
Tomato Mutants Reveal Root and Shoot Strigolactone Involvement in Branching and Broomrape Resistance

Abstract
:
1. Introduction
2. Results
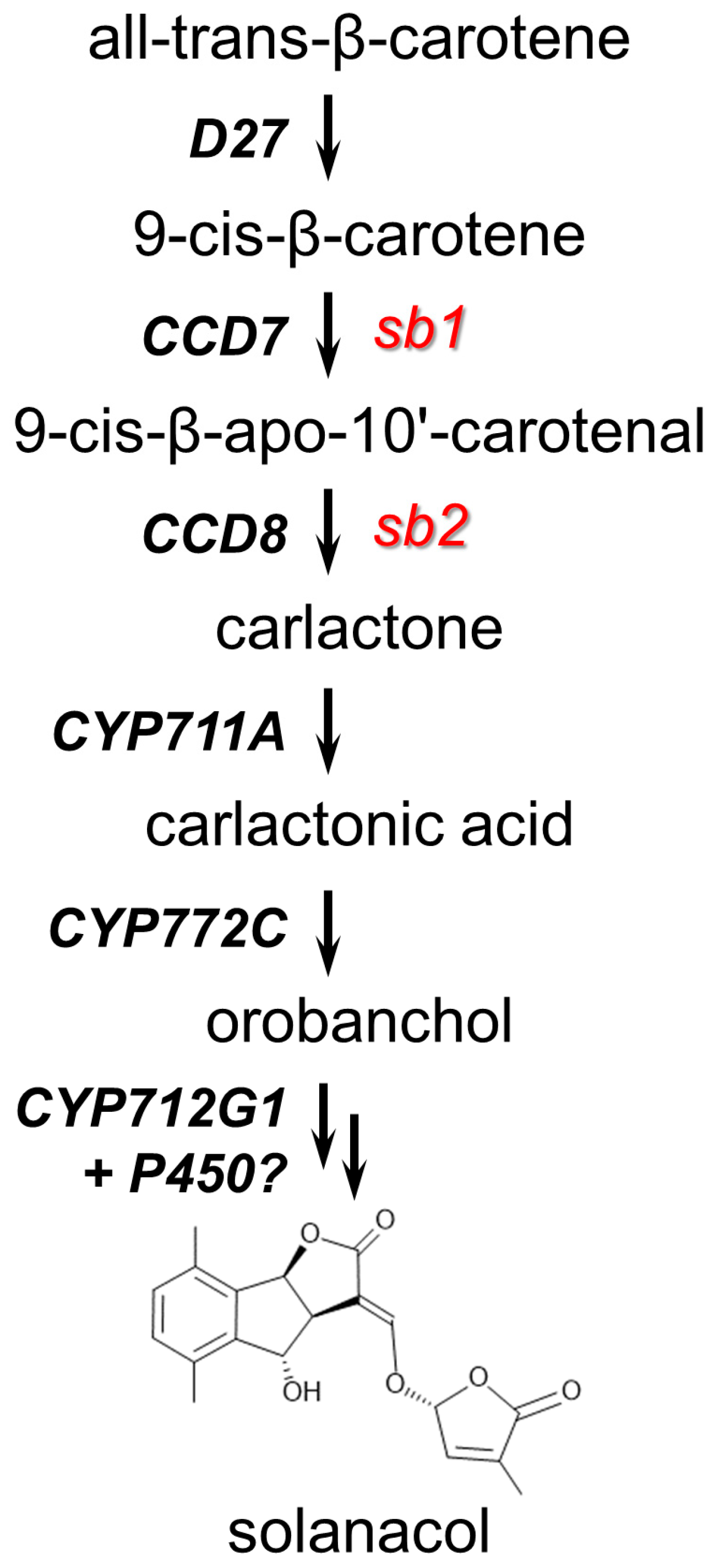
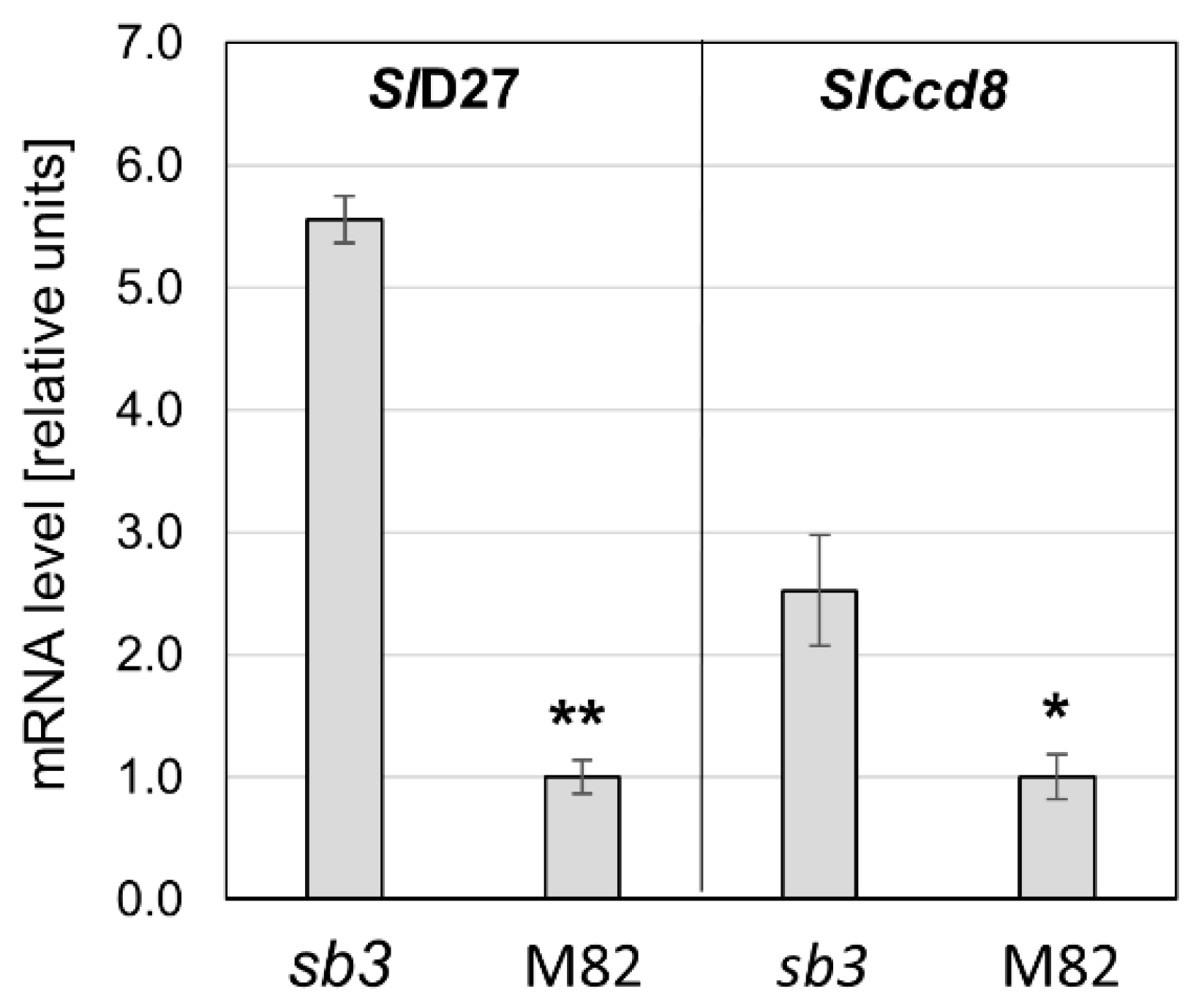
2.1. Isolation and Molecular Characterization of SL Mutants
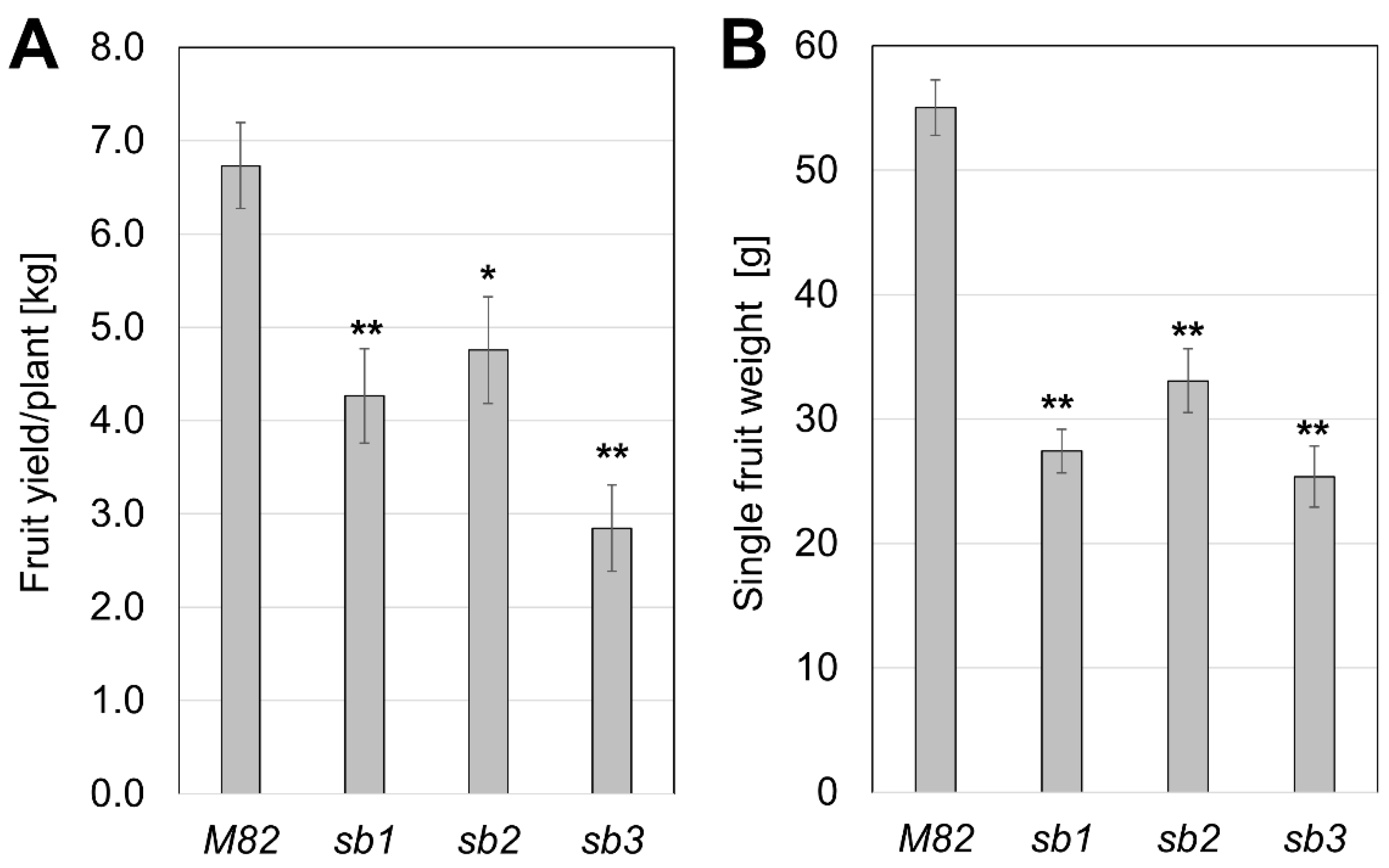
2.2. The Phenotype of sb Mutants in Grafting Experiments
3. Discussion
3.1. Isogenic Mutations in Strigolactone Synthesis and Perception
3.2. Graft Transmissible Effects of SL
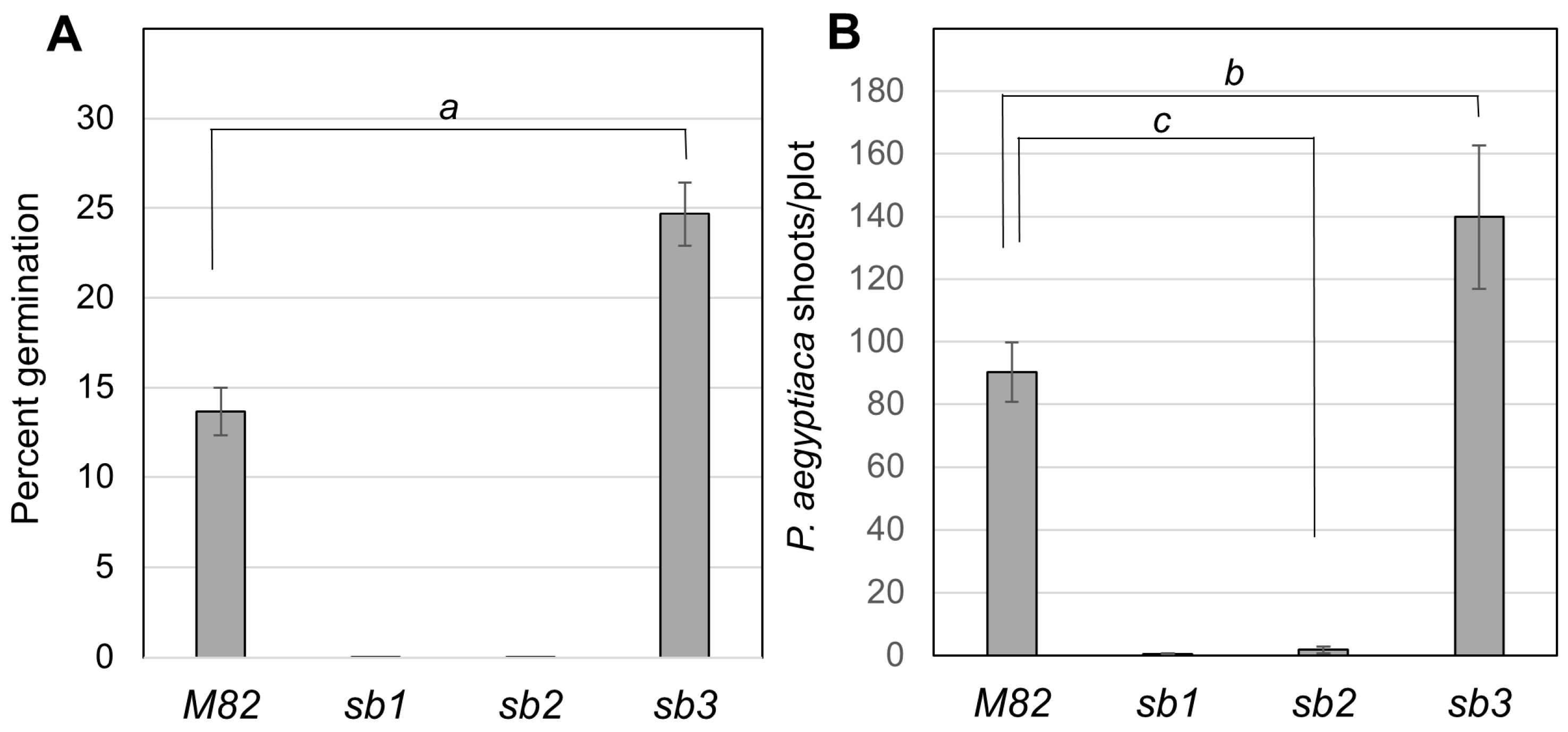
3.3. A Genetic Solution to Broomrape Infestation
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. DNA Extraction and Sequencing
4.3. RNA Extraction, cDNA Sequencing, and Measurement of mRNA with Quantitative Real-Time RT-PCR
4.4. Strigolactone Quantification Bioassay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cook, C.E.; Whichard, L.P.; Turner, B.; Wall, M.E.; Egley, G.H. Germination of witchweed (Striga lutea Lour.): Isolation and properties of a potent stimulant. Science 1966, 154, 1189–1190. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Yoneyama, K.; Yoneyama, K. The strigolactone story. Annu. Rev. Phytopathol. 2010, 48, 93–117. [Google Scholar] [CrossRef] [PubMed]
- Ruyter-Spira, C.; Al-Babili, S.; van der Krol, S.; Bouwmeester, H. The biology of strigolactones. Trends Plant Sci. 2012, 18, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Al-Babili, S.; Bouwmeester, H.J. Strigolactones, a novel carotenoid-derived plant hormone. Annu. Rev. Plant Biol. 2015, 66, 161–186. [Google Scholar] [CrossRef] [PubMed]
- Kyozuka, J.; Nomura, T.; Shimamura, M. Origins and evolution of the dual functions of strigolactones as rhizosphere signaling molecules and plant hormones. Curr. Opin. Plant Biol. 2022, 65, 102–154. [Google Scholar] [CrossRef] [PubMed]
- Umehara, M.; Hanada, A.; Yoshida, S.; Akiyama, K.; Arite, T.; Takeda-Kamiya, N.; Magome, H.; Kamiya, Y.; Shirasu, K.; Yoneyama, K.; et al. Inhibition of shoot branching by new terpenoid plant hormones. Nature 2008, 455, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.T.; Walter, M.H.; Giavalisco, P.; Lytovchenko, A.; Kohlen, W.; Charnikhova, T.; Simkin, A.J.; Goulet, C.; Strack, D.; Bouwmeester, H.J.; et al. SlCCD7 controls strigolactone biosynthesis, shoot branching and mycorrhiza-induced apocarotenoid formation in tomato. Plant J. 2010, 61, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Domagalska, M.A.; Leyser, O. Signal integration in the control of shoot branching. Nat. Rev. Mol. Cell Biol. 2011, 12, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Barbier, F.F.; Dun, E.A.; Kerr, S.C.; Chabikwa, T.G.; Beveridge, C.A. An update on the signals controlling shoot branching. Trends Plant Sci. 2019, 24, 220–236. [Google Scholar] [CrossRef]
