

Transcriptomic Profiling of Sugarcane White Leaf (SCWL) Canes during Maturation Phase


Abstract
1. Introduction
2. Results
2.1. Growth Characteristics and Phytoplasma Detection of SCWL Sugarcanes
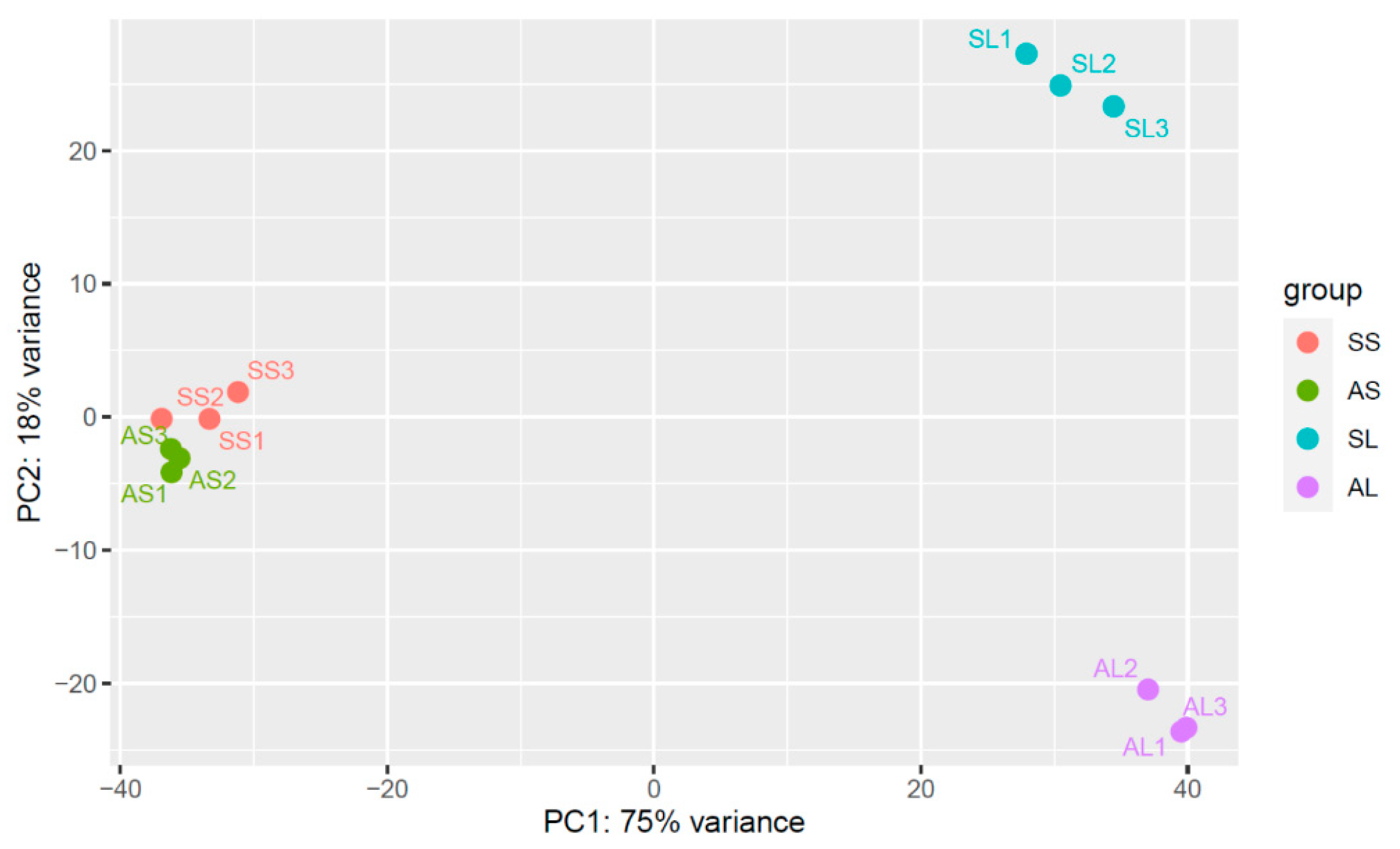
2.2. RNA-Sequencing and Assembly
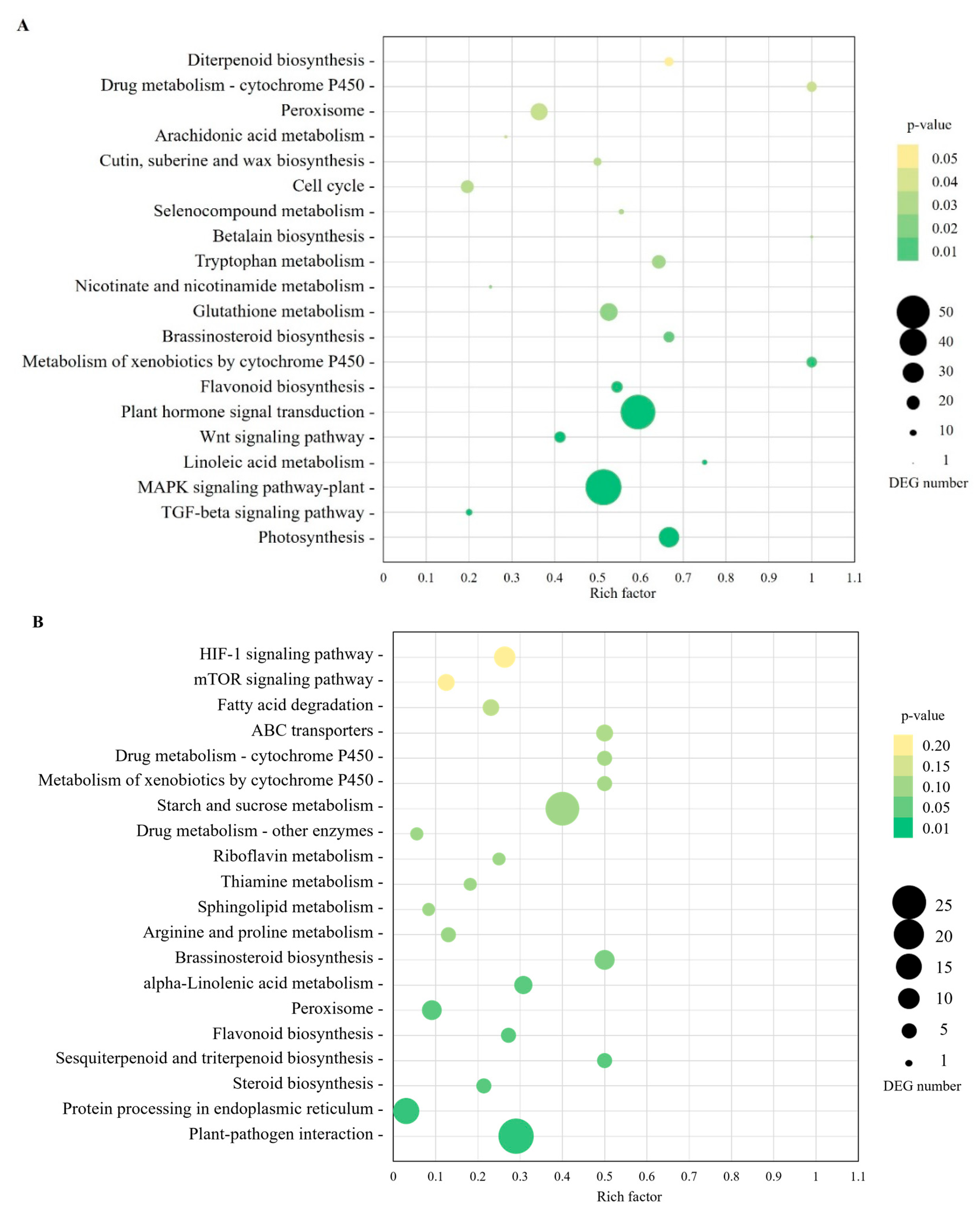
2.3. Gene Ontology (GO) Annotation and KEGG Pathway-Enrichment Analysis
2.4. Phytoplasma-Affected Metabolic Processes of Sugarcane
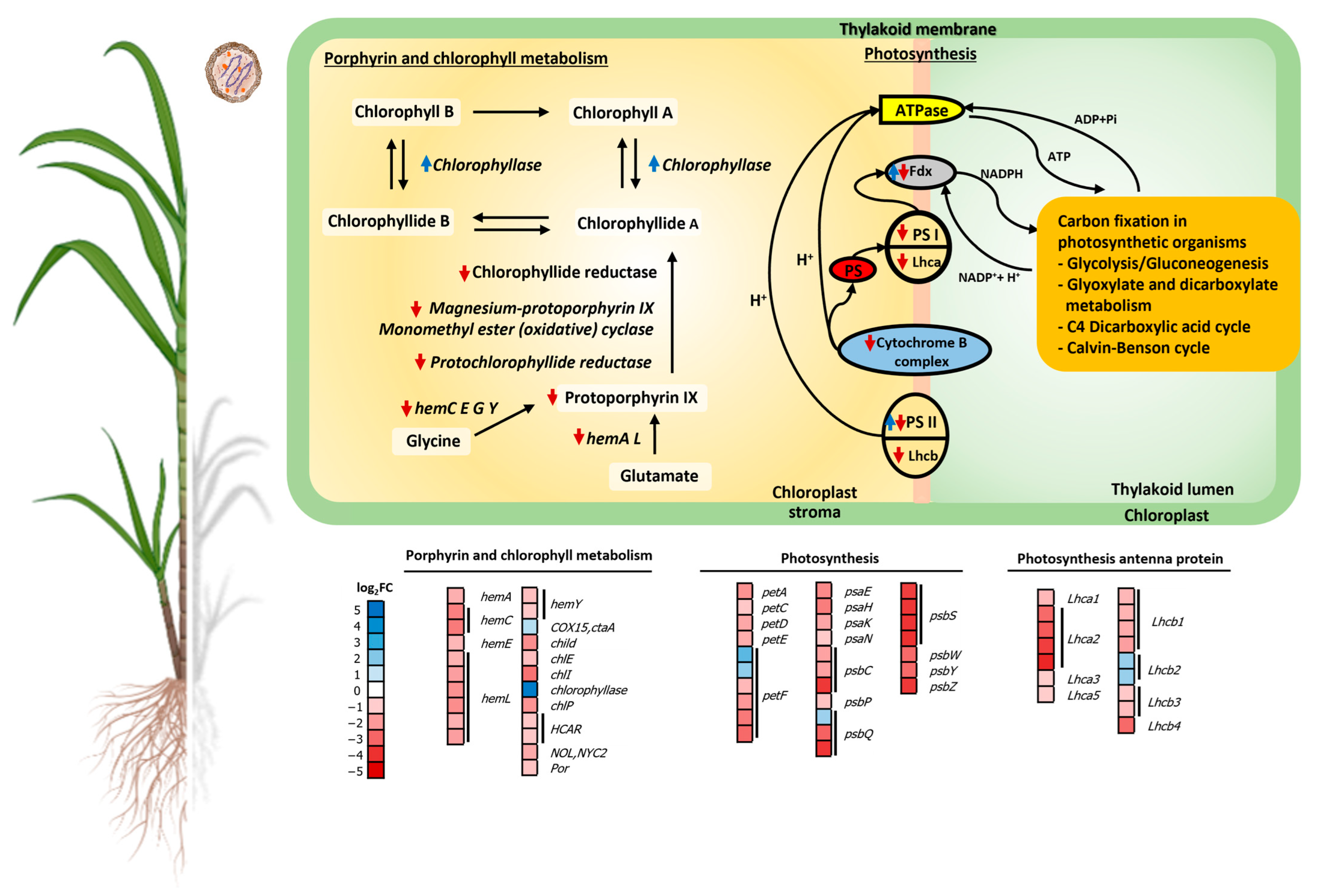
2.4.1. Chlorophyll Metabolism and Photosynthesis
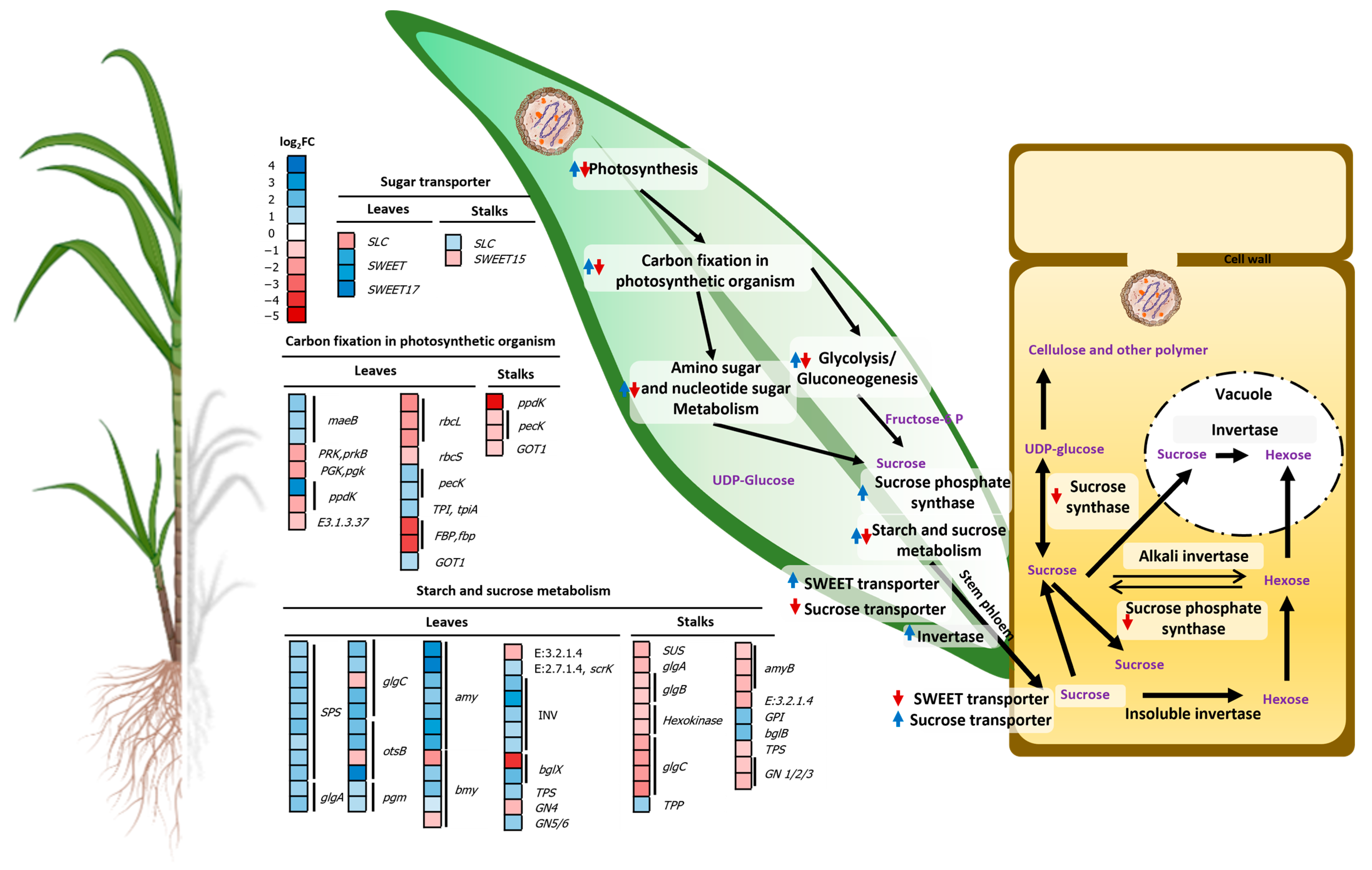
2.4.2. Regulation of Genes Related to Sucrose Accumulation
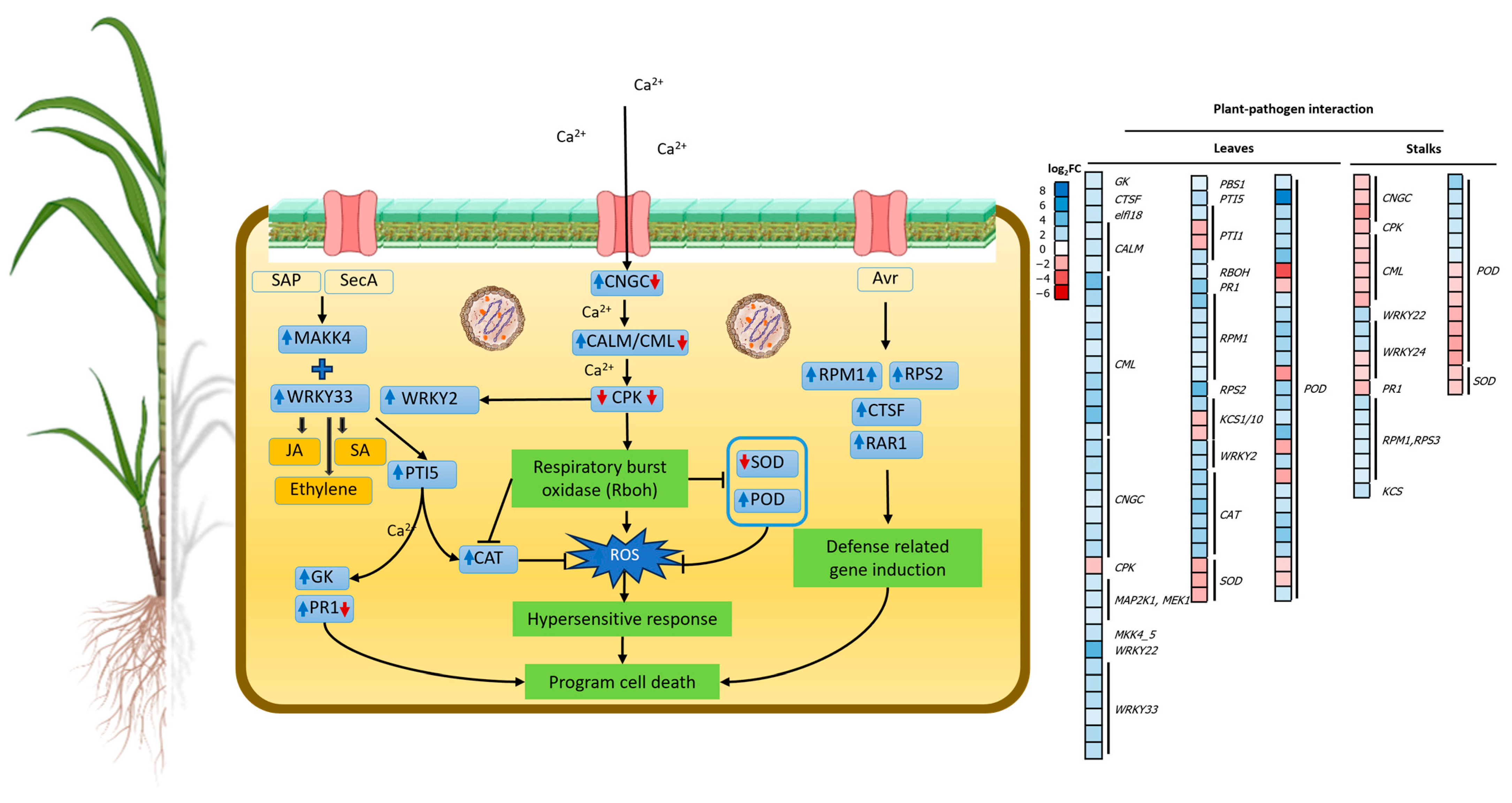
2.4.3. Regulation of Genes Related to Plant–Pathogen Interactions
2.4.4. Regulation of Genes Related to Signal Transduction by Plant Hormones
2.4.5. Regulation of Genes Related to Secondary Metabolites
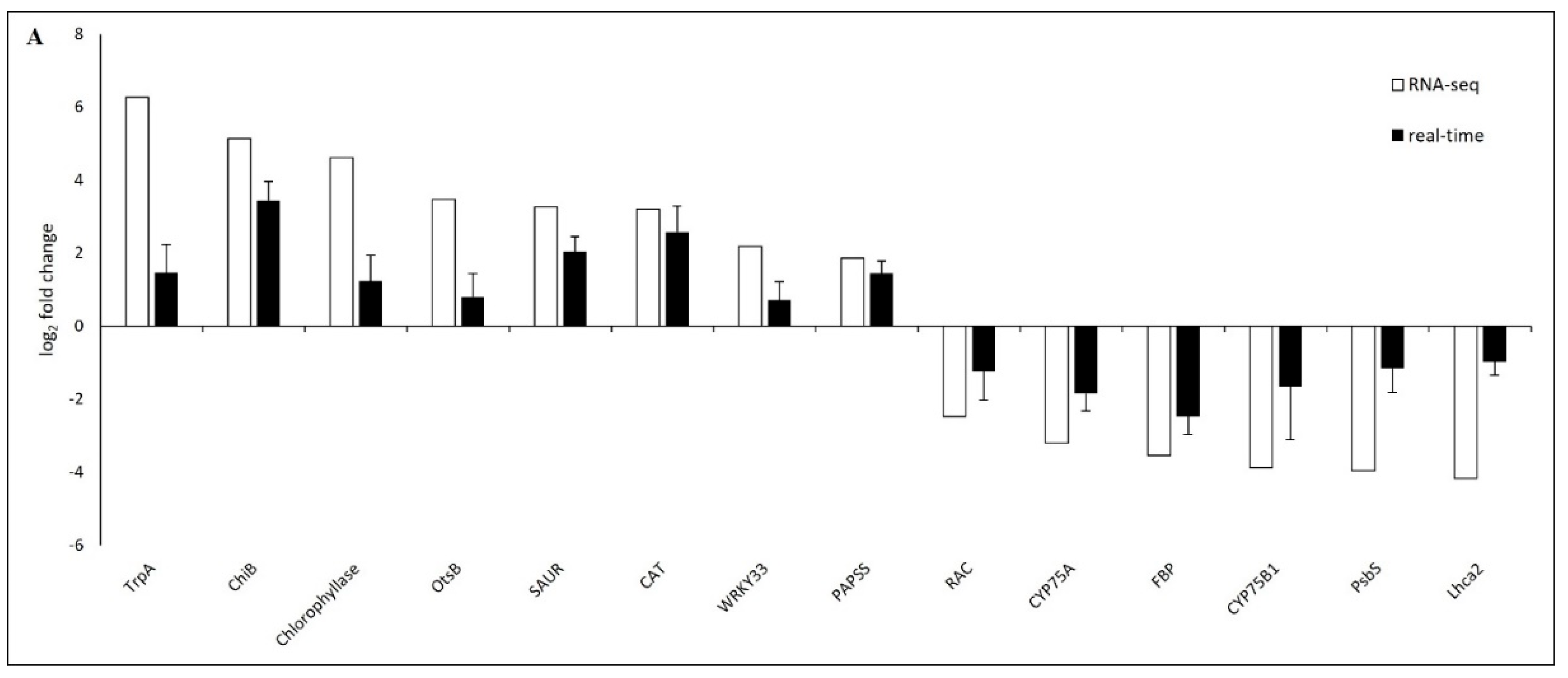
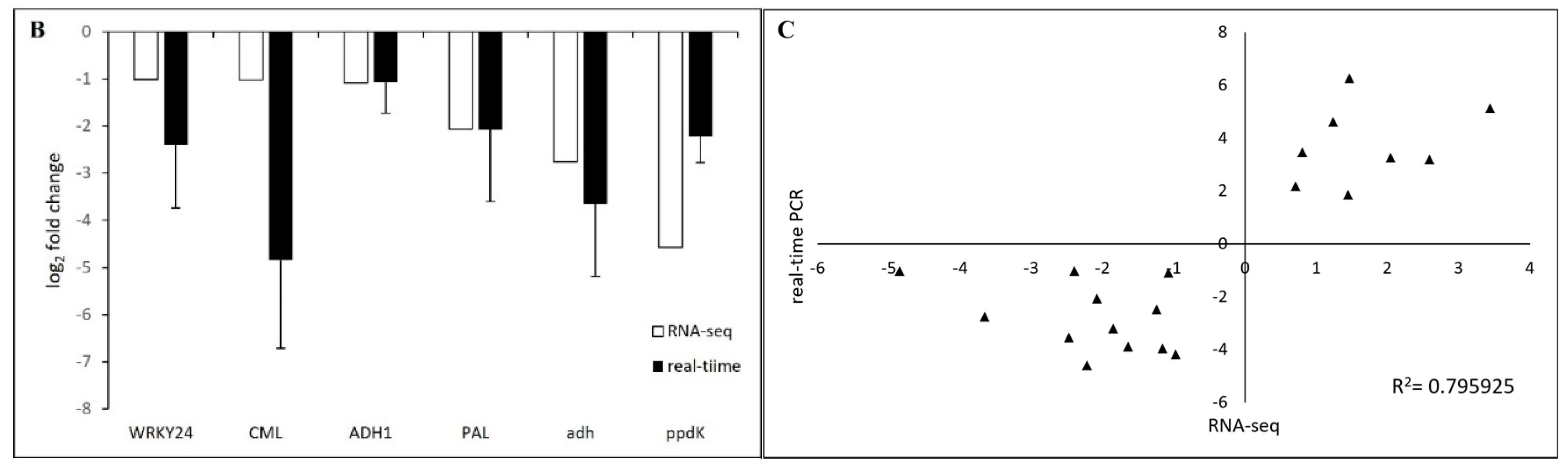
2.5. Verification of DEGs by Real-Time PCR
3. Discussion
3.1. SCWL Phytoplasma Affects Chlorophyll Metabolism and Photosynthesis
3.2. SCWL Phytoplasma Alters the Gene Expression of Sucrose Accumulation
3.3. SCWL Phytoplasma Alters the Expression of Genes Related to Plant–Pathogen Interactions
3.4. SCWL Phytoplasma Accelerates Plant Hormone Signaling Transduction
3.5. SCWL Phytoplasma Alter the Expression of Genes Involved in Secondary Metabolites
4. Conclusions
5. Materials and Methods
5.1. Sugarcane Samples and Plant Growth Parameters
5.2. RNA Extraction
5.3. Detection of Phytoplasma in Sugarcane Using Real-Time PCR
5.4. RNA Sequencing and Transcriptome Analysis
5.5. Quantification of Candidate Genes by Real-Time PCR
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanboonsong, Y.; Wangkeeree, J.; Kobori, Y. Integrated management of the vectors of sugarcane white leaf disease in Thailand: An update. Int. Sugar J. 2017, 119, 220–223. [Google Scholar]
- Wongkaew, P.; Fletcher, J. Sugarcane white leaf phytoplasma in tissue culture: Long-term maintenance, transmission, and oxytetracycline remission. Plant Cell Rep. 2004, 23, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Lee, C.; Teng, W. Studies on sugarcane white leaf disease of Taiwan, with special reference to the transmission by a leafhopper, Epitettix hiroglyphicus Mats. Jpn. J. Phytopathol. 1969, 35, 251–259. [Google Scholar] [CrossRef]
- Hanboonsong, Y.; Ritthison, W.; Choosai, C.; Sirithorn, P. Transmission of sugarcane white leaf phytoplasma by Yamatotettix flavovittatus, a new leafhopper vector. J. Econ. Entomol. 2006, 99, 1531–1537. [Google Scholar] [CrossRef]
- Marcone, C.; Neimark, H.; Ragozzino, A.; Lauer, U.; Seemuller, E. Chromosome sizes of phytoplasmas composing major phylogenetic groups and subgroups. Phytopathology 1999, 89, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Wongkaew, P.; Hanboonsong, Y.; Sirithorn, P.; Choosai, C.; Boonkrong, S.; Tinnangwattana, T.; Kitchareonpanya, R.; Damak, S. Differentiation of phytoplasmas associated with sugarcane and gramineous weed white leaf disease and sugarcane grassy shoot disease by RFLP and sequencing. Theor. Appl. Genet. 1997, 95, 660–663. [Google Scholar] [CrossRef]
- Marcone, C. Phytoplasma diseases of sugarcane. Sugar Tech. 2002, 4, 79–85. [Google Scholar] [CrossRef]
- Nakashima, K.; Murata, N. Destructive plant diseases caused by mycoplasma-like organisms in Asia. Outlook Agric. 1993, 22, 53–58. [Google Scholar] [CrossRef]
