slopes detected using an inclinometer.
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Malone, C. The Italian Neolithic: A synthesis of research. J. World Prehistoric. 2003, 17, 235–312. [Google Scholar] [CrossRef]
- Falcucci, A.; Maiorano, L.; Boitani, L. Changes in land-use/land-cover patterns in Italy and their implications for biodiversity conservation. Landsc. Ecol. 2007, 22, 617–631. [Google Scholar] [CrossRef]
- Blondel, J.; Aronson, J.; Bodiou, J.Y.; Boeuf, G. The Mediterranean Region: Biological Diversity in Space and Time, 2nd ed.; Oxford University Press: Oxford, UK, 2010; pp. 1–376. [Google Scholar]
- Puddu, G.; Falcucci, A.; Maiorano, L. Forest changes over a century in Sardinia: Implications for conservation in a Mediterranean hotspot. Agrofor. Syst. 2012, 85, 319–330. [Google Scholar] [CrossRef]
- Hoekstra, J.M.; Boucher, T.M.; Ricketts, T.H.; Roberts, C. Confronting a biome crisis: Global disparities of habitat loss and protection. Ecol. Lett. 2005, 8, 23–29. [Google Scholar] [CrossRef]
- Blasi, C.; Marchetti, M.; Chiavetta, U.; Aleffi, M.; Audisio, P.; Azzella, M.M.; Brunialti, G.; Capotorti, G.; Del Vico, E.; Lattanzi, E.; et al. Multi-taxon and forest structure sampling for identification of indicators and monitoring of old-growth forest. Plant Biosyst. 2010, 144, 160–170. [Google Scholar] [CrossRef]
- Sabatini, F.M.; Burrascano, S.; Keeton, W.S.; Levers, C.; Lindner, M.; Pötzschner, F.; Verkerk, P.J.; Bauhus, J.; Buchwald, E.; Chaskovsky, O.; et al. Where are Europe’s last primary forests? Divers. Distrib. 2018, 24, 1426–1439. [Google Scholar] [CrossRef]
- Kunzmann, L.; Mai, D.H. Die koniferen der Mastixioideen-Flora von Wiesabei Kamenz (Sachsen, Miozän) unter besonderer Berücksichtigung der Nadelblätter. Palaeontogr. Abt. B 2005, 272, 67–135. [Google Scholar] [CrossRef]
- Ellenberg, H. Vegetation Ecology of Central Europe, 4th ed.; Cambridge University Press: Cambridge, UK, 2009; pp. 1–731. [Google Scholar]
- Thomas, P.A.; Polwart, A. Taxus baccata L. J. Ecol. 2003, 91, 489–524. [Google Scholar]
- Schirone, B.; Caetano Ferreira, R.; Vessella, F.; Schirone, A.; Piredda, R.; Simeone, M.C. Taxus baccata in the Azores: A relict form at risk of imminent extinction. Biodivers. Conserv. 2010, 19, 1547–1565. [Google Scholar] [CrossRef]
- Farris, E.; Fenu, G.; Bacchetta, G. Mediterranean Taxus baccata woodlands in Sardinia: A characterization of the EU priority habitat 9580. Phytocoenologia 2012, 41, 231–246. [Google Scholar] [CrossRef]
- Serra Laliga, L.; García Martí, X. Distribución del tejo (Taxus baccata L.) en España. Ann. Deleg. Garrotxa ICHN 2012, 4, 17–42. [Google Scholar]
- Romo, A.; Iszkuło, G.; Seghir Taleb, M.; Walas, Ł.; Boratyński, A. Taxus baccata in Morocco: A tree in regression in its southern extreme. Dendrobiology 2017, 78, 63–74. [Google Scholar] [CrossRef]
- Kalantari, Z.; Heydari, M.; Esmailzadeh, O.; Asadi, H.; Prévosto, B. Classification of plant communities in the Caspian Hyrcanian English yew (Taxus baccata L.) forests using environmental factors: Testing the modified TWINSPAN method. J. Wildl. Biodivers. 2022, 6, 57–78. [Google Scholar] [CrossRef]
