
A Combination of Three Genomic Regions Conditions High Level of Adult Plant Stripe Rust Resistance in Australian Wheat Cultivar Sentinel



Abstract
:1. Introduction
2. Results
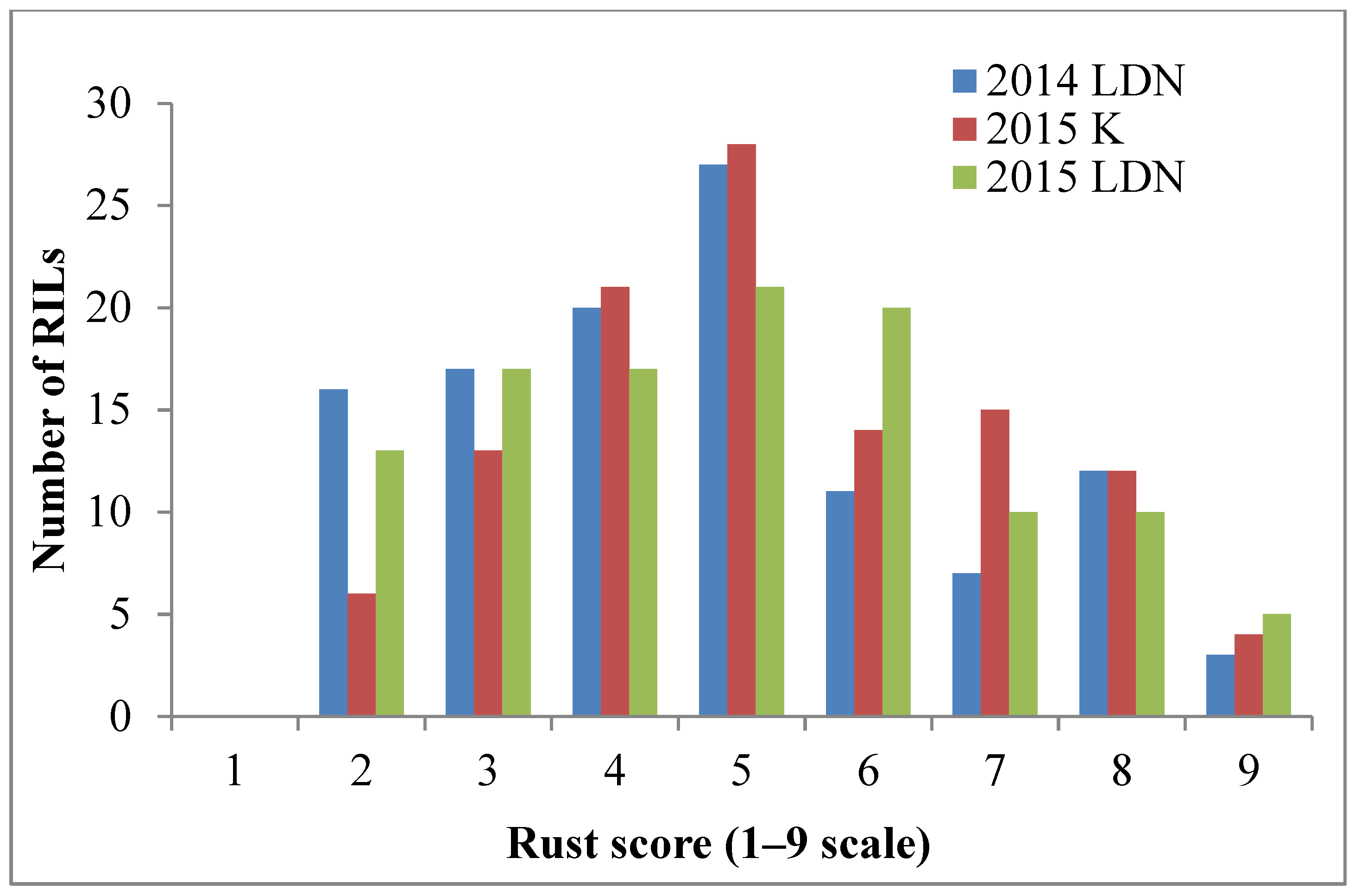
2.1. Stripe Rust Response Assessments
2.1.1. Field Assessment
2.1.2. Greenhouse Assessment
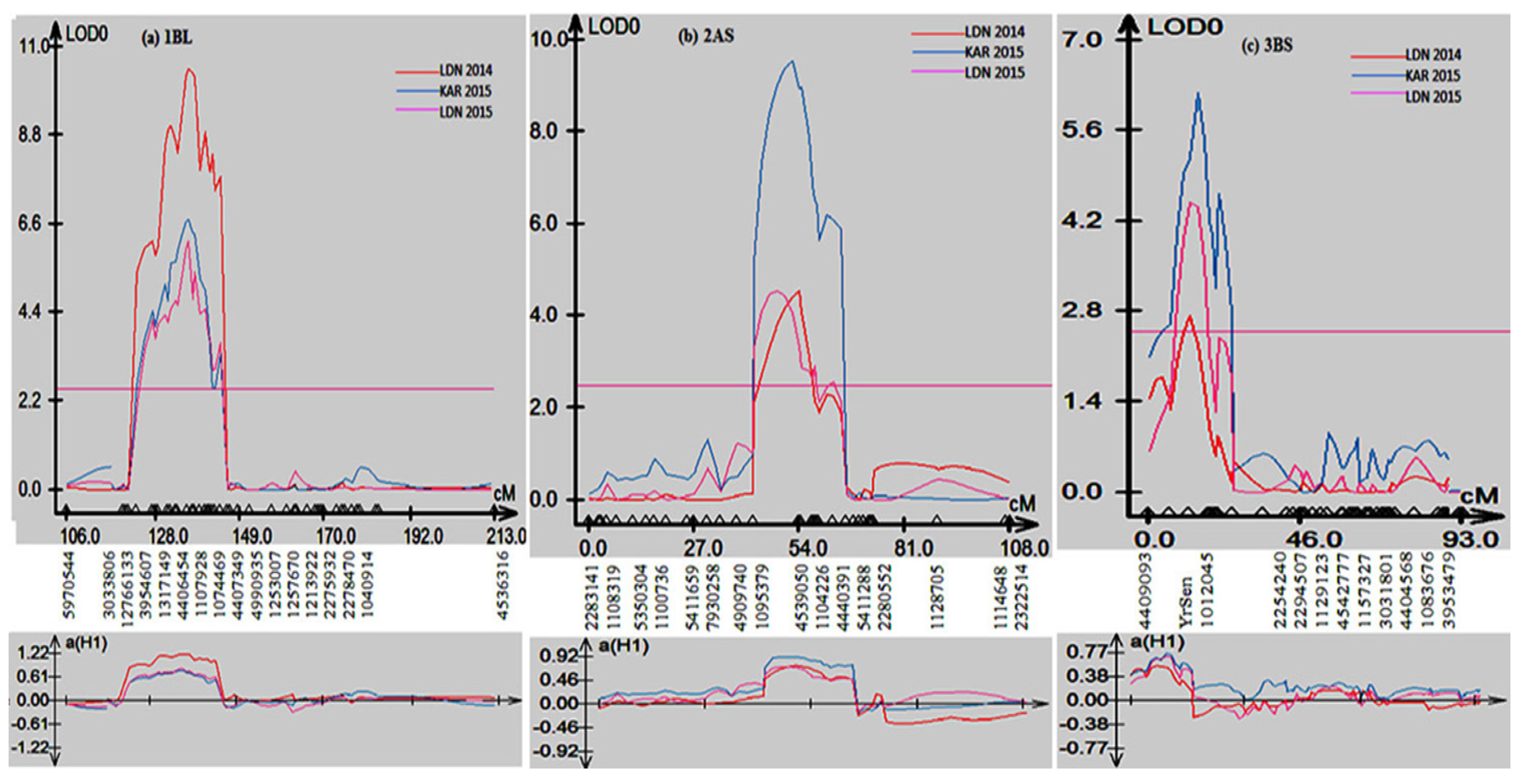
2.2. Construction of Linakge Map and QTL Analysis
2.2.1. Detection of APR Genes Yr18 and Yr29
2.2.2. Linkage Map Construction
2.2.3. QTL Analysis
2.2.4. Contribution of Sentinel Alleles
2.2.5. Mendelization of QYr.sun-2AS
2.2.6. Interaction of QTL
3. Discussion
4. Materials and Methods
4.1. Materials
4.1.1. Plant Material
4.1.2. Pathogen Materials
4.2. Stripe Rust Assessments
4.2.1. Field Assessment
4.2.2. Greenhouse Assessment
4.3. Molecular Mapping
4.3.1. DNA Extraction
4.3.2. Linkage Map Construction and QTL Analysis
4.4. Marker Screening for APR Genes Yr18 and Yr29
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wellings, C.R. Global status of stripe rust: A review of historical and current threats. Euphytica 2011, 179, 129–141. [Google Scholar] [CrossRef]
- Chen, X.M. Epidemiology and control of stripe rust [Puccinia striiformis f. sp. tritici] on wheat. Can. J. Plant Pathol. 2005, 27, 314–337. [Google Scholar] [CrossRef]
- Beddow, J.M.; Pardey, P.G.; Chai, Y.; Hurley, T.M.; Kriticos, D.J.; Braun, H.J.; Park, R.F.; Yonow, T. Research investment implications of shifts in the global geography of wheat stripe rust. Nat. Plants 2015, 1, 5132. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, L.; Brown, J.S.; Young, R.M.; Pascoe, I. Occurrence and distribution of wheat stripe rust in Victoria and susceptibility of commercial wheat cultivars. Austr. Plant Pathol. 1980, 9, 14. [Google Scholar] [CrossRef]
- Wellings, C.R.; Wright, D.G.; Keiper, F.; Loughman, R. First detection of wheat stripe rust in Western Australia: Evidence for a foreign incursion. Austr. Plant Pathol. 2003, 32, 321–322. [Google Scholar] [CrossRef]