- Hagos, H.; Mengistu, L.; Mequanint, Y. Determining optimum harvest age of sugarcane varieties on the newly establishing sugar project in the tropical areas of Tendaho, Ethiopia. Adv. Crop Sci. Technol. 2014, 2, 156. [Google Scholar] [CrossRef]
- Kamwilaisak, K.; Jutakridsada, P.; Iamamornphanth, W.; Saengprachatanarug, K.; Kasemsiri, P.; Konyai, S.; Posom, J.; Chindaprasirt, P. Estimation of sugar content in sugarcane (Saccharum spp.) variety Lumpang 92-11 (LK 92-11) and Khon Kaen 3 (KK 3) by near infrared spectroscopy. Eng. J. 2021, 25, 69–83. [Google Scholar] [CrossRef]
- Garsmeur, O.; Droc, G.; Antonise, R.; Grimwood, J.; Potier, B.; Aitken, K.; Jenkins, J.; Martin, G.; Charron, C.; Hervouet, C.; et al. A mosaic monoploid reference sequence for the highly complex genome of sugarcane. Nat. Commun. 2018, 9, 2638. [Google Scholar] [CrossRef] [PubMed]
- Matile, P.; Hortensteiner, S.; Thomas, H. Chlorophyll degradation. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 67–95. [Google Scholar] [CrossRef] [PubMed]
- Fiedor, L.; Zbyradowski, M.; Pilch, M. Tetrapyrrole pigments of photosynthetic antennae and reaction centers of higher plants: Structures, biophysics, functions, biochemistry, mechanisms of regulation, applications. In Advances in Botanical Research; Grimm, B., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 90, pp. 1–33. [Google Scholar]
- Teixeira, A.; Martins, V.; Frusciante, S.; Cruz, T.; Noronha, H.; Diretto, G.; Geros, H. Flavescence dorée-derived leaf yellowing in grapevine (Vitis vinifera L.) is associated to a general repression of isoprenoid biosynthetic pathways. Front. Plant Sci. 2020, 11, 896. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Jin, X.; Liao, W.; Hu, L.; Dawuda, M.M.; Zhao, X.; Tang, Z.; Gong, T.; Yu, J. 5-Aminolevulinic acid (ALA) alleviated salinity stress in cucumber seedlings by enhancing chlorophyll synthesis pathway. Front. Plant Sci. 2018, 9, 635. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, L.M. Vegetative storage protein, tuberization, senescence, and abscission. In Plant Growth and Development; Srivastava, L.M., Ed.; Academic Press: San Diego, CA, USA, 2002; pp. 473–502. [Google Scholar] [CrossRef]
- Bertamini, M.; Nedunchezhian, N. Effects of phytoplasma [stolbur-subgroup (Bois noir-BN)] on photosynthetic pigments, saccharides, ribulose 1, 5-bisphosphate carboxylase, nitrate and nitrite reductases, and photosynthetic activities in field-grown grapevine (Vitis vinifera L. cv. Chardonnay) leaves. Photosynthetica 2001, 39, 119–122. [Google Scholar] [CrossRef]
- Ji, X.; Gai, Y.; Zheng, C.; Mu, Z. Comparative proteomic analysis provides new insights into mulberry dwarf responses in mulberry (Morus alba L.). Proteomics 2009, 9, 5328–5339. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, E.A.; Farrag, A.A.; Kheder, A.A.; Shaaban, A. Effect of phytoplasma associated with sesame phyllody on ultrastructural modification, physio-biochemical traits, productivity and oil quality. Plants 2022, 11, 477. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhao, J.; Liu, M. Photosynthetic responses to phytoplasma infection in chinese jujube. Plant Physiol. Biochem. 2016, 105, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Liu, Z.; Dai, L.; Bu, J.; Liu, M.; Zhao, Z.; Jiang, Z.; Gao, W.; Zhao, J. Changing host photosynthetic, carbohydrate, and energy metabolisms play important roles in phytoplasma infection. Phytopathology 2018, 108, 1067–1077. [Google Scholar] [CrossRef] [PubMed]
- Hren, M.; Nikolic, P.; Rotter, A.; Blejec, A.; Terrier, N.; Ravnikar, M.; Dermastia, M.; Gruden, K. ‘Bois noir’ phytoplasma induces significant reprogramming of the leaf transcriptome in the field grown grapevine. BMC Genom. 2009, 10, 460. [Google Scholar] [CrossRef] [PubMed]