- Vessella, F.; Salis, A.; Scirè, M.; Piovesan, G.; Schirone, B. Natural regeneration and gender-specific spatial pattern of Taxus baccata in an old-growth population in Foresta Umbra (Italy). Dendrobiology 2015, 73, 75–90. [Google Scholar] [CrossRef]
- Karami-Kordalivand, P.; Esmailzadeh, O.; Willner, W.; Noroozi, J.; Alavi, S.J. Classification of forest communities (co-)dominated by Taxus baccata in the Hyrcanian forests (northern Iran) and their comparison with southern Europe. Eur. J. For. Res. 2021, 140, 463–476. [Google Scholar] [CrossRef]
- Deforce, K.; Bastiaens, J. The Holocene history of Taxus baccata (yew) in Belgium and neighbouring regions. Belg. J. Bot. 2007, 140, 222–237. [Google Scholar]
- Bennett, K.D.; Tzedakis, P.C.; Willis, K.J. Quaternary refugia of north European trees. J. Biogeogr. 1991, 18, 103–115. [Google Scholar] [CrossRef]
- García, D.; Zamora, R.; Hódar, J.A.; Gómez, J.M.; Castro, J. Yew (Taxus baccata L.) regeneration is facilitated by fleshy-fruited shrubs in Mediterranean environments. Biol. Conserv. 2000, 95, 31–38. [Google Scholar] [CrossRef]
- Tittensor, R.M. Ecological history of Yew Taxus baccata L. in southern England. Biol. Conserv. 1980, 17, 243–265. [Google Scholar] [CrossRef]
- Svenning, J.-C.; Magård, E. Population ecology and conservation status of the last natural population of English yew Taxus baccata in Denmark. Biol. Conserv. 1999, 88, 173–182. [Google Scholar] [CrossRef]
- Dhar, A.; Ruprecht, H.; Klumpp, R.; Vacik, H. Stand structure and natural regeneration of English yew (Taxus baccata L.) at Stiwollgraben in Austria. Dendrobiology 2006, 56, 19–26. [Google Scholar]
- Ruprecht, H.; Dhar, A.; Aigner, B.; Oitzinger, G.; Klumpp, R.; Vacik, H. Structural diversity of yew (Taxus baccata L.) populations. Eur. J. For. Ecol. 2010, 129, 189–198. [Google Scholar] [CrossRef]
- Iszkuło, G.; Pers-Kamczyc, E.; Nalepka, D.; Rabska, M.; Walas, Ł.; Dering, M. Postglacial migration dynamics helps to explain current scattered distribution of Taxus baccata. Dendrobiology 2016, 76, 81–89. [Google Scholar] [CrossRef]
- Lewandowski, A.; Burczyk, J.; Mejnartowicz, L. Genetic structure of English yew (Taxus baccata L.) in the Wierzchlas Reserve: Implications for genetic conservation. For. Ecol. Manag. 1995, 73, 221–227. [Google Scholar] [CrossRef]
- Iszkuło, G. The yew (Taxus baccata L.) of the Cisowy Jar reserve near Olecko. Dendrobiology 2001, 46, 33–37. [Google Scholar]
- Iszkuło, G. Influence of biotic and abiotic factors on natural regeneration of European yew (Taxus baccata L.): A review. Span. J. Rural Dev. 2011, 2, 1–6. [Google Scholar] [CrossRef]
- Iszkuło, G.; Didukh, Y.; Giertych, M.; Jasinska, A.K.; Sobierajska, K.; Szmyt, J. Weak competitive ability may explain decline of Taxus baccata. Ann. For. Sci. 2012, 69, 705–712. [Google Scholar] [CrossRef]
- Iszkuło, G.; Nowak-Dyjeta, K.; Sękiewicz, M. Influence of initial light intensity and deer browsing on Taxus baccata saplings: A six years field study. Dendrobiology 2014, 71, 93–99. [Google Scholar] [CrossRef]
- Devaney, J.L.; Whelan, P.M.; Jansen, M.A.K. Light responses of yew (Taxus baccata L.); does size matter? Trees 2015, 29, 109–118. [Google Scholar] [CrossRef]