- Wellings, C.R. Puccinia striiformis in Australia: A review of the incursion, evolution, and adaptation of stripe rust in the period 1979–2006. Austr. J. Agric. Res. 2007, 58, 567–575. [Google Scholar] [CrossRef]
- Ali, S.; Rodriguez-Algaba, J.; Thach, T.; Sørensen, C.K.; Hansen, J.G.; Lassen, P.; Nazari, K.; Hodson, D.P.; Justesen, A.F.; Hovmøller, M.S. Yellow rust epidemics worldwide were caused by pathogen races from divergent genetic lineages. Front. Plant Sci. 2017, 8, 1057. [Google Scholar] [CrossRef]
- Singh, R.P.; Singh, P.K.; Rutkoski, J.; Hodson, D.P.; He, X.; Jørgensen, L.N.; Hovmøller, M.S.; Huerta-Espino, J. Disease impact on wheat yield potential and prospects of genetic control. Annu. Rev. Phytopathol. 2016, 54, 303–322. [Google Scholar] [CrossRef]
- Carmona, M.; Sautua, F.; Pérez-Hérnandez, O.; Reis, E.M. Role of fungicide applications on the integrated management of wheat stripe rust. Front. Plant Sci. 2020, 11, 733. [Google Scholar] [CrossRef]
- Bariana, H.S.; Bansal, U.K.; Schmidt, A.; Lehmensiek, A.; Kaur, J.; Miah, H.; Howes, N.; McIntyre, C.L. Molecular mapping of adult plant stripe rust resistance in wheat and identification of pyramided QTL genotypes. Euphytica 2010, 176, 251–260. [Google Scholar] [CrossRef]
- Chen, X.M. Challenges and solutions for stripe rust control in the United States. Austr. J. Agric. Res. 2007, 58, 648–655. [Google Scholar] [CrossRef]
- Bariana, H.S. Breeding for disease resistance. In Encyclopedia of Applied Plant Sciences; Thomas, B., Murphy, D.J., Murray, B.G., Eds.; Academic Press: London, UK, 2003; pp. 244–253. [Google Scholar]
- Lagudah, E.S. Molecular genetics of race non-specific rust resistance in wheat. Euphytica 2011, 179, 81–91. [Google Scholar] [CrossRef]
- Bariana, H.S.; McIntosh, R.A. Genetics of adult plant stripe rust resistance in four Australian wheats and the French cultivar ‘Hybride-de-Bersee’. Plant Breed. 1995, 114, 485–491. [Google Scholar] [CrossRef]
- Singh, R.P.; Huerta-Espino, J.; Rajaram, S. Achieving near-immunity to leaf and stripe rusts in wheat by combining slow rusting resistance genes. Acta Phytopathol. Entomol. Hung. 2000, 35, 133–139. [Google Scholar]
