
Phenotype, Biomass, Carbon and Nitrogen Assimilation, and Antioxidant Response of Rapeseed under Salt Stress

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effects of Salt Stress on Phenotype
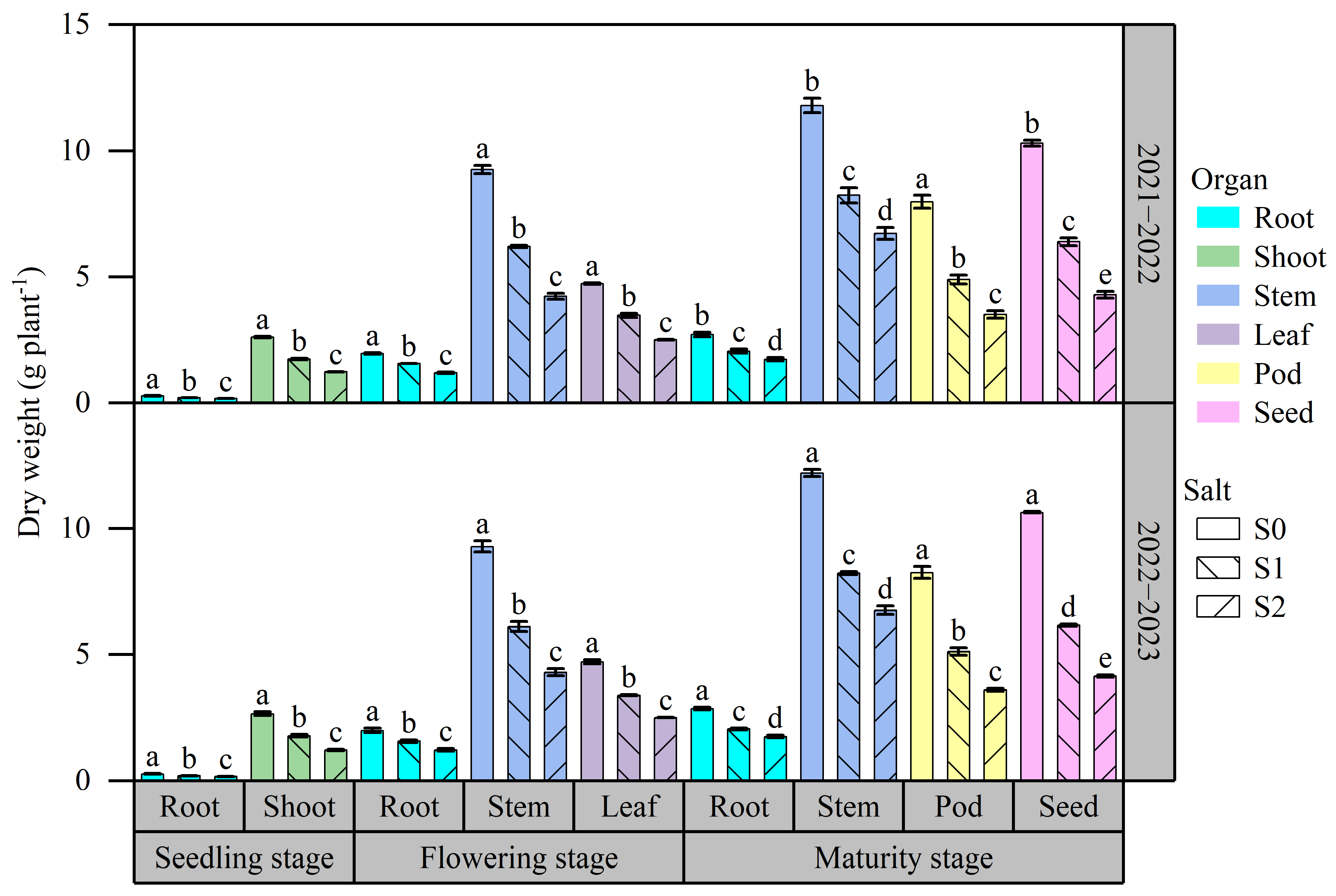
2.2. Effects of Salt Stress on Biomass Accumulation and Distribution
2.3. Effects of Salt Stress on C and N Assimilation
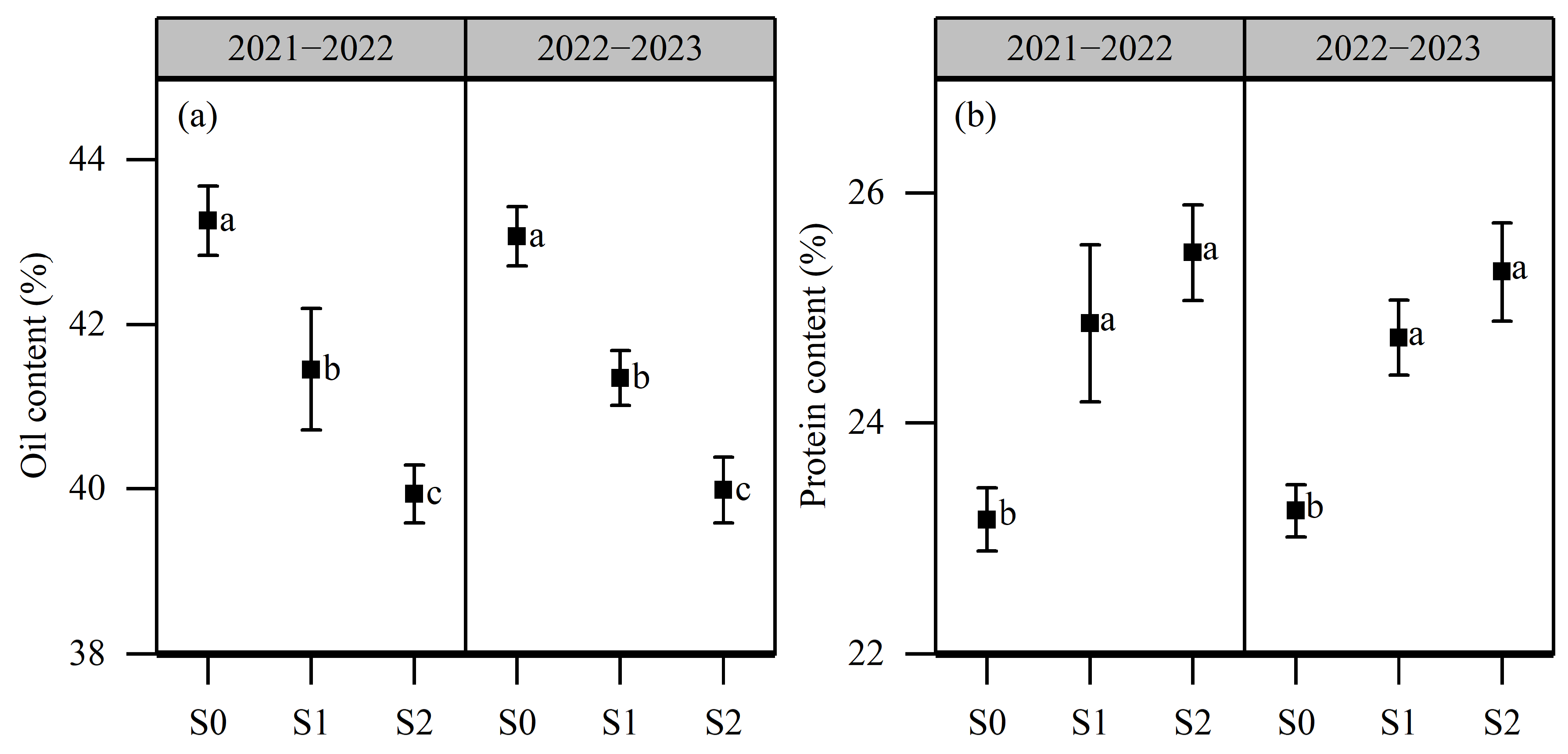
2.4. Effects of Salt Stress on Oil Content and Protein Content
2.5. Effects of Salt Stress on Antioxidants and Osmolytes
3. Discussion
3.1. Response of Phenotypic Traits and Biomass in Rapeseed to Salt Stress
3.2. Relationship between C and N Assimilation and Seed Quality in Rapeseed under Salt Stress
3.3. Response of Antioxidants and Osmolytes in Rapeseed to Salt Stress
4. Materials and Methods
4.1. Experiment Design
4.2. Sample and Measurement
4.2.1. Phenotypic Traits and Biomass
4.2.2. Content and Accumulation of C and N
4.2.3. Antioxidants and Osmolytes
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Khan, Z.; Khan, M.N.; Zhang, K.; Luo, T.; Zhu, K.; Hu, L. The application of biochar alleviated the adverse effects of drought on the growth, physiology, yield and quality of rapeseed through regulation of soil status and nutrients availability. Ind. Crop. Prod. 2021, 171, 113878. [Google Scholar] [CrossRef]
- Koutsouki, A.A.; Tegou, E.; Badeka, A.; Kontakos, S.; Pomonis, P.J.; Kontominas, M.G. In situ and conventional transesterification of rapeseeds for biodiesel production: The effect of direct sonication. Ind. Crop. Prod. 2016, 84, 399–407. [Google Scholar] [CrossRef]
- Qian, J.; Yang, Q.; Sun, F.; He, M.; Chen, Q.; Yun, Z.; Qin, L. Cogeneration of biodiesel and nontoxic rapeseed meal from rapeseed through in-situ alkaline transesterification. Bioresour. Technol. 2013, 128, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of plant responses to salt stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef] [PubMed]
- Machado, R.M.A.; Serralheiro, R.P. Soil Salinity: Effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Jamil, A.; Riaz, S.; Ashraf, M.; Foolad, M.R. Gene expression profiling of plants under salt stress. Crit. Rev. Plant Sci. 2011, 30, 435–458. [Google Scholar] [CrossRef]