- Garbarino, M.; Weisberg, P.J.; Bagnara, L.; Urbinati, C. Sex-related spatial segregation along environmental gradients in the dioecious conifer, Taxus baccata. For. Ecol. Manag. 2015, 358, 122–129. [Google Scholar] [CrossRef]
- Camprodon, J.; Guixé, D.; Casals, P.; Caritat, A.; Buqueras, X.; García-Martí, X.; Reverté, J.; Rios, A.I.; Beltrán, M.; Llovet, J.; et al. Conservation of Mediterranean Yew Forests. Best Practice Handbook. Life TAXUS Project; Forest Science Centre of Catalonia: Solsona, Spain, 2016; pp. 1–142. [Google Scholar]
- Busing, R.T.; Halpern, C.B.; Spies, T.A. Ecology of Pacific yew (Taxus brevifolia) in Western Oregon and Washington. Conserv. Biol. 1995, 9, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Piovesan, G.; Presutti Saba, E.; Biondi, F.; Alessandrini, A.; Di Filippo, A.; Schirone, B. Population ecology of yew (Taxus baccata L.) in the Central Apennines: Spatial patterns and their relevance for conservation strategies. Plant Ecol. 2009, 205, 23–46. [Google Scholar] [CrossRef]
- Hulme, P.E. Natural regeneration of yew (Taxus baccata L.): Microsite, seed or herbivore limitation? J. Ecol. 1996, 84, 853–861. [Google Scholar] [CrossRef]
- García, D.; Obeso, J.R. Facilitation by herbivore-mediated nurse plants in a threatened tree, Taxus baccata: Local effects and landscape level consistency. Ecography 2003, 26, 739–750. [Google Scholar] [CrossRef]
- Farris, E.; Filigheddu, R. Effects of browsing in relation to vegetation cover on common yew (Taxus baccata L.) recruitment in Mediterranean environments. Plant Ecol. 2008, 199, 309–318. [Google Scholar] [CrossRef]
- Hageneder, F. Yew: A History; The History Press Ltd.; Sutton Publishing: London, UK, 2007. [Google Scholar]
- Ballian, D.; Memišević Hodžić, M. Preservation of Taxus baccata L. in Bosnia and Herzegovina. In Conservación y Restauración de los Bosques de tejo en Europa; Ramil-Rego, P., Rodríguez Guitián, M.A., Ferreiro da Costa, J., Gómez-Orellana, L., Eds.; Monografías do Ibader, Serie Biodiversidade: Lugo, Spain, 2023; pp. 93–101. [Google Scholar]
- European Commission. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. L 1992, 206, 7–50. [Google Scholar]
- Biondi, E.; Casavecchia, S.; Pesaresi, S. Interpretation and management of the forest habitats of the Italian peninsula. Acta Bot. Gall. 2010, 157, 687–719. [Google Scholar] [CrossRef]
- Epstein, Y.; López-Bao, J.V.; Chapron, G. A legal-ecological understanding of favorable conservation status for species in Europe. Conserv. Lett. 2016, 9, 81–88. [Google Scholar] [CrossRef]
- Desole, L. Distribuzione dell'Ilex aquifolium L. e del Taxus baccata L. in Sardegna. Prima nota. Atti Soc. Toscana Sci. Nat. 1948, 55, 1–38. [Google Scholar]
- Desole, L. Distribuzione geografica dell'Ilex aquifolium L. e del Taxus baccata L. in Sardegna. Seconda e ultima nota. Bull. Ist. Bot. Sassari 1966, 7, 5–67. [Google Scholar]
- Bacchetta, G.; Farris, E. Studio fitosociologico, ecologico e corologico dei boschi di Taxus baccata L. in Sardegna (Italia). In I Jornadas Internacionales Sobre el Tejo y las Tejedas en el Mediterraneo Occidental; Serra, L., Ed.; Generalidad Valenciana: Alcoy, Spain, 2007; pp. 195–204. [Google Scholar]
- Camarda, I. Grandi Alberi e Foreste Vetuste Della Sardegna; Carlo Delfino Editore: Sassari, Italy, 2020. [Google Scholar]
