
Field Evaluation of Rice Lines Derived from Suakoko 8 X Bao Thai for Iron Tolerance in the South Saharan African Farming System
 , , and
, , and
Abstract
:1. Introduction
2. Results
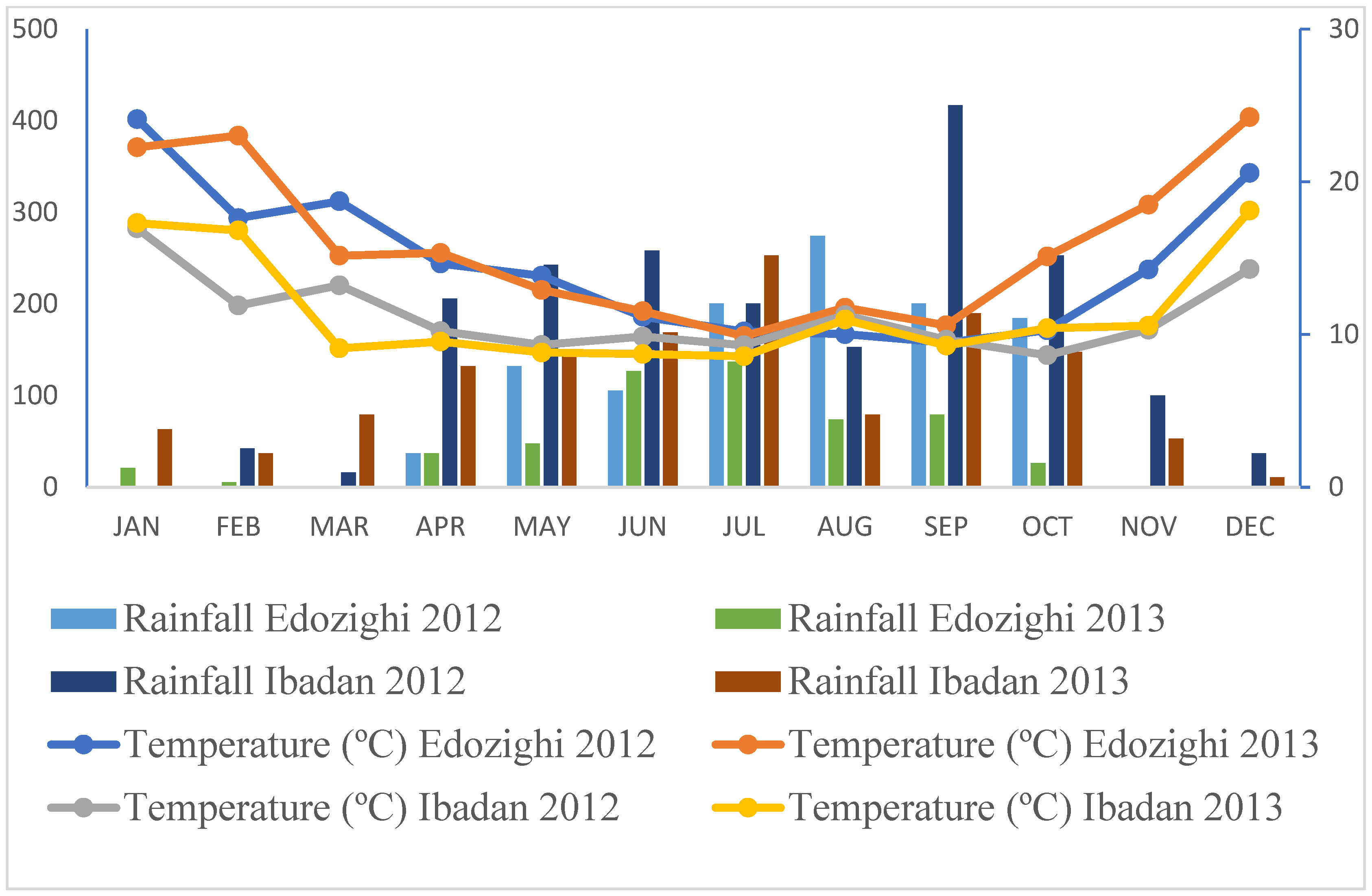
2.1. Weather Patterns at the Screening Sites
2.2. Probability Study
2.3. Mean Variabilities
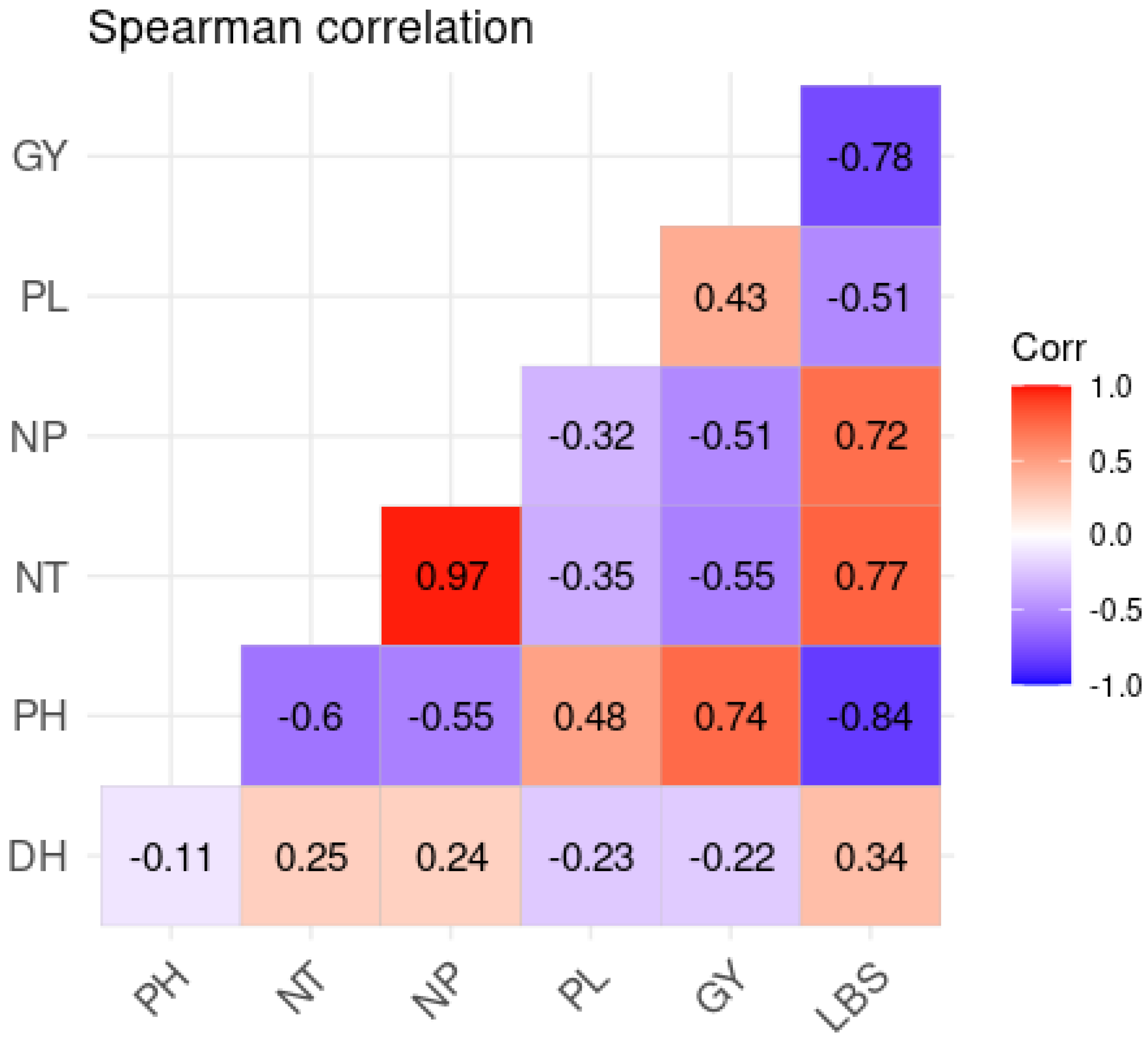
2.4. Correlation Analysis
2.5. Path Analysis
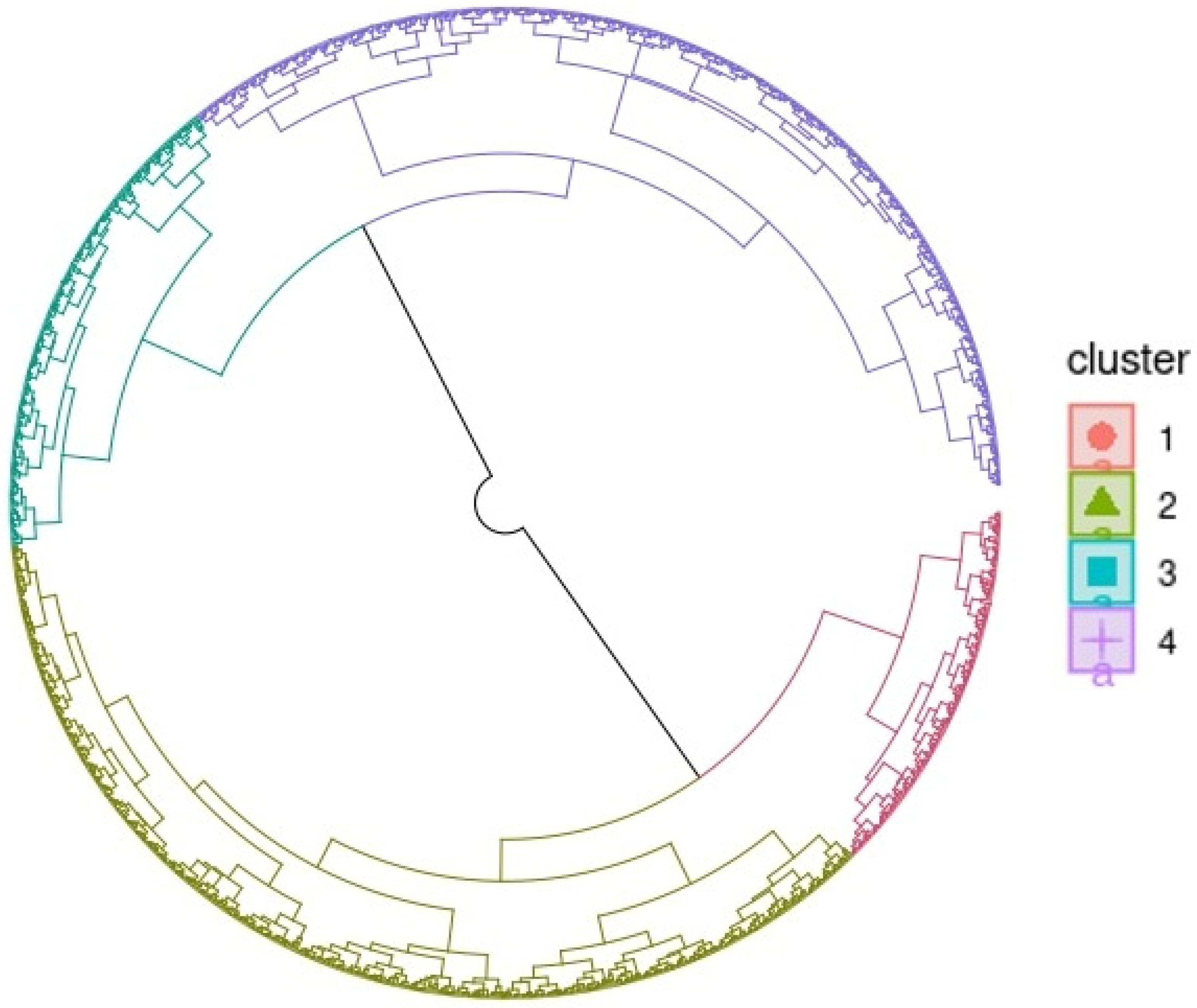
2.6. Classification of Genotypes and Principal Component Analysis
2.7. Determination of the Top 10 Performing Genotypes
3. Materials and Methods
3.1. Experimental Site and Genetic Materials
3.2. Field Management
3.3. Data Collection
3.4. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Oort, P. Balancing rice supply and demand in Africa. Nat. Food 2023, 4, 460–461. [Google Scholar] [CrossRef] [PubMed]
- Bocco, R.; Gandonou, C.B.; Amoussou, P.-L.; Togola, A.; Dieng, I.; Ndjiondjop, M.N.; Seck, P.A.; Tamo, M. Rapid phenotyping for identification of rice resistant varieties to Diopsis apicalis (Diptera: Diopsidae) Westwood. Cogent Biol. 2019, 5, 1649851. [Google Scholar]
- Africa Rice Center. Overview of Recent Developments in Sub-Saharan Africa Rice Trends. Cotonou, Benin, October 2007. Available online: https://www.eldis.org/document/A33885 (accessed on 31 January 2022).
- USDA (United States Department of Agriculture). Food Availability (Per Capita) Data System. 2014. Available online: http://www.ers.usda.gov (accessed on 11 March 2015).
- Fofana, I.; Goundan, A.; Magne Domgho, L.V. Impact Simulation of ECOWAS Rice Self-Sufficiency Policy; IFPRI Discussion Paper 01405; International Food Policy Research Institute (IFPRI): Washington, DC, USA, 2014; Available online: https://ebrary.ifpri.org/cdm/ref/collection/p15738coll2/id/128894 (accessed on 31 January 2022).