- Koltai, H. Strigolactones are regulators of root development. New Phytol. 2011, 190, 545–549. [Google Scholar] [CrossRef]
- Khuvung, K.; Silva Gutierrez, F.A.O.; Reinhardt, D. How strigolactone shapes shoot architecture. Front. Plant Sci. 2022, 13, 889045. [Google Scholar] [CrossRef] [PubMed]
- Zwanenburg, B.; Blanco-Ania, D. Strigolactones: New plant hormones in the spotlight. J. Exp. Bot. 2018, 69, 2205–2218. [Google Scholar] [CrossRef] [PubMed]
- Waters, M.T.; Gutjahr, C.; Bennett, T.; Nelson, D.C. Strigolactone signaling and evolution. Annu. Rev. Plant Biol. 2017, 68, 291–322. [Google Scholar] [CrossRef] [PubMed]
- Trasoletti, M.; Visentin, I.; Campo, E.; Schubert, A.; Cardinale, F. Strigolactones as a hormonal hub for the acclimation and priming to environmental stress in plants. Plant Cell Environ. 2022, 45, 3611–3630. [Google Scholar] [CrossRef] [PubMed]
- Omoarelojie, L.O.; Kulkarni, M.G.; Finnie, J.F.; Van, S.J. Strigolactones and their crosstalk with other phytohormones. Ann. Bot. 2019, 124, 749–767. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Gao, Y.; Yang, W.; Sui, N.; Zhu, J. Biological functions of strigolactones and their crosstalk with other phytohormones. Front. Plant Sci. 2022, 13, 821563. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Raez, J.A.; Charnikhova, T.; Gomez-Roldan, V.; Matusova, R.; Kohlen, W.; De, V.R.; Verstappen, F.; Puech-Pages, V.; Becard, G.; Mulder, P.; et al. Tomato strigolactones are derived from carotenoids and their biosynthesis is promoted by phosphate starvation. New Phytol. 2008, 178, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Umehara, M.; Hanada, A.; Magome, H.; Takeda-Kamiya, N.; Yamaguchi, S. Contribution of strigolactones to the inhibition of tiller bud outgrowth under phosphate deficiency in rice. Plant Cell Physiol. 2010, 51, 1118–1126. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Fang, P.; Zhang, H.; Chi, C.; Song, L.; Xia, X.; Shi, K.; Zhou, Y.; Zhou, J.; Yu, J. Strigolactones positively regulate defense against root-knot nematodes in tomato. J. Exp. Bot. 2019, 70, 1325–1337. [Google Scholar] [CrossRef]
- Joel, D.M.; Gressel, J.; Musselman, L.J. Parasitic Orobanchaceae: Parasitic Mechanisms and Control Strategies; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1–513. [Google Scholar]