- Calcagno, V.; de Mazancourt, C. glmulti: An R Package for Easy Automated Model Selection with (Generalized) Linear Models. J. Stat. Softw. 2010, 34, 1–29. [Google Scholar] [CrossRef]
- Debussche, M.; Isenmann, P. Bird-dispersed seed rain and seedling establishment in patchy Mediterranean vegetation. Oikos 1994, 69, 414–426. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; Zamora, R.; Castro, J.; Hódar, J.A. Facilitation of tree saplings by nurse plants: Microhabitat amelioration or protection against herbivores? J. Veg. Sci. 2008, 19, 161–172. [Google Scholar] [CrossRef]
- Sanz, R.; Pulido, F.; Nogués-Bravo, D. Predicting mechanisms across scales: Amplified effects of abiotic constraints on the recruitment of yew Taxus baccata. Ecography 2009, 3, 993–1000. [Google Scholar] [CrossRef]
- Linares, J.C. Shifting limiting factors for population dynamics and conservation status of the endangered English yew (Taxus baccata L., Taxaceae). For. Ecol. Manag. 2013, 291, 119–127. [Google Scholar] [CrossRef]
- Casals, P.; Camprodon, J.; Caritat, A.; Rios, A.I.; Guixé, D.; García-Martí, X.; Martín-Alcón, S.; Coll, L. Forest structure of Mediterranean yew (Taxus baccata L.) populations and neighbor effects on juvenile yew performance in the NE Iberian Peninsula. For. Syst. 2015, 24, e042. [Google Scholar] [CrossRef]
- Casula, P.; Murgia, A. Selectivity and context dependence of Corsican red deer browsing in a Mediterranean coppice system. Hystrix 2017, 28, 157–164. [Google Scholar] [CrossRef]
- Fitzsimons, M.S.; Miller, R.M.; Jastrow, J.D. Scale-dependent niche axes of arbuscular mycorrhizal fungi. Oecologia 2008, 158, 117–127. [Google Scholar] [CrossRef]
- Richard, F.; Selosse, M.-A.; Gardes, M. Facilitated establishment of Quercus ilex in shrub-dominated communities within a Mediterranean ecosystem: Do mycorrhizal partners matter? FEMS Microbiol. Ecol. 2009, 68, 14–24. [Google Scholar] [CrossRef]
- López-García, Á.; Hempel, S.; Miranda, J.D.; Rillig, M.C.; Barea, J.M.; Azcón-Aguilar, C. The influence of environmental degradation processes on the arbuscular mycorrhizal fungal community associated with yew (Taxus baccata L.), an endangered tree species from Mediterranean ecosystems of Southeast Spain. Plant Soil 2013, 370, 355–366. [Google Scholar] [CrossRef]
- Balaguer-Romano, R.; Sainz-Ollero, H.; Vasco-Encuentra, F. Yew (Taxus baccata L.) population dynamics in the Iberian Mediterranean Mountains: Natural regeneration and expansion in East Central System (Spain). For. Syst. 2020, 29, eSC03. [Google Scholar] [CrossRef]
- Perrin, P.M.; Mitchell, F. Effects of shade on growth, biomass allocation and leaf morphology in European yew (Taxus baccata L.). Eur. J. For. Res. 2012, 132, 211–218. [Google Scholar] [CrossRef]
- Mendoza, I.; Zamora, R.; Castro, J. A seeding experiment for testing tree community recruitment under variable environments: Implications for forest regeneration and conservation in Mediterranean habitats. Biol. Conserv. 2009, 142, 1491–1499. [Google Scholar] [CrossRef]
- Marzolff, I.; Stephan, R.; Kirchhoff, M.; Seeger, M.; AïtHssaïne, A.; Ries, J.B. UAV-based classification of tree-browsing intensity in open woodlands. In Proceedings of the EGU General Assembly 2020, Online, 4–8 May 2020. [Google Scholar]