- ECOWAS (Economic Community of West African States Commission); WAEMU (West African Economic and Monetary Union Commission; NPCA (NEPAD Planning and Coordinating Agency). Accelerating the ECOWAP/CAADP implementation: Support for the implementation of the Regional Offensive for sustainable and sustained recovery of rice production in West Africa. In Proceedings of the Rice Task Force Meeting, Cotonou, Republic of Benin, 24–26 March 2014. [Google Scholar]
- del Villar, P.M.; Lançon, F. West African rice development: Beyond protectionism versus liberalization. Glob. Food Secur. 2015, 5, 56–61. [Google Scholar] [CrossRef]
- Fitzgerald, M.A.; McCouch, S.R.; Hall, R.D. Not just a grain of rice: The quest for quality. Trends Plant Sci. 2009, 14, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Sarma, B.; Kashtoh, H.; Lama Tamang, T.; Bhattacharyya, P.N.; Mohanta, Y.K.; Baek, K.H. Abiotic Stress in Rice: Visiting the Physiological Response and Its Tolerance Mechanisms. Plants 2023, 12, 3948. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, W.; Lan, P. The understanding of the plant iron deficiency responses in strategy I plants and the role of ethylene in this process by omic approaches. Front. Plant Sci. 2017, 8, 40. [Google Scholar] [CrossRef] [PubMed]
- Aung, M.S.; Masuda, H.; Kobayashi, T.; Nishizawa, N.K. Physiological and transcriptomic analysis of responses to different levels of iron excess stress in various rice tissues. Soil Sci. Plant Nutr. 2018, 64, 370–385. [Google Scholar] [CrossRef]
- Rasheed, A.; Hassan, M.U.; Aamer, M.; Bian, J.M.; Xu, Z.R.; He, X.; Yan, G.; Wu, Z. Iron toxicity, tolerance and quantitative trait loci mapping in rice: A review. Appl. Ecol. Environ. Res. 2020, 18, 7483–7498. [Google Scholar] [CrossRef]
- Mahender, A.; Swamy, B.P.M.; Anandan, A.; Ali, J. Tolerance of iron-deficient and -toxic soil conditions in rice. Plants 2019, 8, 31. [Google Scholar] [CrossRef]
- Guangjie, L.; Jinlin, W.; Kronzucker Herbert, J.; Baohai, L.; Weiming, S. Physiological and molecular mechanisms of plant-root responses to iron toxicity. J. Plant Physiol. 2024, 297, 154257. [Google Scholar] [CrossRef]