- Eizenberg, H.; Goldwasser, Y. Control of Egyptian broomrape in processing tomato: A summary of 20 years of research and successful implementation. Plant Dis. 2018, 102, 1477–1488. [Google Scholar] [CrossRef]
- Hershenhorn, J.; Eizenberg, H.; Dor, E.; Kapulnik, Y.; Goldwasser, Y. Phelipanche aegyptiaca management in tomato. Weed Res. 2009, 49, 34–47. [Google Scholar] [CrossRef]
- Matusova, R.; Rani, K.; Verstappen, F.W.; Franssen, M.C.; Beale, M.H.; Bouwmeester, H.J. The strigolactone germination stimulants of the plant-parasitic Striga and Orobanche spp. are derived from the carotenoid pathway. Plant Physiol. 2005, 139, 920–934. [Google Scholar] [CrossRef] [PubMed]
- Jia, K.P.; Baz, L.; Al-Babili, S. From carotenoids to strigolactones. J. Exp. Bot. 2018, 69, 2189–2204. [Google Scholar] [CrossRef] [PubMed]
- Bonhomme, S.; Guillory, A. Synthesis and signalling of strigolactone and KAI2-ligand signals in bryophytes. J. Exp. Bot. 2022, 73, 4487–4495. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, K.; Brewer, P.B. Strigolactones, how are they synthesized to regulate plant growth and development? Curr. Opin. Plant Biol. 2021, 63, 102072. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, T.; Hamana, M.; Mori, A.; Akiyama, R.; Ueno, K.; Osakabe, K.; Osakabe, Y.; Suzuki, H.; Takikawa, H.; Mizutani, M.; et al. Direct conversion of carlactonoic acid to orobanchol by cytochrome P450 CYP722C in strigolactone biosynthesis. Sci. Adv. 2019, 5, eaax9067. [Google Scholar] [CrossRef] [PubMed]
- Mashiguchi, K.; Seto, Y.; Yamaguchi, S. Strigolactone biosynthesis, transport and perception. Plant J. 2021, 105, 335–350. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, T.; Moriyama, D.; Miyamoto, A.; Okamura, H.; Shiotani, N.; Shimizu, N.; Mizutani, M.; Takikawa, H.; Sugimoto, Y. Identification of novel canonical strigolactones produced by tomato. Front. Plant Sci. 2022, 13, 1064378. [Google Scholar] [CrossRef]
- Wang, Y.; Durairaj, J.; Suárez Duran, H.G.; van Velzen, R.; Flokova, K.; Liao, C.Y.; Chojnacka, A.; MacFarlane, S.; Schranz, M.E.; Medema, M.H.; et al. The tomato cytochrome P450 CYP712G1 catalyses the double oxidation of orobanchol en route to the rhizosphere signalling strigolactone, solanacol. New Phytol. 2022, 235, 1884–1899. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.; Ming, Z.; Yan, L.; Li, S.; Wang, F.; Ma, S.; Yu, C.; Yang, M.; Chen, L.; Chen, L.; et al. DWARF14 is a non-canonical hormone receptor for strigolactone. Nature 2016, 536, 469–473. [Google Scholar] [CrossRef]