- Shahzad, B.; Rehman, A.; Tanveer, M.; Wang, L.; Park, S.K.; Ali, A. Salt stress in Brassica: Effects, tolerance mechanisms, and management. J. Plant Growth Regul. 2022, 41, 781–795. [Google Scholar] [CrossRef]
- Wu, H. Plant salt tolerance and Na+ sensing and transport. Crop J. 2018, 6, 215–225. [Google Scholar] [CrossRef]
- Dolatabadi, N.; Toorchi, M.; Valizadeh, M.; Bandehagh, A. The proteome response of salt-sensitive rapeseed (Brassica napus L.) genotype to salt stress. Not. Bot. Horti Agrobot. 2019, 47, 17–23. [Google Scholar] [CrossRef]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Chloride in soils and its uptake and movement within the plant: A review. Ann. Bot. 2001, 88, 967–988. [Google Scholar] [CrossRef]
- Mittal, S.; Kumari, N.; Sharma, V. Differential response of salt stress on Brassica Juncea: Photosynthetic performance, pigment, proline, d1 and antioxidant enzymes. Plant Physiol. Biochem. 2012, 54, 17–26. [Google Scholar] [CrossRef]
- Jia, K.; Yan, C.; Yan, H.; Gao, J. Physiological responses of turnip (Brassica rapa L. Subsp. Rapa) seedlings to salt stress. Hortscience 2020, 55, 1567–1574. [Google Scholar] [CrossRef]
- Verma, K.; Dixit, S.; Shekhawat, G.S.; Alam, A. Antioxidant activity of heme oxygenase 1 in Brassica juncea (L.) czern. (Indian Mustard) under salt stress. Turk. J. Biol. 2015, 39, 540–549. [Google Scholar] [CrossRef]
- Dietz, K.-J.; Mittler, R.; Noctor, G. Recent progress in understanding the role of reactive oxygen species in plant cell signaling. Plant Physiol. 2016, 171, 1535–1539. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.-M.; Zheng, D.-F.; Feng, N.-J.; Zhou, G.-S.; Khan, A.; Lu, X.-T.; Deng, P.; Zhou, H.; Du, Y.-W. Regulatory effects of hemin on prevention and rescue of salt stress in rapeseed (Brassica napus L.) seedlings. BMC Plant Biol. 2023, 23, 558. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Cai, J.; Li, J.; Lu, G.; Li, C.; Fu, G.; Zhang, X.; Ma, H.; Liu, Q.; Zou, X.; et al. Exogenous application of a low concentration of melatonin enhances salt tolerance in rapeseed (Brassica napus L.) seedlings. J. Integr. Agric. 2018, 17, 328–335. [Google Scholar] [CrossRef]