- Audebert, A.; Fofana, M. Rice yield gap due to iron toxicity in West Africa. J. Agron. Crop Sci. 2009, 195, 66–76. [Google Scholar] [CrossRef]
- Engel, K.; Asch, F.; Becker, M. Classification of rice genotypes based on their mechanisms of adaptation to iron toxicity. J. Plant Nutr. Soil Sci. 2012, 175, 871–881. [Google Scholar] [CrossRef]
- Sikirou, M.; Saito, K.; Dramé, K.N.; Saidou, A.; Dieng, I.; Ahanchédé, A.; Venuprasad, R. Soil-based screening for iron toxicity tolerance in rice using pots. Plant Prod. Sci. 2016, 19, 489–496. [Google Scholar] [CrossRef]
- Tisarum, R.; Pongprayoon, W.; Sithtisarn, S.; Sampumphuang, T.; Sotesaritkul, T.; Datta, A.; Singh, H.P.; Cha-Um, S. Expression levels of genes involved in metal homeostasis, physiological adaptation, and growth characteristics of rice (Oryza sativa L.) genotypes under Fe and/or Al toxicity. Protoplasma 2022, 259, 1013–1028. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Y.; Zeng, F.; Zhuang, J.; Yu, S.; Zhu, L.; Jin, Q.; Zhang, G. Genetic analysis of genotype × iron nutrition interaction on coleoptile elongation rate in rice (Oryza sativa L.). Euphytica 2007, 156, 311–318. [Google Scholar] [CrossRef]
- Dufey, I.; Draye, X.; Lutts, S.; Lorieux, M.; Martinez, C.; Bertin, P. Novel QTLs in an interspecific backcross Oryza sativa × Oryza glaberrima for resistance to iron toxicity in rice. Euphytica 2015, 204, 609–625. [Google Scholar] [CrossRef]
- Diop, B.; Wang, D.R.; Drame, K.N.; Gracen, V.; Tongoona, P.; Dzidzienyo, D.; Nartey, E.; Greenberg, A.J.; Djiba, S.; Danquah, E.Y.; et al. Bridging old and new: Diversity and evaluation of high iron-associated stress response of rice cultivated in West Africa. J. Exp. Bot. 2020, 71, 4188–4200. [Google Scholar] [CrossRef] [PubMed]
- Melandri, G.; Sikirou, M.; Arbelaez, J.D.; Shittu, A.; Semwal, V.K.; Konaté, K.A.; Maji, A.T.; Ngaujah, S.A.; Akintayo, I.; Govindaraj, V.; et al. Multiple small-effect alleles of indica origin enhance high iron-associated stress tolerance in rice under field conditions in West Africa. Front. Plant Sci. 2020, 11, 604938. [Google Scholar] [CrossRef] [PubMed]