- Thomas, P.A.; García-Martí, X. Response of European yews to climate change: A review. For. Syst. 2015, 24, eR01. [Google Scholar] [CrossRef]
- Mysterud, A.; Østbye, E. Roe deer (Capreolus capreolus) browsing pressure affects yew (Taxus baccata) recruitment within nature reserves in Norway. Biol. Conserv. 2004, 120, 545–548. [Google Scholar] [CrossRef]
- Balaguer-Romano, R.; Fernández-González, P.; Alcaide, A.; Braschi, J.; Vasco-Encuentra, F. Short communication: The mosquito yew wood: Survey and analysis of Mediterranean Taxus baccata L. woods (Habitat 9580*). For. Syst. 2022, 31, eSC03. [Google Scholar] [CrossRef]
- Iszkuło, G.; Jasinska, A.K.; Giertych, M.J.; Boratynsky, A. Do secondary sexual dimorphism and female intolerance to drought influence the sex ratio and extinction risk of Taxus baccata? Plant Ecol. 2009, 200, 229–240. [Google Scholar] [CrossRef]
- Perrin, P.M.; Kelly, D.L.; Mitchell, F.J.G. Long-term deer exclusion in yew wood and oakwood habitats in southwest Ireland: Natural regeneration and stand dynamics. For. Ecol. Manag. 2006, 236, 356–367. [Google Scholar] [CrossRef]
- Calvia, G.; Ruggero, A. The vascular flora of Mount Limbara: From a troubled past to an uncertain future. Fl Medit 2020, 30, 293–313. [Google Scholar] [CrossRef]
- Manier, D.J.; Hobbs, N.T. Large herbivores influence the composition and diversity of shrub-steppe communities in the Rocky Mountains, USA. Oecologia 2006, 146, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Riedel, J.L.; Bernués, A.; Casasús, I. Livestock grazing impacts on herbage and shrub dynamics in a Mediterranean Natural Park. Rangel. Ecol. Manag. 2012, 66, 224–233. [Google Scholar] [CrossRef]
- Carmignani, L.; Oggiano, G.; Funedda, A.; Conti, P.; Pasci, S. The geological map of Sardinia (Italy) at 1:250,000 scale. J. Maps 2016, 12, 826–835. [Google Scholar] [CrossRef]
- Bacchetta, G.; Bagella, S.; Biondi, E.; Farris, E.; Filigheddu, R.; Mossa, L. Vegetazione forestale e serie di vegetazione della Sardegna (con rappresentazione cartografica alla scala 1:350.000). Fitosociologia 2009, 46, 3–82. [Google Scholar]
- Canu, S.; Rosati, L.; Fiori, M.; Motroni, A.; Filigheddu, R.; Farris, E. Bioclimate map of Sardinia (Italy). J. Maps 2015, 11, 711–718. [Google Scholar] [CrossRef]
- Barreca, F. La Sardegna Fenicia e Punica; Chiarella Editore: Sassari, Italy, 1974. [Google Scholar]
- Meloni, P. La Sardegna Romana; Chiarella Editore: Sassari, Italy, 1975. [Google Scholar]
- Pungetti, G. Anthropological approach to agricultural landscape history in Sardinia. Landsc. Urban Plan. 1985, 31, 47–56. [Google Scholar] [CrossRef]
- Caterini, F. Colpi di Scure e Sensi di Colpa: Storia del Disboscamento dalle Origini a Oggi; Carlo Delfino Editore: Sassari, Italy, 2013. [Google Scholar]
- Fois, M.; Fenu, G.; Bacchetta, G. Estimating land market values from real estate offers: A replicable method in support of biodiversity conservation strategies. AMBIO 2019, 48, 313–323. [Google Scholar] [CrossRef]