- Seto, Y.; Yasui, R.; Kameoka, H.; Tamiru, M.; Cao, M.M.; Terauchi, R.; Sakurada, A.; Hirano, R.; Kisugi, T.; Hanada, A.; et al. Strigolactone perception and deactivation by a hydrolase receptor DWARF14. Nat. Commun. 2019, 10, 191. [Google Scholar] [CrossRef]
- Wang, L.; Wang, B.; Yu, H.; Guo, H.; Lin, T.; Kou, L.; Wang, A.; Shao, N.; Ma, H.; Xiong, G.; et al. Transcriptional regulation of strigolactone signalling in Arabidopsis. Nature 2020, 583, 277–281. [Google Scholar] [CrossRef]
- Bythell-Douglas, R.; Rothfels, C.J.; Stevenson, D.W.D.; Graham, S.W.; Wong, G.K.; Nelson, D.C.; Bennett, T. Evolution of strigolactone receptors by gradual neo-functionalization of KAI2 paralogues. BMC Biol. 2017, 15, 52–0397. [Google Scholar] [CrossRef]
- Li, Z.; Pi, Y.; Zhai, C.; Xu, D.; Ma, W.; Chen, H.; Li, Y.; Wu, H. The strigolactone receptor SlDWARF14 plays a role in photosynthetic pigment accumulation and photosynthesis in tomato. Plant Cell Rep. 2022, 41, 2089–2105. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Aparicio, M.; Delavault, P.; Timko, M.P. Management of infection by parasitic weeds: A review. Plants 2020, 9, 1184. [Google Scholar] [CrossRef] [PubMed]
- Kountche, B.A.; Jamil, M.; Yonli, D.; Nikiema, M.P.; Blanco-Ania, D.; Asami, T.; Zwanenburg, B.; Al-Babili, S. Suicidal germination as a control strategy for Striga hermonthica (Benth.) in smallholder farms of sub-Saharan Africa. Plants People Planet 2019, 1, 107–118. [Google Scholar] [CrossRef]
- Dor, E.; Alperin, B.; Wininger, S.; Ben-Dor, B.; Somvanshi, V.S.; Koltai, H.; Kapulnik, Y.; Hershenhorn, J. Characterization of a novel tomato mutant resistant to the weedy parasites Orobanche and Phelipanche spp. Euphytica 2010, 171, 371–380. [Google Scholar] [CrossRef]
- Dor, E.; Yoneyama, K.; Wininger, S.; Kapulnik, Y.; Yoneyama, K.; Koltai, H.; Xie, X.; Hershenhorn, J. Strigolactone deficiency confers resistance in tomato line SL-ORT1 to the parasitic weeds Phelipanche and Orobanche spp. Phytopathology 2011, 101, 213–222. [Google Scholar] [CrossRef]
- Hasegawa, S.; Tsutsumi, T.; Fukushima, S.; Okabe, Y.; Saito, J.; Katayama, M.; Shindo, M.; Yamada, Y.; Shimomura, K.; Yoneyama, K.; et al. Low infection of Phelipanche aegyptiaca in Micro-Tom mutants deficient in CAROTENOIDCLEAVAGE DIOXYGENASE 8. Int. J. Mol. Sci. 2018, 19, 2645. [Google Scholar] [CrossRef]