- Kaewcheenchai, R.; Vejchasarn, P.; Hanada, K.; Shirai, K.; Jantasuriyarat, C.; Juntawong, P. Genome-wide association study of local Thai indica rice seedlings exposed to excessive iron. Plants 2021, 10, 798. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Hunt, L.A.; Sheng, Q.; Szlavnics, Z. Cultivar evaluation and mega-environment investigation based on the GGE biplot. Crop Sci. 2000, 40, 597–605. [Google Scholar] [CrossRef]
- Haruna, A.; Adu, G.B.; Buah, S.S.; Kanton, R.A.; Kudzo, A.I.; Seidu, A.M.; Kwadwo, O.-A.; Moral, M.T. Analysis of genotype by environment interaction for grain yield of intermediate maturing drought tolerant top-cross maize hybrids under rain-fed conditions. Cogent Food Agric. 2017, 3, 1333243. [Google Scholar] [CrossRef]
- Mafouasson, H.N.A.; Gracen, V.; Yeboah, M.; Ntsomboh-Ntsefong, G.; Tandzi, L.; Mutengwa, C. Genotype-by-environment interaction and yield stability of maize single cross hybrids developed from tropical inbred lines. Agronomy 2018, 8, 62. [Google Scholar] [CrossRef]
- Carcedo, A.J.P.; Pardo, P.A.; Gambin, B.L. Secondary traits explaining sorghum genotype by environment interactions for grain yield. Crop Pasture Sci. 2017, 68, 599–608. [Google Scholar] [CrossRef]
- Fotokian, M.H.; Agahi, K. Biplot analysis of genotype by environment for cooking quality in hybrid rice: A tool for line × tester data. Rice Sci. 2014, 21, 282–287. [Google Scholar] [CrossRef]
- Balestre, M.; Borges, V.; Soares, A.A.; Reis, M. Stability and adaptability of upland rice genotypes. Crop Breed. Appl. Biotechnol. 2010, 10, 357–363. [Google Scholar] [CrossRef]
- Hussain, T.; Anothai, J.; Nualsri, C.; Soonsuwon, W. Evaluating performance of sixteen upland rice genotypes under field conditions for further breeding process. J. Agric. Sci. 2018, 10, 144. [Google Scholar] [CrossRef]
- Gauch, H.G. A simple protocol for AMMI analysis of yield trials. Crop Sci. 2013, 53, 1860–1869. [Google Scholar] [CrossRef]