- Fantini, S.; Fois, M.; Casula, P.; Fenu, G.; Calvia, G.; Bacchetta, G. Structural heterogeneity and old-growthness: A first regional-scale assessment of Sardinian forests. Ann. For. Res. 2020, 63, 103–120. [Google Scholar] [CrossRef]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theuirillat, J.-P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; Gavilán García, R.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Angius, V. Geografia, storia e statistica dell’Isola di Sardegna, vol. 17 bis, 18 ter, 18 quarter. In Dizionario Geografico-Storico-Statistico-Commerciale Degli Stati di S. M. il Re di Sardegna; Casalis, G., Ed.; Maspero e Marzorati: Torino, Italy, 1851. [Google Scholar]
- Obeso, J.R. The costs of reproduction in plants. New Phytol. 2002, 155, 321–348. [Google Scholar] [CrossRef]
- Leigh, A.; Cosgrove, M.J.; Nicotra, A.B. Reproductive allocation in a gender dimorphic shrub: Anomalous female investment in Gynatrix pulchella? J. Ecol. 2006, 94, 1261–1271. [Google Scholar] [CrossRef]
- Massei, G.; Watkins, R.; Hartley, S.E. Sex-related growth and secondary compounds in Juniperus oxycedrus macrocarpa. Acta Oecol. 2006, 29, 135–140. [Google Scholar] [CrossRef]
- Rabska, M.; Giertych, M.J.; Nowak, K.; Pers-Kamczyc, E.; Iszkuło, G. Consequences of the Reproductive Effort of Dioecious Taxus baccata L. Females in a Generative Bud Removal Experiment—Important Role of Nitrogen in Female Reproduction. Int. J. Mol. Sci. 2022, 23, 14225. [Google Scholar] [CrossRef]
- Sinclair, J.P.; Emlen, J.; Freeman, D.C. Biased sex ratios in plants: Theory and trends. Bot. Rev. 2012, 78, 63–86. [Google Scholar] [CrossRef]
- Iszkuło, G.; Jasinska, A.K.; Sobierajska, K. Dendrological differences between Taxus baccata males and females in comparison with monoecious Abies alba. Dendrobiology 2011, 65, 55–61. [Google Scholar]
- Dovciak, M. Population dynamics of the endangered English yew (Taxus baccata L.) and its management implications for biosphere reserves of the Western Carpathians. In Young Scientist Award 2002 Research Study; U. MAB Secretariat Division of Ecological Sciences, Ed.; Department of Applied Ecology, Faculty of Ecology and Environmental Sciences, Zvolen Technical University: Banskaá Štiavnica, Slovakia, 2002; pp. 1–43. [Google Scholar]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Fernández-González, P.; Fernández-Morcuende, A.; García-Gomariz, E.; Rodríguez-Rivas, M.J.; Sánchez-Amador, E.; Vasco-Encuentra, F. Yew matriarchies of the Sierra de Francia. Dynamics and ecology of recently identified Yew populations in the Central Iberian Mountain Range (Sistema Central). For. Syst. 2015, 24, e043. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2020; ISBN 3-900051-07-0. Available online: http://www.R-project.org/ (accessed on 21 February 2021).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: Berlin/Heidelberg, Germany, 2002; pp. 1–515. [Google Scholar]
- Kent, J.T. Robust properties of likelihood ratio tests. Biometrika 1982, 69, 19–27. [Google Scholar] [CrossRef]
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c). R Package Version 2.2-2. 2019. [Google Scholar]
- Zeileis, A.; Hothorn, T. Diagnostic checking in regression relationships. R News 2002, 2, 7–10. Available online: https://CRAN.R-project.org/doc/Rnews/ (accessed on 21 February 2021).