- Santi, S.; De Marco, F.; Polizzotto, R.; Grisan, S.; Musetti, R. Recovery from stolbur disease in grapevine involves changes in sugar transport and metabolism. Front. Plant Sci. 2013, 4, 171. [Google Scholar] [CrossRef] [PubMed]
- Jagoueix-Eveillard, S.; Tarendeau, F.; Guolter, K.; Danet, J.L.; Bove, J.M.; Garnier, M. Catharanthus roseus genes regulated differentially by mollicute infections. Mol. Plant Microbe Interact. 2001, 14, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Nejat, N.; Cahill, D.M.; Vadamalai, G.; Ziemann, M.; Rookes, J.; Naderali, N. Transcriptomics-based analysis using RNA-Seq of the coconut (Cocos nucifera) leaf in response to yellow decline phytoplasma infection. Mol. Genet. Genom. 2015, 290, 1899–1910. [Google Scholar] [CrossRef] [PubMed]
- Bilgin, D.D.; Zavala, J.A.; Zhu, J.; Clough, S.J.; Ort, D.R.; DeLucia, E.H. Biotic stress globally downregulates photosynthesis genes. Plant Cell Environ. 2010, 33, 1597–1613. [Google Scholar] [CrossRef] [PubMed]
- Bertamini, M.; Grando, M.S.; Nedunchezhian, N. Effects of phytoplasma infection on pigments, chlorophyll-protein complex and photosynthetic activities in field grown apple leaves. Biol. Plant 2003, 46, 237–242. [Google Scholar] [CrossRef]
- Asudi, G.O.; Omenge, K.M.; Paulmann, M.K.; Reichelt, M.; Grabe, V.; Mithofer, A.; Oelmuller, R.; Furch, A.C.U. The physiological and biochemical effects on napier grass plants following napier grass stunt phytoplasma infection. Phytopathology 2021, 111, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Pradit, N.; Rodriguez-Saona, C.; Kawash, J.; Polashock, J. Phytoplasma infection influences gene expression in American cranberry. Front. Ecol. Evol. 2019, 7, 178. [Google Scholar] [CrossRef]
- Ahmad, J.N.; Renaudin, J.; Eveillard, S. Expression of defence genes in stolbur phytoplasma infected tomatoes, and effect of defence stimulators on disease development. Eur. J. Plant Pathol. 2013, 139, 39–51. [Google Scholar] [CrossRef]
- Young, A.; Frank, H. Energy transfer reactions involving carotenoids: Quenching of chlorophyll fluorescence. J. Photochem. Photobiol. B Biol. 1996, 36, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Youssef, S.; Safwat, G.; Baset, A.; Shalaby, A.; El-Beltagi, H. Effect of phytoplasma infection on plant hormones, enzymes and their role in infected sesame. Fresenius Environ. Bull. 2018, 27, 5727–5735. [Google Scholar]
- Polívka, T.; Frank, H.A. Molecular factors controlling photosynthetic light harvesting by carotenoids. Acc. Chem. Res. 2010, 43, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Wang, Z.; Li, X.; Zhao, Z.; Deng, M.; Dong, Y.; Cao, X.; Fan, G. Comparative analysis of Paulownia fortunei response to phytoplasma infection with dimethyl sulfate treatment. Int. J. Genom. 2017, 2017, 6542075. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Wang, H.; Chen, P.; Fu, B.; Zhang, M.; Li, J.; Zheng, X.; Tan, B.; Feng, J. Combination of iTRAQ proteomics and RNA-seq transcriptomics reveals multiple levels of regulation in phytoplasma-infected Ziziphus jujuba Mill. Hortic. Res. 2017, 4, 17080. [Google Scholar] [CrossRef] [PubMed]