- Yan, W.; Kang, M.S.; Ma, B.; Woods, S.; Cornelius, P.L. GGE biplot vs. AMMI analysis of genotype-by-environment data. Crop Sci. 2007, 47, 643–653. [Google Scholar] [CrossRef]
- Acuña, T.L.B.; Lafitte, H.R.; Wade, L.J. Genotype × environment interactions for grain yield of upland rice backcross lines in diverse hydrological environments. Field Crops Res. 2008, 108, 117–125. [Google Scholar] [CrossRef]
- Inabangan-Asilo, M.A.; Mallikarjuna Swamy, B.P.; Amparado, A.F.; Descalsota-Empleo, G.I.L.; Arocena, E.C.; Reinke, R. Stability and G × E analysis of zinc-biofortified rice genotypes evaluated in diverse environments. Euphytica 2019, 215, 61. [Google Scholar] [CrossRef]
- Nassir, A.L. Genotype × Environment analysis of some yield components of upland rice (Oryza sativa L.) under two ecologies in Nigeria. Int. J. Plant Breed. Genet. 2013, 7, 105–114. [Google Scholar] [CrossRef]
- Ferrari, S.; Pagliari, P.; Trettel, J. Optimum sowing date and genotype testing for upland rice production in Brazil. Sci. Rep. 2018, 8, 8227. [Google Scholar] [CrossRef]
- Pushpam, R.; Manonmani, S.; Vishnu Varthini, N.; Robin, S. Studies on yield, root characters related to drought tolerance and their association in upland rice genotypes. Electron. J. Plant Breed. 2018, 9, 856. [Google Scholar] [CrossRef]
- Nwilene, F.E.; Nacro, S.; Tamò, M.; Menozzi, P.; Heinrichs, E.A.; Hamadoun, A.; Dakouo, D.; Adda, C.; Togola, A. Managing insect pests of rice in Africa. In Realizing Africa’s Rice Promise; CAB International: Oxfordshire, UK, 2013; pp. 229–240. [Google Scholar]
- Abah, J.; Umar, A.; Bashir, M.; Drame, K.N.; Manneh, B.; Abo, M.E.; Sie, M. Soil characteristics of three iron toxic sites in Nigeria. J. Sci. Multidiscip. Res. 2012, 4, 68–72. [Google Scholar]