- Chen, L.; Liu, L.; Lu, B.; Ma, T.; Jiang, D.; Li, J.; Zhang, K.; Sun, H.; Zhang, Y.; Bai, Z.; et al. Exogenous melatonin promotes seed germination and osmotic regulation under salt stress in cotton (Gossypium hirsutum L.). PLoS ONE 2020, 15, e0228241. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Zhou, H. Plant salt response: Perception, signaling, and tolerance. Front. Plant Sci. 2023, 13, 1053699. [Google Scholar] [CrossRef]
- Lawlor, D.W. Carbon and nitrogen assimilation in relation to yield: Mechanisms are the key to understanding production systems. J. Exp. Bot. 2002, 53, 773–787. [Google Scholar] [CrossRef]
- Carmo-Silva, E.; Scales, J.C.; Madgwick, P.J.; Parry, M.A.J. Optimizing rubisco and its regulation for greater resource use efficiency. Plant Cell Environ. 2015, 38, 1817–1832. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hu, B.; Chu, C. Nitrogen assimilation in plants: Current status and future prospects. J. Genet. Genom. 2022, 49, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhou, C.; Burnap, R.L.; Peng, L. Carbon/nitrogen metabolic balance: Lessons from cyanobacteria. Trends Plant Sci. 2018, 23, 1116–1130. [Google Scholar] [CrossRef] [PubMed]
- Busch, F.A.; Sage, R.F.; Farquhar, G.D. Plants increase CO2 uptake by assimilating nitrogen via the photorespiratory pathway. Nat. Plants 2018, 4, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Mostafa, S.; Lu, Z.; Jin, B. Melatonin-mediated abiotic stress tolerance in plants. Front. Plant Sci. 2022, 13, 847175. [Google Scholar] [CrossRef] [PubMed]
- Bybordi, A. The influence of salt stress on seed germination, growth and yield of canola cultivars. Not. Bot. Horti Agrobot. 2010, 38, 128–133. [Google Scholar]
- Pitann, B.; Schubert, S.; Muehling, K.H. Decline in leaf growth under salt stress is due to an inhibition of H+-pumping activity and increase in apoplastic pH of maize leaves. J. Plant Nutr. Soil Sci. 2009, 172, 535–543. [Google Scholar] [CrossRef]
- Wang, L.; Zheng, J.; Zhou, G.; Li, J.; Qian, C.; Lin, G.; Li, Y.; Zuo, Q. Moderate nitrogen application improved salt tolerance by enhancing photosynthesis, antioxidants, and osmotic adjustment in rapeseed (Brassica napus L.). Front. Plant Sci. 2023, 14, 1196319. [Google Scholar] [CrossRef]