- Minges, A.; Groth, G. Small-molecule inhibition of pyruvate phosphate dikinase targeting the nucleotide binding site. PLoS ONE 2017, 12, e0181139. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Fan, G.; Wang, Z.; Gu, Z. Phytoplasma-induced changes in the acetylome and succinylome of Paulownia tomentosa provide evidence for involvement of acetylated proteins in witches’ broom disease. Mol. Cell Proteom. 2019, 18, 1210–1226. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Inaba, J.; Zhao, Y.; Mowery, J.D.; Hammond, R. Phytoplasma infection blocks starch breakdown and triggers chloroplast degradation, leading to premature leaf senescence, sucrose reallocation, and spatiotemporal redistribution of phytohormones. Int. J. Mol. Sci. 2022, 23, 1810. [Google Scholar] [CrossRef] [PubMed]
- Goren, S.; Lugassi, N.; Stein, O.; Yeselson, Y.; Schaffer, A.A.; David-Schwartz, R.; Granot, D. Suppression of sucrose synthase affects auxin signaling and leaf morphology in tomato. PLoS ONE 2017, 12, e0182334. [Google Scholar] [CrossRef] [PubMed]
- Eom, J.S.; Chen, L.Q.; Sosso, D.; Julius, B.T.; Lin, I.W.; Qu, X.Q.; Braun, D.M.; Frommer, W.B. SWEETs, transporters for intracellular and intercellular sugar translocation. Curr. Opin. Plant Biol. 2015, 25, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Maust, B.E.; Espadas, F.; Talavera, C.; Aguilar, M.; Santamaria, J.M.; Oropeza, C. Changes in carbohydrate metabolism in coconut palms infected with the lethal yellowing phytoplasma. Phytopathology 2003, 93, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Lepka, P.; Stitt, M.; Moll, E.; Seemüller, E. Effect of phytoplasmal infection on concentration and translocation of carbohydrates and amino acids in periwinkle and tobacco. Physiol. Mol. Plant Pathol. 1999, 55, 59–68. [Google Scholar] [CrossRef]
- Marco, F.; Batailler, B.; Thorpe, M.R.; Razan, F.; Le Hir, R.; Vilaine, F.; Bouchereau, A.; Martin-Magniette, M.L.; Eveillard, S.; Dinant, S. Involvement of SUT1 and SUT2 sugar transporters in the impairment of sugar transport and changes in phloem exudate contents in phytoplasma-infected plants. Int. J. Mol. Sci. 2021, 22, 745. [Google Scholar] [CrossRef] [PubMed]
- Morkunas, I.; Ratajczak, L. The role of sugar signaling in plant defense responses against fungal pathogens. Acta Physiol. Plant 2014, 36, 1607–1619. [Google Scholar] [CrossRef]
- Tauzin, A.S.; Giardina, T. Sucrose and invertases, a part of the plant defense response to the biotic stresses. Front. Plant Sci. 2014, 5, 293. [Google Scholar] [CrossRef] [PubMed]
- Gayler, K.; Glasziou, K. Physiological functions of acid and neutral invertases in growth and sugar storage in sugar cane. Physiol. Plant. 1972, 27, 25–31. [Google Scholar] [CrossRef]
- Koch, K. Sucrose metabolism: Regulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr. Opin. Plant Biol. 2004, 7, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Christensen, N.M.; Nicolaisen, M.; Hansen, M.; Schulz, A. Distribution of phytoplasmas in infected plants as revealed by real-time PCR and bioimaging. Mol. Plant Microbe Interact. 2004, 17, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Musetti, R.; Paolacci, A.; Ciaffi, M.; Tanzarella, O.A.; Polizzotto, R.; Tubaro, F.; Mizzau, M.; Ermacora, P.; Badiani, M.; Osler, R. Phloem cytochemical modification and gene expression following the recovery of apple plants from apple proliferation disease. Phytopathology 2010, 100, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Tena, G.; Boudsocq, M.; Sheen, J. Protein kinase signaling networks in plant innate immunity. Curr. Opin. Plant Biol. 2011, 14, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, M.; Liu, Z.; Dai, L.; Zhang, M.; Wang, L.; Zhao, J.; Liu, M. Genome-wide identification of CNGC genes in Chinese jujube (Ziziphus jujuba Mill.) and ZjCNGC2 mediated signalling cascades in response to cold stress. BMC Genom. 2020, 21, 191. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Fan, G.; Li, X. Genome-wide analysis of three histone marks and gene expression in Paulownia fortunei with phytoplasma infection. BMC Genom. 2019, 20, 234. [Google Scholar] [CrossRef] [PubMed]