- Bariana, H.S.; Bansal, U.K. Breeding for disease resistance. In Encyclopedia of Applied Plant Sciences; Brian, T., Brian, G.M., Dennis, M., Eds.; Academic Press: Waltham, MA, USA, 2017; Volume 3, pp. 69–76. [Google Scholar]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef]
- Varshney, R.K.; Nayak, S.N.; May, G.D.; Jackson, S.A. Next-generation sequencing technologies and their implications for crop genetics and breeding. Trends Biotechnol. 2009, 27, 522–530. [Google Scholar] [CrossRef]
- Rosewarne, G.M.; Herrera-Foessel, S.A.; Singh, R.P.; Huerta-Espino, J.; Lan, C.X.; He, Z.H. Quantitative trait loci of stripe rust resistance in wheat. Theor. Appl. Genet. 2013, 126, 2427–2449. [Google Scholar] [CrossRef]
- Bansal, U.K.; Kazi, A.G.; Singh, B.; Hare, R.A.; Bariana, H.S. Mapping of durable stripe rust resistance in a durum wheat cultivar Wollaroi. Mol. Breed. 2014, 33, 51–59. [Google Scholar] [CrossRef]
- Agenbag, G.M.; Pretorius, Z.A.; Boyd, L.A.; Bender, C.M.; Prins, R. Identification of adult plant resistance to stripe rust in the wheat cultivar Cappelle-Desprez. Theor. Appl. Genet. 2012, 125, 109–120. [Google Scholar] [CrossRef]
- Boukhatem, N.; Baret, P.V.; Mingeot, D.; Jacquemin, J.M. Quantitative trait loci for resistance against yellow rust in two wheat-derived recombinant inbred line populations. Theor. Appl. Genet. 2002, 104, 111–118. [Google Scholar] [CrossRef]
- Chhuneja, P.; Kaur, S.; Garg, T.; Ghai, M.; Kaur, S.; Prashar, M.; Bains, N.S.; Goel, R.K.; Keller, B.; Dhaliwal, H.S.; et al. Mapping of adult plant stripe rust resistance genes in diploid A genome wheat species and their transfer to bread wheat. Theor. Appl. Genet. 2008, 116, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Dedryver, F.; Paillard, S.; Mallard, S.; Robert, O.; Trottet, M.; Negre, S.; Verplancke, G.; Jahier, J. Characterization of genetic components involved in durable resistance to stripe rust in the bread wheat ‘Renan’. Phytopathology 2009, 99, 968–973. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Chen, Z.; Wang, Y.; Bland, D.; Buck, J.; Brown-Guedira, G.; Johnson, J. Characterization of a major QTL for adult plant resistance to stripe rust in US soft red winter wheat. Theor. Appl. Genet. 2011, 123, 1401–1411. [Google Scholar] [CrossRef] [PubMed]