- Sikirou, M.; Shittu, A.; Konaté, K.A.; Maji, A.T.; Ngaujah, A.S.; Sanni, K.A.; Ogunbayo, S.A.; Akintayo, I.; Saito, K.; Ahanchédé, A.; et al. Screening African rice (Oryza glaberrima) for tolerance to abiotic stress: I. Fe toxicity. Field Crops Res. 2018, 220, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Drame, K.N.; Saito, K.; Koné, B.; Chabi, A.; Dakouo, D.; Annan-Afful, E.; Monh, S.; Abo, E.; Sié, M. Coping with iron toxicity in the lowlands of sub-Saharan Africa: Experience from Africa Rice Center. In Proceedings of the second Africa Rice Congress, Bamako, Mali, 22–26 March 2010. [Google Scholar]
- Kwanchai, A.G. Techniques for Field Experiments with Rice; Experimental Agriculture; International Rice Research Institute: Manila, Philippines, 1972; p. 40. [Google Scholar]
- IRRI. Standard Evaluation System (SES) for Rice; International Rice Research Institute: Manilia, Philippines, 2002; p. 52. [Google Scholar]
- Reyniers, F.N.; Truong, B.; Jacquinot, L.; Nicou, R. Breeding for drought resistance in dryland rice. In Drought Resistance in Crops with Emphasis on Rice; International Rice Research Institute: Los Banõs, Philippines, 1982; pp. 273–292. [Google Scholar]
- Rajonandraina, T.; Rakotoson, T.; Wissuwa, M.; Ueda, Y.; Razafimbelo, T.; Andriamananjara, A.; Kirk, G.J.D. Mechanisms of genotypic differences in tolerance of iron toxicity in field-grown rice. Field Crops Res. 2023, 298, 108953. [Google Scholar] [CrossRef]
- Moukoumbi, Y.D.; Bayendi Loudit, S.M.; Sikirou, M.; Mboj, D.; Hussain, T.; Bocco, R.; Manneh, B. Evaluation of Genotypic Variability and Analysis of Yield and Its Components in Irrigated Rice to Stabilize Yields in the Senegal River Valley Affected by Climate Change. Agronomy 2023, 13, 2218. [Google Scholar] [CrossRef]
- Hussain, T.; Hussain, N.; Tahir, M.; Raina, A.; Ikram, S.; Maqbool, S.; Fraz Ali, M.; Duangpan, S. Impacts of Drought Stress on Water Use Efficiency and Grain Productivity of Rice and Utilization of Genotypic Variability to Combat Climate Change. Agronomy 2022, 12, 2518. [Google Scholar] [CrossRef]
- Zeng, L.; Shannon, M.; Grieve, C. Evaluation of salt tolerance in rice genotypes by multiple agronomic parameters. Euphytica 2002, 127, 235–245. [Google Scholar] [CrossRef]