- Mardi, M.; Farsad, L.K.; Gharechahi, J.; Salekdeh, G.H. In-depth transcriptome sequencing of Mexican lime trees infected with Candidatus Phytoplasma aurantifolia. PLoS ONE 2015, 10, e0130425. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, A.; Jayaraman, D.; Grau, C.; Hill, J.H.; Whitham, S.A.; Ane, J.M.; Smith, D.L.; Kabbage, M. The pathogenic development of Sclerotinia sclerotiorum in soybean requires specific host NADPH oxidases. Mol. Plant Pathol. 2018, 19, 700–714. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.-Y.; Jiao, J.-L.; Wang, W.-W.; Jie, X.-R.; Wang, H.-Z. Silencing of the calcium-dependent protein kinase TaCDPK27 improves wheat resistance to powdery mildew. BMC Plant Biol. 2023, 23, 134. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.-J.; Li, P.; Shimono, M.; Corrion, A.; Higaki, T.; He, S.Y.; Day, B. Arabidopsis calcium-dependent protein kinase 3 regulates actin cytoskeleton organization and immunity. Nat. Commun. 2020, 11, 6234. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhao, Z.; Xue, C.; Wang, L.; Wang, L.; Feng, C.; Zhang, L.; Yu, Z.; Zhao, J.; Liu, M. Three main genes in the MAPK cascade involved in the Chinese jujube-phytoplasma interaction. Forests 2019, 10, 392. [Google Scholar] [CrossRef]
- Yang, S.H.; Sharrocks, A.D.; Whitmarsh, A.J. Transcriptional regulation by the MAP kinase signaling cascades. Gene 2003, 320, 3–21. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, C.; Sadanandom, A.; Kitagawa, K.; Freialdenhoven, A.; Shirasu, K.; Schulze-Lefert, P. The RAR1 interactor SGT1, an essential component of R gene-triggered disease resistance. Science 2002, 295, 2073–2076. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, S.; Li, M.; Su, Y.; Sun, Z.; Ma, H. Combined transcriptome and metabolome analysis of Nerium indicum L. elaborates the key pathways that are activated in response to witches’ broom disease. BMC Plant Biol. 2022, 22, 291. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Warren, R.F.; Zhao, T.; Shan, L.; Zhu, L.; Tang, X.; Zhou, J.M. Overexpression of Pti5 in tomato potentiates pathogen-induced defense gene expression and enhances disease resistance to Pseudomonas syringae pv. tomato. Mol. Plant Microbe Interact. 2001, 14, 1453–1457. [Google Scholar] [CrossRef] [PubMed]
- Agrios, G. How plants defend themselves against pathogens. In Plant Pathology; Elsevier: Amsterdam, The Netherlands, 2005; pp. 207–248. [Google Scholar] [CrossRef]
- Tai, C.F.; Lin, C.P.; Sung, Y.C.; Chen, J.C. Auxin influences symptom expression and phytoplasma colonisation in periwinkle infected with periwinkle leaf yellowing phytoplasma. Ann. Appl. Biol. 2013, 163, 420–429. [Google Scholar] [CrossRef]
- Lu, Y.T.; Li, M.Y.; Cheng, K.T.; Tan, C.M.; Su, L.W.; Lin, W.Y.; Shih, H.T.; Chiou, T.J.; Yang, J.Y. Transgenic plants that express the phytoplasma effector SAP11 show altered phosphate starvation and defense responses. Plant Physiol. 2014, 164, 1456–1469. [Google Scholar] [CrossRef] [PubMed]
- Musetti, R.; Farhan, K.; De Marco, F.; Polizzotto, R.; Paolacci, A.; Ciaffi, M.; Ermacora, P.; Grisan, S.; Santi, S.; Osler, R. Differentially-regulated defence genes in Malus domestica during phytoplasma infection and recovery. Eur. J. Plant Pathol. 2013, 136, 13–19. [Google Scholar] [CrossRef]