- Mallard, S.; Gaudet, D.; Aldeia, A.; Abelard, C.; Besnard, A.L.; Sourdille, P.; Dedryver, F. Genetic analysis of durable resistance to yellow rust in bread wheat. Theor. Appl. Genet. 2005, 110, 1401–1409. [Google Scholar] [CrossRef] [PubMed]
- Randhawa, M.S.; Bariana, H.S.; Mago, R.; Bansal, U.K. Mapping of a new stripe rust resistance locus Yr57 on chromosome 3BS of wheat. Mol. Breed. 2015, 35, 65. [Google Scholar] [CrossRef]
- Chhetri, M.; Bariana, H.S.; Kandiah, P.; Bansal, U.K. Yr58: A new stripe rust resistance gene and its interaction with Yr46 for enhanced resistance. Phytopathology 2016, 106, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Maree, G.J.; Prins, R.; Boyd, L.A.; Castelyn, H.D.; Bender, C.M.; Boshoff, W.H.P.; Pretorius, Z.A. Assessing the individual and combined effects of QTLfor adult plant stripe rust resistance derived from Cappelle-Desprez. Agronomy 2019, 9, 154. [Google Scholar] [CrossRef]
- Bariana, H.S.; Miah, H.; Brown, G.N.; Willey, N.; Lehmensiek, A. Molecular mapping of durable rust resistance in wheat and its implication in breeding. In Wheat production in stressed environments: Proceedings of the 7th International Wheat Conference, Mar del Plata, Argentina, 27 November–2 December 2005; Buck, H.T., Nisi, J.E., Salomón, N., Eds.; Springer: Netherlands, Dordrecht, 2 December 2007; pp. 723–728. [Google Scholar]
- Randhawa, M.; Bansal, U.; Valarik, M.; Klocova, B.; Dolezel, J.; Bariana, H. Molecular mapping of stripe rust resistance gene Yr51 in chromosome 4AL of wheat. Theor. Appl. Genet. 2014, 127, 317–324. [Google Scholar] [CrossRef]
- McIntosh, R.A.; Park, R.F.; Wellings, C.R. Wheat Rusts—An Atlas of Resistance Genes; CSIRO Publishing: Collingwood, Australia, 1995; pp. 9–12. [Google Scholar]
- Manly, K.F.; Cudmore, R.H., Jr.; Meer, J.M. Map Manager QTX, cross-platform software for genetic mapping. Mammal. Genom. 2001, 12, 930–932. [Google Scholar] [CrossRef]