- Theerawitaya, C.; Wanchana, S.; Ruanjaichon, V.; Tisaram, R.; Samphumphuang, T.; Sotesaritkul, T.; Cha-Um, S.; Toojinda, T. Determination of traits responding to iron toxicity stress at different stages and genome-wide association analysis for iron toxicity tolerance in rice (Oryza sativa L.). Front. Plant Sci. 2022, 13, 994560. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- SAhmed, F.; Ullah, H.; Aung, M.Z.; Tisarum, R.; Cha-Um, S.; Datta, A. Iron Toxicity Tolerance of Rice Genotypes in Relation to Growth, Yield and Physiochemical Characters. Rice Sci. 2023, 30, 321–334. [Google Scholar] [CrossRef]
- Fageria, N.K.; Rabelo, N.A. Tolerance of rice cultivars to iron toxicity. J. Plant Nutr. 1987, 10, 653–661. [Google Scholar] [CrossRef]
- Zipporah, P.; Tokpah David, P.; Drame Khady, N.; Zogbo, L.; Voor Victor, M.; King Charles, F. Morphological variation of iron toxicity tolerance in lowland rice (Oryza sativa L.) varieties. World J. Adv. Res. Rev. 2022, 13, 038–046. [Google Scholar] [CrossRef]
- Arthaud, F. Formes d’adaptation des plantes aux milieux aquatiques. In L’eau à Découvert; CNRS Éditions: Paris, France, 2015; pp. 118–119. [Google Scholar]
- Li, R.; Li, M.; Ashraf, U.; Liu, S.; Zhang, J. Exploring the Relationships between Yield and Yield-Related Traits for Rice Varieties Released in China from 1978 to 2017. Front. Plant Sci. 2019, 10, 543. [Google Scholar] [CrossRef] [PubMed]
- Moukoumbi, Y.D.; Sié, M.; Vodouhe, R.; N’dri, B.; Toulou, B.; Ogunbayo, S.A.; Ahanchede, A. Assessing phenotypic diversity of interspecific rice varieties using agro-morphological characterization. J. Plant Breed. Crop Sci. 2011, 3, 74–86. [Google Scholar]