- Paolacci, A.R.; Catarcione, G.; Ederli, L.; Zadra, C.; Pasqualini, S.; Badiani, M.; Musetti, R.; Santi, S.; Ciaffi, M. Jasmonate-mediated defence responses, unlike salicylate-mediated responses, are involved in the recovery of grapevine from bois noir disease. BMC Plant Biol. 2017, 17, 118. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhao, J.; Liu, P.; Xing, H.; Li, C.; Wei, G.; Kang, Z. Glycerol-3-phosphate metabolism in wheat contributes to systemic acquired resistance against Puccinia striiformis f. sp. tritici. PLoS ONE 2013, 8, e81756. [Google Scholar] [CrossRef] [PubMed]
- Hsu, F.C.; Chou, M.Y.; Chou, S.J.; Li, Y.R.; Peng, H.P.; Shih, M.C. Submergence confers immunity mediated by the WRKY22 transcription factor in arabidopsis. Plant Cell 2013, 25, 2699–2713. [Google Scholar] [CrossRef] [PubMed]
- Ntambo, M.S.; Meng, J.Y.; Rott, P.C.; Henry, R.J.; Zhang, H.L.; Gao, S.J. Comparative transcriptome profiling of resistant and susceptible sugarcane cultivars in response to infection by Xanthomonas albilineans. Int. J. Mol. Sci. 2019, 20, 6138. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Tang, X.; Martin, G.B. The Pto kinase conferring resistance to tomato bacterial speck disease interacts with proteins that bind a cis-element of pathogenesis-related genes. EMBO J. 1997, 16, 3207–3218. [Google Scholar] [CrossRef] [PubMed]
- Meddya, S.; Meshram, S.; Sarkar, D.; S, R.; Datta, R.; Singh, S.; Avinash, G.; Kumar Kondeti, A.; Savani, A.K.; Thulasinathan, T. Plant stomata: An unrealized possibility in plant defense against invading pathogens and stress tolerance. Plants 2023, 12, 3380. [Google Scholar] [CrossRef] [PubMed]
- Ana, L.-M.; Rogelio, S.; Xose Carlos, S.; Rosa Ana, M. Cell wall composition impacts structural characteristics of the stems and thereby the biomass yield. J. Agric. Food Chem. 2022, 70, 3136–3141. [Google Scholar] [CrossRef] [PubMed]
- De León, I.P.; Montesano, M. Activation of defense mechanisms against pathogens in mosses and flowering plants. Int. J. Mol. Sci. 2013, 14, 3178–3200. [Google Scholar] [CrossRef] [PubMed]
- Hren, M.; Ravnikar, M.; Brzin, J.; Ermacora, P.; Carraro, L.; Bianco, P.; Casati, P.; Borgo, M.; Angelini, E.; Rotter, A. Induced expression of sucrose synthase and alcohol dehydrogenase I genes in phytoplasma-infected grapevine plants grown in the field. Plant Pathol. 2009, 58, 170–180. [Google Scholar] [CrossRef]
- Fan, G.; Xu, E.; Deng, M.; Zhao, Z.; Niu, S. Phenylpropanoid metabolism, hormone biosynthesis and signal transduction-related genes play crucial roles in the resistance of Paulownia fortunei to paulownia witches’ broom phytoplasma infection. Genes. Genom. 2015, 37, 913–929. [Google Scholar] [CrossRef]
- Sugio, A.; Kingdom, H.N.; MacLean, A.M.; Grieve, V.M.; Hogenhout, S.A. Phytoplasma protein effector SAP11 enhances insect vector reproduction by manipulating plant development and defense hormone biosynthesis. Proc. Natl. Acad. Sci. USA 2011, 108, E1254–E1263. [Google Scholar] [CrossRef] [PubMed]
- Schommer, C.; Palatnik, J.F.; Aggarwal, P.; Chetelat, A.; Cubas, P.; Farmer, E.E.; Nath, U.; Weigel, D. Control of jasmonate biosynthesis and senescence by miR319 targets. PLoS Biol. 2008, 6, e230. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Yu, H.; Yuan, R.; Yang, Y.; An, F.; Qin, G. Arabidopsis transcription factor TCP5 controls plant thermomorphogenesis by positively regulating PIF4 activity. iScience 2019, 15, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Dermastia, M.; Nikolic, P.; Chersicola, M.; Gruden, K. Transcriptional profiling in infected and recovered grapevine plant responses to ‘Candidatus Phytoplasma solani’. Phytopathog. Mollicutes 2015, 5, S123–S124. [Google Scholar] [CrossRef]