- Wang, L.; Zuo, Q.; Zheng, J.; You, J.; Yang, G.; Leng, S. Salt stress decreases seed yield and postpones growth process of canola (Brassica napus L.) by changing nitrogen and carbon characters. Sci. Rep. 2022, 12, 17884. [Google Scholar] [CrossRef]
- Bakirov, A.; Zhang, Y.; Zhang, Q.; Seitahmetovna, S.A.; Yu, X.; Shi, Y.; Xu, Y.; Wang, K.; Qin, M.; Xu, A.; et al. Screening of salt tolerance traits and the salt tolerance evaluation method in Brassica napus at the seed germination stage. Ital. J. Agron. 2022, 17, 2011. [Google Scholar] [CrossRef]
- Hooks, T.; Niu, G.; Ganjegunte, G. Seedling Emergence and Seedling growth of mustard and rapeseed genotypes under salt stress. Agrosyst. Geosci. Environ. 2019, 2, 1–8. [Google Scholar] [CrossRef]
- Bandeh-Hagh, A.; Toorchi, M.; Mohammadi, A.; Chaparzadeh, N.; Salekdeh, G.H.; Kazemnia, H. Growth and osmotic adjustment of canola genotypes in response to salinity. J. Food Agric. Environ. 2008, 6, 201–208. [Google Scholar]
- Ahmadi, F.I.; Karimi, K.; Struik, P.C. Effect of exogenous application ofmethyl jasmonate on physiological and biochemical characteristics of Brassica napus L. Cv. Talaye under salinity stress. S. Afr. J. Bot. 2018, 115, 5–11. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Sha, H.; Liu, H.; Hu, B.; Gu, J.; Hu, W.; Jia, Y.; Wang, X.; Chang, H.; Zhao, H. Effect of salicylic acid on the dry matter and nitrogen accumulation, partitioning and translocation in two contrasting rice genotypes under salt stress. Pak. J. Bot. 2019, 51, 1541–1550. [Google Scholar] [CrossRef]
- Perez-Lopez, U.; Robredo, A.; Miranda-Apodaca, J.; Lacuesta, M.; Munoz-Rueda, A.; Mena-Petite, A. Carbon dioxide enrichment moderates salinity-induced effects on nitrogen acquisition and assimilation and their impact on growth in barley plants. Environ. Exp. Bot. 2013, 87, 148–158. [Google Scholar] [CrossRef]
- Zuo, Q.; Liu, J.; Shan, J.; Zhou, J.; Wang, L.; Yang, G.; Leng, S.; Liu, H. Carbon and nitrogen assimilation and partitioning in canola (Brassica napus L.) in saline environment. Commun. Soil Sci. Plant Anal. 2019, 50, 1700–1709. [Google Scholar] [CrossRef]