- Lagudah, E.S.; McFadden, H.; Singh, R.P.; Huerta-Espino, J.; Bariana, H.S.; Spielmeyer, W. Molecular genetic characterization of the Lr34/Yr18 slow rusting resistance gene region in wheat. Theor. Appl. Genet. 2006, 114, 21–30. [Google Scholar] [CrossRef]
- Lagudah, E.S.; Krattinger, S.G.; Herrera-Foessel, S.; Singh, R.P.; Huerta-Espino, J.; Spielmeyer, W.; Brown-Guedira, G.; Selter, L.L.; Keller, B. Gene-specific markers for the wheat gene Lr34/Yr18/Pm38 which confers resistance to multiple fungal pathogens. Theor. Appl. Genet. 2009, 119, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Wright, S. Volume 1. Genetic and Biometric Foundations. In Evolution and the Genetics of Populations; University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Cockerham, C.C. Covariances of Relatives from Self-Fertilization 1. Crop Sci. 1983, 23, 1177–1180. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Number of RILs | |||
|---|---|---|---|
| Response Category | Observed | Expected | χ21:1 |
| Homozygous Resistant (HR) | 58 | 58.5 | 0.004 |
| Homozygous Susceptible (HS) | 59 | 58.5 | 0.004 |
| Total | 117 | 117 | 0.008 * |
| QTL | Season/Site | Peak Marker | Flanking Markers | LOD | R2 |
|---|---|---|---|---|---|
| QYr.sun-1BL | 2014 LDN | 4406454 | 12766133 & 1074469 | 10.45 | 26 |
| 2015 K | 4406454 | 12766133 &1074469 | 6.69 | 13 | |
| 2015 LDN | 4406454 | 12766133 &1074469 | 6.18 | 15 | |
| Mean | 18 | ||||
| QYr.sun-2AS | 2014 LDN | 4539050 | 1095379 & 4440391 | 4.54 | 11.4 |
| 2015 K | 4539050 | 1095379 & 4440391 | 9.54 | 22 | |
| 2015 LDN | 4539050 | 1095379 & 4440391 | 4.54 | 13 | |
| Mean | 15.3 | ||||
| QYr.sun-3BS | 2014 LDN | YrSen | 4409093 & 1012045 | 2.73 | 6 |
| 2015 K | YrSen | 4409093 & 1012045 | 6.17 | 13.6 | |
| 2015 LDN | YrSen | 4409093 &1012045 | 4.47 | 12.1 | |
| Mean | 10.6 |
| Severity (%) | Severity Reduction (%) | |||||
|---|---|---|---|---|---|---|
| QTL | Experiment | Chromosome | Marker | Sentinel Allele | Nyb3 Allele | |