- Giorno, F.; Guerriero, G.; Biagetti, M.; Ciccotti, A.M.; Baric, S. Gene expression and biochemical changes of carbohydrate metabolism in in vitro micro-propagated apple plantlets infected by ‘Candidatus Phytoplasma mali’. Plant Physiol. Biochem. 2013, 70, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.Y.; Tseng, H.I.; Lin, C.P.; Lin, Y.Y.; Huang, Y.H.; Huang, C.K.; Chang, T.H.; Lin, S.S. High-throughput transcriptome analysis of the leafy flower transition of Catharanthus roseus induced by peanut witches’-broom phytoplasma infection. Plant Cell Physiol. 2014, 55, 942–957. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Dong, X. Systemic acquired resistance: Turning local infection into global defense. Annu. Rev. Plant Biol. 2013, 64, 839–863. [Google Scholar] [CrossRef] [PubMed]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D. Hormone crosstalk in plant disease and defense: More than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Mou, H.Q.; Lu, J.; Zhu, S.F.; Lin, C.L.; Tian, G.Z.; Xu, X.; Zhao, W.J. Transcriptomic analysis of paulownia infected by paulownia witches’-broom phytoplasma. PLoS ONE 2013, 8, e77217. [Google Scholar] [CrossRef] [PubMed]
- Stratilova, B.; Kozmon, S.; Stratilova, E.; Hrmova, M. Plant xyloglucan xyloglucosyl transferases and the cell wall structure: Subtle but significant. Molecules 2020, 25, 5619. [Google Scholar] [CrossRef] [PubMed]
- Geng, D.; Shen, X.; Xie, Y.; Yang, Y.; Bian, R.; Gao, Y.; Li, P.; Sun, L.; Feng, H.; Ma, F.; et al. Regulation of phenylpropanoid biosynthesis by MdMYB88 and MdMYB124 contributes to pathogen and drought resistance in apple. Hortic. Res. 2020, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.J.; Reddy, M.S.; Wang, L. The phenylpropanoid pathway and plant defence-a genomics perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Ye, Z.H. MYB46 and MYB83 bind to the SMRE sites and directly activate a suite of transcription factors and secondary wall biosynthetic genes. Plant Cell Physiol. 2012, 53, 368–380. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Zhang, J.; Tschaplinski, T.J.; Tuskan, G.A.; Chen, J.G.; Muchero, W. Regulation of lignin biosynthesis and its role in growth-defense tradeoffs. Front. Plant Sci. 2018, 9, 1427. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, M.S.; Stevenson, P.C. Effects of isoflavonoids from Cicer on larvae of Heliocoverpa armigera. J. Chem. Ecol. 2001, 27, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Karre, S.; Kumar, A.; Yogendra, K.; Kage, U.; Kushalappa, A.; Charron, J.-B. HvWRKY23 regulates flavonoid glycoside and hydroxycinnamic acid amide biosynthetic genes in barley to combat Fusarium head blight. Plant Mol. Biol. 2019, 100, 591–605. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.M.; Zerbe, P. Specialized diterpenoid metabolism in monocot crops: Biosynthesis and chemical diversity. Phytochemistry 2020, 172, 112289. [Google Scholar] [CrossRef] [PubMed]
- Olszewski, N.; Sun, T.P.; Gubler, F. Gibberellin signaling: Biosynthesis, catabolism, and response pathways. Plant Cell 2002, 14, S61–S80. [Google Scholar] [CrossRef] [PubMed]
- Bruni, R.; Pellati, F.; Bellardi, M.G.; Benvenuti, S.; Paltrinieri, S.; Bertaccini, A.; Bianchi, A. Herbal drug quality and phytochemical composition of Hypericum perforatum L. affected by ash yellows phytoplasma infection. J. Agric. Food Chem. 2005, 53, 964–968. [Google Scholar] [CrossRef] [PubMed]