- Guo, L.; Lu, Y.; Bao, S.; Zhang, Q.; Geng, Y.; Shao, X. Carbon and nitrogen metabolism in rice cultivars affected by salt-alkaline stress. Crop Pasture Sci. 2021, 72, 372–382. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Torabian, S. Biochar improved nodulation and nitrogen metabolism of soybean under salt stress. Symbiosis 2018, 74, 215–223. [Google Scholar] [CrossRef]
- Camejo, D.; Guzman-Cedeno, A.; Moreno, A. Reactive oxygen species, essential molecules, during plant-pathogen interactions. Plant Physiol. Biochem. 2016, 103, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.R.; Asgher, M.; Khan, N.A. Alleviation of salt-induced photosynthesis and growth inhibition by salicylic acid involves glycinebetaine and ethylene in mungbean (Vigna radiata L.). Plant Physiol. Biochem. 2014, 80, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Apodaca, J.; Agirresarobe, A.; Martinez-Goni, X.S.; Yoldi-Achalandabaso, A.; Perez-Lopez, U. N metabolism performance in Chenopodium quinoa subjected to drought or salt stress conditions. Plant Physiol. Biochem. 2020, 155, 725–734. [Google Scholar] [CrossRef]
- Hurtado, A.C.; Chiconato, D.A.; Prado, R.d.M.; Sousa, G.d.S., Jr.; Viciedo, D.O.; Piccolo, M.d.C. Silicon application induces changes c:n:p stoichiometry and enhances stoichiometric homeostasis of sorghum and sunflower plants under salt stress. Saudi J. Biol. Sci. 2020, 27, 3711–3719. [Google Scholar] [CrossRef]
- Gupta, M.; Bhaskar, P.B.; Sriram, S.; Wang, P. Integration of omics approaches to understand oil/protein content during seed development in oilseed crops. Plant Cell Rep. 2017, 36, 637–652. [Google Scholar] [CrossRef]
- Zhu, J.; Dai, W.; Chen, B.; Cai, G.; Wu, X.; Yan, G. Research Progress on the Effect of Nitrogen on Rapeseed between Seed Yield and Oil Content and Its Regulation Mechanism. Int. J. Mol. Sci. 2023, 24, 14504. [Google Scholar] [CrossRef] [PubMed]
- Steppuhn, H.; Falk, K.C.; Zhou, R. Emergence, height, grain yield and oil content of camelina and canola grown in saline media. Can. J. Soil Sci. 2010, 90, 151–164. [Google Scholar] [CrossRef]
- Tian, T.; Wang, J.; Wang, H.; Cui, J.; Shi, X.; Song, J.; Li, W.; Zhong, M.; Qiu, Y.; Xu, T. Nitrogen application alleviates salt stress by enhancing osmotic balance, ROS scavenging, and photosynthesis of rapeseed seedlings (Brassica napus). Plant Signal. Behav. 2022, 17, e2081419. [Google Scholar] [CrossRef]