- Bai, J.; Wei, Q.; Shu, J.; Gan, Z.; Li, B.; Yan, D.; Huang, Z.; Guo, Y.; Wang, X.; Zhang, L.; et al. Exploration of resistance to Phelipanche aegyptiaca in tomato. Pest. Manag. Sci. 2020, 76, 3806–3821. [Google Scholar] [CrossRef]
- Bari, V.K.; Nassar, J.A.; Aly, R. CRISPR/Cas9 mediated mutagenesis of MORE AXILLARY GROWTH 1 in tomato confers resistance to root parasitic weed Phelipanche aegyptiaca. Sci. Rep. 2021, 11, 3905–82897. [Google Scholar] [CrossRef]
- Galili, S.; Hershenhorn, J.; Smirnov, E.; Yoneyama, K.; Xie, X.; Amir-Segev, O.; Bellalou, A.; Dor, E. Characterization of a chickpea mutant resistant to Phelipanche aegyptiaca Pers. and Orobanche crenata Forsk. Plants 2021, 10, 2552. [Google Scholar] [CrossRef] [PubMed]
- Aly, R.; Matzrafi, M.; Bari, V.K. Using biotechnological approaches to develop crop resistance to root parasitic weeds. Planta 2021, 253, 97. [Google Scholar] [CrossRef]
- Menda, N.; Semel, Y.; Peled, D.; Eshed, Y.; Zamir, D. In silico screening of a saturated mutation library of tomato. Plant J. 2004, 38, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Koltai, H.; LekKala, S.P.; Bhattacharya, C.; Mayzlish-Gati, E.; Resnick, N.; Wininger, S.; Dor, E.; Yoneyama, K.; Yoneyama, K.; Hershenhorn, J.; et al. A tomato strigolactone-impaired mutant displays aberrant shoot morphology and plant interactions. J. Exp. Bot. 2010, 61, 1739–1749. [Google Scholar] [CrossRef]
- Kohlen, W.; Charnikhova, T.; Liu, Q.; Bours, R.; Domagalska, M.A.; Beguerie, S.; Verstappen, F.; Leyser, O.; Bouwmeester, H.; Ruyter-Spira, C. Strigolactones are transported through the xylem and play a key role in shoot architectural response to phosphate deficiency in nonarbuscular mycorrhizal host Arabidopsis. Plant Physiol. 2011, 155, 974–987. [Google Scholar] [CrossRef]
- Bari, V.K.; Nassar, J.A.; Kheredin, S.M.; Gal-On, A.; Ron, M.; Britt, A.; Steele, D.; Yoder, J.; Aly, R. CRISPR/Cas9-mediated mutagenesis of CAROTENOID CLEAVAGE DIOXYGENASE 8 in tomato provides resistance against the parasitic weed Phelipanche aegyptiaca. Sci. Rep. 2019, 9, 11438–47893. [Google Scholar] [CrossRef]
- Burger, M.; Chory, J. The many models of strigolactone signaling. Trends Plant Sci. 2020, 25, 395–405. [Google Scholar] [CrossRef]
- Batra, R.; Agarwal, P.; Tyagi, S.; Saini, D.K.; Kumar, V.; Kumar, A.; Kumar, S.; Balyan, H.S.; Pandey, R.; Gupta, P.K. A study of CCD8 genes/proteins in seven monocots and eight dicots. PLoS ONE 2019, 14, e0213531. [Google Scholar] [CrossRef]