- Rid, M.; Mesca, C.; Ayasse, M.; Gross, J. Apple proliferation phytoplasma influences the pattern of plant volatiles emitted depending on pathogen virulence. Front. Ecol. Evol. 2016, 3, 152. [Google Scholar] [CrossRef]
- Honeycutt, R.J.; Sobral, B.W.S.; Keim, P.; Irvine, J.E. A rapid DNA extraction method for sugarcane and its relatives. Plant Mol. Biol. Rep. 1992, 10, 66–72. [Google Scholar] [CrossRef]
- Gai, Y.P.; Yuan, S.S.; Liu, Z.Y.; Zhao, H.N.; Liu, Q.; Qin, R.L.; Fang, L.J.; Ji, X.L. Integrated phloem sap mRNA and protein expression analysis reveals phytoplasma-infection responses in mulberry. Mol. Cell Proteom. 2018, 17, 1702–1719. [Google Scholar] [CrossRef] [PubMed]
- Staroscik, A. Calculator for Determining the Number of Copies of a Template. Available online: https://cels.uri.edu/gsc/cndna.html (accessed on 25 December 2019).
- Ikten, C.; Ustun, R.; Catal, M.; Yol, E.; Uzun, B. Multiplex real-time qPCR assay for simultaneous and sensitive detection of phytoplasmas in sesame plants and insect vectors. PLoS ONE 2016, 11, e0155891. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 3 October 2019).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. agriGO v2.0: A GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y. KEGG Mapper for inferring cellular functions from protein sequences. Protein Sci. 2020, 29, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Varemo, L.; Nielsen, J.; Nookaew, I. Enriching the gene set analysis of genome-wide data by incorporating directionality of gene expression and combining statistical hypotheses and methods. Nucleic Acids Res. 2013, 41, 4378–4391. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sugarcane | Height (cm) | Diameter (cm) | Sugar Content (°Brix) | 16S/18S rRNA Gene Ratio | |
|---|---|---|---|---|---|
| Leaves | Stalks | ||||
| Asymptom | 302.20 ± 12.77 | 3.34 ± 0.13 | 22.84 ± 0.25 | 2.35 × 10−5 ± 0.82 × 10−5 | 8.96 × 10−5 ± 3.00 × 10−5 |
| Symptom | 199.20 ± 16.76 * | 2.18 ± 0.16 * | 15.86 ± 1.14 * | 0.33 ± 0.10 * | 0.57 ± 0.14 * |
| Category | Number of DEGs | |
|---|---|---|
| Leaves | Stalks | |
| Significantly expressed gene (p-value < 0.05) | 6612 | 2173 |
| |log2Fold Change| > 1.0 | 6284 | 1737 |
| - Upregulated genes | 4799 | 994 |
| - Downregulate genes | 1485 | 743 |
| Gene ontology | 3711 (59.06%) | 987 (56.82%) |
| - Molecular function | 3350 | 901 |
| - Cellular component | 884 | 186 |
| - Biological process | 2523 | 675 |
| KEGG pathway | 2248 (35.79%) | 531 (30.57%) |
| - Upregulated DEGs | 1701 | 294 |
| - Downregulated DEGs | 547 | 237 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lohmaneeratana, K.; Leetanasaksakul, K.; Thamchaipenet, A. Transcriptomic Profiling of Sugarcane White Leaf (SCWL) Canes during Maturation Phase. Plants 2024, 13, 1551. https://doi.org/10.3390/plants13111551
Lohmaneeratana K, Leetanasaksakul K, Thamchaipenet A. Transcriptomic Profiling of Sugarcane White Leaf (SCWL) Canes during Maturation Phase. Plants. 2024; 13(11):1551. https://doi.org/10.3390/plants13111551
Chicago/Turabian StyleLohmaneeratana, Karan, Kantinan Leetanasaksakul, and Arinthip Thamchaipenet. 2024. "Transcriptomic Profiling of Sugarcane White Leaf (SCWL) Canes during Maturation Phase" Plants 13, no. 11: 1551. https://doi.org/10.3390/plants13111551
APA StyleLohmaneeratana, K., Leetanasaksakul, K., & Thamchaipenet, A. (2024). Transcriptomic Profiling of Sugarcane White Leaf (SCWL) Canes during Maturation Phase. Plants, 13(11), 1551. https://doi.org/10.3390/plants13111551