- Yildiz, M.; Akcali, N.; Terzi, H. Proteomic and biochemical responses of canola (Brassica napus L.) exposed to salinity stress and exogenous lipoic acid. J. Plant Physiol. 2015, 179, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Kumar, R.; Jain, V.; Jain, S. Differential behavior of the antioxidant system in response to salinity induced oxidative stress in salt-tolerant and salt-sensitive cultivars of Brassica juncea L. Biocatal. Agric. Biotechnol. 2018, 13, 12–19. [Google Scholar] [CrossRef]
- El-Badri, A.M.; Batool, M.; Mohamed, I.A.A.; Wang, Z.; Khatab, A.; Sherif, A.; Ahmad, H.; Khan, M.N.; Hassan, H.M.; Elrewainy, I.M.; et al. Antioxidative and metabolic contribution to salinity stress responses in two rapeseed cultivars during the early seedling stage. Antioxidants 2021, 10, 1227. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Year | Soil Salinity | Seedling Stage | Flowering Stage | Maturity Stage | |||||
|---|---|---|---|---|---|---|---|---|---|
| Plant Height (cm) | Root Neck Diameter (mm) | Leaf Area (cm2 Plant−1) | Plant Height (cm) | Root Neck Diameter (mm) | Leaf Area (cm2 Plant−1) | Plant Height (cm) | Root Neck Diameter (mm) | ||
| 2021–2022 | S0 | 15.14 a | 2.72 a | 174.0 b | 82.46 a | 14.63 a | 967 a | 121.90 b | 14.95 a |
| S1 | 13.70 b | 2.60 bc | 118.6 c | 74.08 b | 14.00 b | 713 b | 111.49 c | 14.33 b | |
| S2 | 12.27 c | 2.50 d | 82.5 d | 67.15 c | 13.50 c | 525 c | 104.28 d | 13.53 c | |
| 2022–2023 | S0 | 15.59 a | 2.76 a | 177.9 a | 84.09 a | 14.46 a | 978 a | 126.44 a | 15.01 a |
| S1 | 13.91 b | 2.61 b | 121.4 c | 73.61 b | 13.95 b | 718 b | 113.08 c | 14.23 b | |
| S2 | 12.42 c | 2.53 cd | 83.2 d | 68.13 c | 13.52 c | 536 c | 102.22 d | 13.82 bc | |
| ANOVA | |||||||||
| Year | * | ns | * | ns | ns | ns | ns | ns | |
| Soil salinity | ** | ** | ** | ** | ** | ** | ** | ** | |
| Year × soil salinity | ns | ns | ns | ns | ns | ns | * | ns | |
| Year | Soil Salinity | Seedling Stage | Flowering Stage | Maturity Stage | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Root | Shoot | Root | Stem | Leaf | Root | Stem | Pod | Seed | ||