| QYr.sun-1BL | 2014 LDN | 1BL | 4406454 | 34.6 | 77.2 | 42.6 |
| 2015 K | 1BL | 4406454 | 56.1 | 79.4 | 23.4 | |
| 2015 LDN | 1BL | 4406454 | 45.0 | 75.3 | 30.3 | |
| QYr.sun-2AS | 2014 LDN | 2AS | 4539050 | 44.1 | 77.2 | 33.1 |
| 2015 K | 2AS | 4539050 | 48.0 | 79.4 | 31.4 | |
| 2015 LDN | 2AS | 4539050 | 47.9 | 75.3 | 27.4 | |
| QYr.sun-3BS | 2014 LDN | 3BS | YrSen | 26.7 | 77.2 | 50.6 |
| 2015 K | 3BS | YrSen | 49.2 | 79.4 | 30.3 | |
| 2015 LDN | 3BS | YrSen | 50.0 | 75.3 | 25.3 | |
| Loci | Rust Severity (%) | ||
|---|---|---|---|
| 2014 | 2015 | ||
| LDN | K | LDN | |
| QYr.sun-1BL | 34.58 | 56.07 | 45.00 |
| QYr.sun-2AS | 44.11 | 48.04 | 47.91 |
| QYr.sun-3BS | 26.67 | 49.17 | 50.00 |
| QYr.sun-1BL + QYr.sun-2AS | 11.67 | 34.50 | 28.75 |
| QYr.sun-1BL + QYr.sun-3BS | 17.06 | 34.38 | 30.74 |
| QYr.sun-2AS + QYr.sun-3BS | 28.50 | 28.75 | 26.25 |
| QYr.sun-1BL + QYr.sun-2AS + QYr.sun-3BS | 6.72 | 12.94 | 13.53 |
| Critical Difference (CD) | 9.68 ± 4.98 | 10.77 ± 5.54 | 9.88 ± 5.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chemayek, B.; Wagoire, W.; Bansal, U.; Bariana, H. A Combination of Three Genomic Regions Conditions High Level of Adult Plant Stripe Rust Resistance in Australian Wheat Cultivar Sentinel. Plants 2024, 13, 129. https://doi.org/10.3390/plants13010129
Chemayek B, Wagoire W, Bansal U, Bariana H. A Combination of Three Genomic Regions Conditions High Level of Adult Plant Stripe Rust Resistance in Australian Wheat Cultivar Sentinel. Plants. 2024; 13(1):129. https://doi.org/10.3390/plants13010129
Chicago/Turabian StyleChemayek, Bosco, William Wagoire, Urmil Bansal, and Harbans Bariana. 2024. "A Combination of Three Genomic Regions Conditions High Level of Adult Plant Stripe Rust Resistance in Australian Wheat Cultivar Sentinel" Plants 13, no. 1: 129. https://doi.org/10.3390/plants13010129
APA StyleChemayek, B., Wagoire, W., Bansal, U., & Bariana, H. (2024). A Combination of Three Genomic Regions Conditions High Level of Adult Plant Stripe Rust Resistance in Australian Wheat Cultivar Sentinel. Plants, 13(1), 129. https://doi.org/10.3390/plants13010129