- Kretzschmar, T.; Kohlen, W.; Sasse, J.; Borghi, L.; Schlegel, M.; Bachelier, J.B.; Reinhardt, D.; Bours, R.; Bouwmeester, H.J.; Martinoia, E. A petunia ABC protein controls strigolactone-dependent symbiotic signalling and branching. Nature 2012, 483, 341–344. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, X.; Wang, Y.; Díez-Simón, C.; Flokova, K.; Bimbo, A.; Bouwmeester, H.J.; Ruyter-Spira, C. The tomato MAX1 homolog, SlMAX1, is involved in the biosynthesis of tomato strigolactones from carlactone. New Phytol. 2018, 219, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Stirnberg, P.; Furner, I.J.; Ottoline Leyser, H.M. MAX2 participates in an SCF complex which acts locally at the node to suppress shoot branching. Plant J. 2007, 50, 80–94. [Google Scholar] [CrossRef] [PubMed]
- Arite, T.; Umehara, M.; Ishikawa, S.; Hanada, A.; Maekawa, M.; Yamaguchi, S.; Kyozuka, J. d14, a strigolactone-insensitive mutant of rice, shows an accelerated outgrowth of tillers. Plant Cell Physiol. 2009, 50, 1416–1424. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Lin, Q.; Zhu, L.; Ren, Y.; Zhou, K.; Shabek, N.; Wu, F.; Mao, H.; Dong, W.; Gan, L.; et al. D14-SCF(D3)-dependent degradation of D53 regulates strigolactone signalling. Nature 2013, 504, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Johnson, X.; Brcich, T.; Dun, E.A.; Goussot, M.; Haurogné, K.; Beveridge, C.A.; Rameau, C. Branching genes are conserved across species. Genes controlling a novel signal in pea are coregulated by other long-distance signals. Plant Physiol. 2006, 142, 1014–1026. [Google Scholar] [CrossRef] [PubMed]
- Mashiguchi, K.; Sasaki, E.; Shimada, Y.; Nagae, M.; Ueno, K.; Nakano, T.; Yoneyama, K.; Suzuki, Y.; Asami, T. Feedback-regulation of strigolactone biosynthetic genes and strigolactone-regulated genes in Arabidopsis. Biosci. Biotechnol Biochem. 2009, 73, 2460–2465. [Google Scholar] [CrossRef] [PubMed]
- Mashiguchi, K.; Morita, R.; Tanaka, K.; Kodama, K.; Kameoka, H.; Kyozuka, J.; Seto, Y.; Yamaguchi, S. Activation of strigolactone biosynthesis by the DWARF14-LIKE/KARRIKIN-INSENSITIVE2 pathway in mycorrhizal angiosperms, but Not in Arabidopsis, a non-mycorrhizal plant. Plant Cell Physiol. 2023, 64, 1066–1078. [Google Scholar] [CrossRef] [PubMed]
- Kameoka, H.; Kyozuka, J. Spatial regulation of strigolactone function. J. Exp. Bot. 2018, 69, 2255–2264. [Google Scholar] [CrossRef]
- Beveridge, C.A.; Symons, G.M.; Turnbull, C.G. Auxin inhibition of decapitation-induced branching is dependent on graft-transmissible signals regulated by genes Rms1 and Rms2. Plant Physiol. 2000, 123, 689–698. [Google Scholar] [CrossRef]