| 2021–2022 | S0 | 9.45 c | 90.55 a | 12.30 c | 58.05 a | 29.65 bc | 8.27 c | 35.97 c | 24.31 a | 31.45 a |
| S1 | 10.07 b | 89.93 b | 13.94 b | 55.17 b | 30.89 a | 9.53 b | 38.15 b | 22.69 b | 29.63 b | |
| S2 | 12.15 a | 87.85 c | 15.11 a | 53.28 c | 31.61 a | 10.67 a | 41.32 a | 21.57 c | 26.44 c | |
| 2022–2023 | S0 | 9.44 c | 90.56 a | 12.49 c | 58.05 a | 29.46 c | 8.43 c | 35.91 c | 24.32 a | 31.34 a |
| S1 | 10.08 b | 89.92 b | 14.17 b | 55.21 b | 30.62 ab | 9.54 b | 38.19 b | 23.71 a | 28.56 b | |
| S2 | 12.22 a | 87.78 c | 15.27 a | 53.49 c | 31.24 a | 10.8 a | 41.51 a | 22.15 bc | 25.53 c | |
| ANOVA | ||||||||||
| Year | ns | ns | ns | ns | ns | ns | ns | * | ns | |
| Soil salinity | ** | ** | ** | ** | ** | ** | ** | ** | ** | |
| Year × salt | ns | ns | ns | ns | ns | ns | ns | ns | ** | |
| Year | Soil Salinity | Flowering Stage | Maturity Stage | |||||
|---|---|---|---|---|---|---|---|---|
| Root | Stem | Leaf | Root | Stem | Pod | Seed | ||
| 2021–2022 | S0 | 0.81 a | 3.68 a | 1.95 a | 1.11 b | 4.90 a | 3.26 a | 6.00 b |
| S1 | 0.64 b | 2.39 b | 1.40 b | 0.83 c | 3.34 b | 1.98 b | 3.64 c | |
| S2 | 0.48 c | 1.60 c | 0.99 c | 0.70 d | 2.70 c | 1.41 c | 2.42 e | |
| 2022–2023 | S0 | 0.82 a | 3.68 a | 1.94 a | 1.17 a | 5.07 a | 3.38 a | 6.20 a |
| S1 | 0.64 b | 2.35 b | 1.36 b | 0.83 c | 3.35 b | 2.07 b | 3.51 d | |
| S2 | 0.49 c | 1.62 c | 0.99 c | 0.71 d | 2.72 c | 1.44 c | 2.34 e | |
| ANOVA | ||||||||
| Year | ns | ns | ns | ns | ns | ns | ns | |
| Soil salinity | ** | ** | ** | ** | ** | ** | ** | |
| Year × soil salinity | ns | ns | ns | ns | ns | ns | ** | |
| Year | Soil Salinity | Flowering Stage | Maturity Stage | |||||
|---|---|---|---|---|---|---|---|---|
| Root | Stem | Leaf | Root | Stem | Pod | Seed | ||
| 2021–2022 | S0 | 23.19 a | 152.68 a | 184.46 a | 17.13 a | 73.71 ab | 56.41 a | 365.07 b |
| S1 | 20.52 b | 111.44 b | 142.34 b | 15.49 b | 68.01 cd | 39.07 b | 241.46 c | |
| S2 | 16.40 c | 81.02 c | 105.10 c | 14.54 b | 62.97 d | 30.90 c | 165.42 e | |
| 2022–2023 | S0 | 23.61 a | 153.88 a | 183.73 a | 17.88 a | 75.79 a | 58.89 a | 380.33 a |
| S1 | 20.72 b | 110.66 b | 138.62 b | 15.51 b | 68.54 bc | 41.01 b | 234.17 d | |