{kind=link}
| Rank | Model Structure | K | AICc | ∆AICc | LL | Dev. Expl. |
|---|---|---|---|---|---|---|
| 1 | Recr ~ Shrub | 2 | 130.2244 | 0.0000 | −62.9501 | 0.6606 |
| 2 | Recr ~ SR | 2 | 183.2421 | 53.0176 | −89.4589 | 0.2877 |
| 3 | Recr ~ DBH | 2 | 187.7552 | 57.5308 | −91.7154 | 0.2560 |
| 4 | Recr ~ Morph | 2 | 209.8880 | 79.6635 | −102.7818 | 0.1003 |
| 5 | Recr ~ Bro | 2 | 211.1093 | 80.8849 | −103.3925 | 0.0917 |
| 6 | Recr ~ Ssmo | 2 | 213.5470 | 83.3225 | −104.6113 | 0.0746 |
| 7 | M0: Recr ~ 1 | 1 | 221.9372 | 91.7127 | −109.9159 | 0.0000 |
| 8 | Recr ~ Clos | 2 | 223.8796 | 93.6551 | −109.7776 | 0.0000 |
| 9 | Recr ~ Pend | 2 | 224.1561 | 93.9317 | −109.9159 | 0.0000 |
| Rank | Effects on Yew Regeneration | K | AICc | ∆AICc | wi | LL | Dev. Expl. |
|---|---|---|---|---|---|---|---|
| 1 | Shrub + Morph + Bro | 4 | 127.1372 | 0.0000 | 0.4248 | −58.9972 | 0.7162 |
| 2 | Shrub + Morph | 3 | 128.1617 | 1.0245 | 0.2545 | −60.7475 | 0.6916 |
| 3 | Shrub + DBH + Morph + Bro | 5 | 129.3459 | 2.2087 | 0.1408 | −58.7906 | 0.7191 |
| 4 | Shrub | 2 | 130.2244 | 3.0872 | 0.0907 | −62.9501 | 0.6606 |
| 5 | Shrub + DBH + Morph + Bro + Ssmo | 6 | 131.4385 | 4.3013 | 0.0494 | −58.4465 | 0.7240 |
| 6 | Shrub + Bro | 3 | 132.5451 | 5.4079 | 0.0284 | −62.9392 | 0.6608 |
| 7 | Shrub + SR + DBH + Morph + Bro + Ssmo | 7 | 134.3926 | 7.2554 | 0.0113 | −58.4463 | 0.7240 |
| 8 | Morph + Bro | 3 | 208.3445 | 81.2073 | 0.0000 | −100.8389 | 0.1277 |
| 9 | Morph | 2 | 209.8880 | 82.7508 | 0.0000 | −102.7818 | 0.1003 |
| 10 | Bro | 2 | 211.1093 | 83.9721 | 0.0000 | −103.3925 | 0.0917 |
| 11 | None | 1 | 221.9372 | 94.8000 | 0.0000 | −109.9159 | 0.0000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calvia, G.; Casula, P.; Farris, E.; Fenu, G.; Fantini, S.; Bacchetta, G. Shrub Cover and Soil Moisture Affect Taxus baccata L. Regeneration at Its Southern Range. Plants 2023, 12, 1819. https://doi.org/10.3390/plants12091819
Calvia G, Casula P, Farris E, Fenu G, Fantini S, Bacchetta G. Shrub Cover and Soil Moisture Affect Taxus baccata L. Regeneration at Its Southern Range. Plants. 2023; 12(9):1819. https://doi.org/10.3390/plants12091819
Chicago/Turabian StyleCalvia, Giacomo, Paolo Casula, Emmanuele Farris, Giuseppe Fenu, Sergio Fantini, and Gianluigi Bacchetta. 2023. "Shrub Cover and Soil Moisture Affect Taxus baccata L. Regeneration at Its Southern Range" Plants 12, no. 9: 1819. https://doi.org/10.3390/plants12091819
APA StyleCalvia, G., Casula, P., Farris, E., Fenu, G., Fantini, S., & Bacchetta, G. (2023). Shrub Cover and Soil Moisture Affect Taxus baccata L. Regeneration at Its Southern Range. Plants, 12(9), 1819. https://doi.org/10.3390/plants12091819