- Morris, S.E.; Turnbull, C.G.; Murfet, I.C.; Beveridge, C.A. Mutational analysis of branching in pea. Evidence that Rms1 and Rms5 regulate the same novel signal. Plant Physiol. 2001, 126, 1205–1213. [Google Scholar] [CrossRef]
- Simons, J.L.; Napoli, C.A.; Janssen, B.J.; Plummer, K.M.; Snowden, K.C. Analysis of the DECREASED APICAL DOMINANCE genes of petunia in the control of axillary branching. Plant Physiol. 2007, 143, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Mashiguchi, K.; Seto, Y.; Onozuka, Y.; Suzuki, S.; Takemoto, K.; Wang, Y.; Dong, L.; Asami, K.; Noda, R.; Kisugi, T.; et al. A carlactonoic acid methyltransferase that contributes to the inhibition of shoot branching in Arabidopsis. Proc. Natl. Acad. Sci. USA 2022, 119, e2111565119. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.M.; Ledger, S.E.; Janssen, B.J.; Luo, Z.; Drummond, R.S.M.; Tomes, S.; Karunairetnam, S.; Waite, C.N.; Funnell, K.A.; van Hooijdonk, B.M.; et al. Expression of MdCCD7 in the scion determines the extent of sylleptic branching and the primary shoot growth rate of apple trees. J. Exp. Bot. 2018, 69, 2379–2390. [Google Scholar] [CrossRef]
- Beveridge, T.J.; Kadurugamuwa, J.L. Periplasm, periplasmic spaces, and their relation to bacterial wall structure: Novel secretion of selected periplasmic proteins from Pseudomonas aeruginosa. Microb. Drug Resist. 1996, 2, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kameoka, H.; Dun, E.A.; Lopez-Obando, M.; Brewer, P.B.; de Saint, G.A.; Rameau, C.; Beveridge, C.A.; Kyozuka, J. Phloem transport of the receptor DWARF14 protein Is required for full function of strigolactones. Plant Physiol. 2016, 172, 1844–1852. [Google Scholar] [CrossRef]
- Chevalier, F.; Nieminen, K.; Sánchez-Ferrero, J.C.; Rodríguez, M.L.; Chagoyen, M.; Hardtke, C.S.; Cubas, P. Strigolactone promotes degradation of DWARF14, an a/b hydrolase essential for strigolactone signaling in Arabidopsis. Plant Cell. 2014, 26, 1134–1150. [Google Scholar] [CrossRef]
- Gobena, D.; Shimels, M.; Rich, P.J.; Ruyter-Spira, C.; Bouwmeester, H.; Kanuganti, S.; Mengiste, T.; Ejeta, G. Mutation in sorghum LOW GERMINATION STIMULANT 1 alters strigolactones and causes Striga resistance. Proc. Natl. Acad. Sci. USA 2017, 114, 4471–4476. [Google Scholar] [CrossRef]