| S2 | 16.60 c | 82.39 c | 104.85 c | 14.56 b | 63.5 cd | 31.96 c | 162.97 e | |
| ANOVA | ||||||||
| Year | ns | ns | ns | ns | ns | ns | ns | |
| Soil salinity | ** | ** | ** | ** | ** | ** | ** | |
| Year × salt | ns | ns | ns | ns | ns | ns | ** | |
| Year | Soil Salinity | SOD (U g−1 FW) | CAT (U g−1 FW) | Soluble Protein (mg g−1 FW) | Soluble Sugar (mg g−1 FW) |
|---|---|---|---|---|---|
| 2021–2022 | S0 | 467 c | 126 c | 7.63 c | 104 c |
| S1 | 625 b | 154 b | 8.19 b | 114 b | |
| S2 | 735 a | 170 a | 8.77 a | 119 a | |
| 2022–2023 | S0 | 466 c | 126 c | 7.55 c | 103 c |
| S1 | 621 b | 152 b | 8.22 b | 113 b | |
| S2 | 742 a | 167 a | 8.77 a | 118 a | |
| ANOVA | |||||
| Year | ns | ns | ns | ns | |
| Soil salinity | ** | ** | ** | ** | |
| Year × salt | ns | ns | ns | ns | |
| Year | Soil Salinity | SOD (U g−1 FW) | CAT (U g−1 FW) | Soluble Protein (mg g−1 FW) | Soluble Sugar (mg g−1 FW) |
|---|---|---|---|---|---|
| 2021–2022 | S0 | 485 c | 154 d | 8.63 c | 157 cd |
| S1 | 591 b | 173 bc | 9.02 b | 159 bc | |
| S2 | 670 a | 187 a | 9.60 a | 165 ab | |
| 2022–2023 | S0 | 479 c | 146 d | 8.57 c | 152 d |
| S1 | 606 b | 166 c | 9.07 b | 159 bcd | |
| S2 | 650 a | 181 ab | 9.67 a | 169 a | |
| ANOVA | |||||
| Year | ns | ** | ns | ns | |
| Soil salinity | ** | ** | ** | ** | |
| Year × salt | * | ns | ns | ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Lin, G.; Li, Y.; Qu, W.; Wang, Y.; Lin, Y.; Huang, Y.; Li, J.; Qian, C.; Yang, G.; et al. Phenotype, Biomass, Carbon and Nitrogen Assimilation, and Antioxidant Response of Rapeseed under Salt Stress. Plants 2024, 13, 1488. https://doi.org/10.3390/plants13111488
Wang L, Lin G, Li Y, Qu W, Wang Y, Lin Y, Huang Y, Li J, Qian C, Yang G, et al. Phenotype, Biomass, Carbon and Nitrogen Assimilation, and Antioxidant Response of Rapeseed under Salt Stress. Plants. 2024; 13(11):1488. https://doi.org/10.3390/plants13111488
Chicago/Turabian StyleWang, Long, Guobing Lin, Yiyang Li, Wenting Qu, Yan Wang, Yaowei Lin, Yihang Huang, Jing Li, Chen Qian, Guang Yang, and et al. 2024. "Phenotype, Biomass, Carbon and Nitrogen Assimilation, and Antioxidant Response of Rapeseed under Salt Stress" Plants 13, no. 11: 1488. https://doi.org/10.3390/plants13111488