- Li, C.; Dong, L.; Durairaj, J.; Guan, J.C.; Yoshimura, M.; Quinodoz, P.; Horber, R.; Gaus, K.; Li, J.; Setotaw, Y.B.; et al. Maize resistance to witchweed through changes in strigolactone biosynthesis. Science 2023, 379, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Pavan, S.; Schiavulli, A.; Marcotrigiano, A.R.; Bardaro, N.; Bracuto, V.; Ricciardi, F.; Charnikhova, T.; Lotti, C.; Bouwmeester, H.; Ricciardi, L. Characterization of low-strigolactone germplasm in pea (Pisum sativum L.) resistant to crenate broomrape (Orobanche crenata Forsk.). Mol. Plant Microbe Interact. 2016, 29, 743–749. [Google Scholar] [CrossRef]
- Eshed, Y.; Zamir, D. An introgression line population of Lycopersicon pennellii in the cultivated tomato enables the identification and fine mapping of yield-associated QTL. Genetics 1995, 141, 1147–1162. [Google Scholar] [CrossRef]
- Sun, Z.; Hans, J.; Walter, M.H.; Matusova, R.; Beekwilder, J.; Verstappen, F.W.; Ming, Z.; van Echtelt, E.; Strack, D.; Bisseling, T.; et al. Cloning and characterisation of a maize carotenoid cleavage dioxygenase (ZmCCD1) and its involvement in the biosynthesis of apocarotenoids with various roles in mutualistic and parasitic interactions. Planta 2008, 228, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Chen, G.E.; Jamil, M.; Braguy, J.; Sioud, S.; Liew, K.X.; Balakrishna, A.; Al-Babili, S. Protocol for characterizing strigolactones released by plant roots. STAR Protoc. 2022, 3, 101352. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Locus | Mutation | Impact of the Mutation |
|---|---|---|---|
| Carotenoid cleavage dioxygenase 7 (SlCcd7) Solyc01g090660 | sb1 | G→C Position 3268 G→T position 3272 | Intron/exon #7 junction, deletion of 7bp in the mRNA of Ccd7 exon #7 |
| Carotenoid cleavage dioxygenase 8 (SlCcd8) Solyc08g066650 | sb2 | G→A Position 2659 | A change from Glu529 to Lys |
| α/β-hydrolase receptor Dwarf14 (D14) Solyc04g077860 | sb3 | G→A position 1582 | Intron/exon #2 junction, deletion of 17 bp in the mRNA of DWARF14 exon #2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karniel, U.; Koch, A.; Bar Nun, N.; Zamir, D.; Hirschberg, J. Tomato Mutants Reveal Root and Shoot Strigolactone Involvement in Branching and Broomrape Resistance. Plants 2024, 13, 1554. https://doi.org/10.3390/plants13111554
Karniel U, Koch A, Bar Nun N, Zamir D, Hirschberg J. Tomato Mutants Reveal Root and Shoot Strigolactone Involvement in Branching and Broomrape Resistance. Plants. 2024; 13(11):1554. https://doi.org/10.3390/plants13111554
Chicago/Turabian StyleKarniel, Uri, Amit Koch, Nurit Bar Nun, Dani Zamir, and Joseph Hirschberg. 2024. "Tomato Mutants Reveal Root and Shoot Strigolactone Involvement in Branching and Broomrape Resistance" Plants 13, no. 11: 1554. https://doi.org/10.3390/plants13111554