- Bocco, R.; Lorieux, M.; Seck, P.A.; Futakuchi, K.; Manneh, B.; Baimey, H.; Ndjiondjop, M.N. Agro-morphological characterization of a population of introgression lines derived from crosses between IR 64 (Oryza sativa indica) and TOG 5681 (Oryza glaberrima) for drought tolerance. Plant Sci. 2012, 183, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, K.; Vera Cruz, C.M.; Gruissem, W.; Bhullar, N.K. Large scale germplasm screening for identification of novel rice blast resistance sources. Front. Plant Sci. 2014, 5, 505. [Google Scholar] [CrossRef] [PubMed]
- Sultana, A.; Islam, M.M.; Sultana, A.; Khanom, M.S.R. Screening of rice genotypes for submergence stress at seedling stage. J. Agrofor. Environ. 2019, 13, 13–16. [Google Scholar]
- Makkar, G.S.; Bentur, J.S. Breeding for Stem Borer and Gall Midge Resistance in Rice. In Breeding Insect Resistant Crops for Sustainable Agriculture; Arora, R., Sandhu, S., Eds.; Springer: Singapore, 2017. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Mean Sq | Df | Pr (>F) |
|---|---|---|---|
| Days to heading | 153 | 1 | 0.001 *** |
| Plant height | 967 | 1 | 0.001 *** |
| Number of tillers per plant | 811 | 1 | 0.001 *** |
| Number of panicles per plant | 702 | 1 | 0.001 *** |
| Panicle length | 350 | 1 | 0.001 *** |
| Grain yield | 835 | 1 | 0.001 *** |
| Traits | Control Treatment (Ibadan) (N = 680) | Site with High Iron Content (Edozighi) (N = 680) | Overall (N = 1360) |
|---|---|---|---|
| Days to heading | |||
| Mean (SD) | 85.7 (8.79) | 91.8 (9.85) | 88.7 (9.82) |
| Median [Min; Max] | 87.0 [62.0; 110] | 94.0 [69.0; 116] | 90.0 [62.0; 116] |
| Plant height | |||
| Mean (SD) | 160 (17.5) | 108 (12.6) | 134 (30.5) |
| Median [Min; Max] | 163 [102; 201] | 108 [58.3; 148] | 127 [58.3; 201] |
| Number of tillers | |||
| Mean (SD) | 11.0 (2.97) | 19.5 (4.82) | 15.2 (5.85) |
| Median [Min; Max] | 10.7 [5.00; 23.0] | 18.8 [10.0; 36.3] | 14.3 [5.00; 36.3] |
| Number of panicles | |||
| Mean (SD) | 9.47 (2.80) | 16.5 (4.69) | 13.0 (5.21) |
| Median [Min; Max] | 9.00 [3.33; 26.7] | 15.7 [6.67; 34.3] | 12.0 [3.33; 34.3] |
| Panicle length | |||
| Mean (SD) | 28.8 (3.17) | 25.2 (2.99) | 27.0 (3.57) |
| Median [Min; Max] | 29.0 [20.0; 41.0] | 25.0 [18.3; 32.0] | 27.0 [18.3; 41.0] |
| Grain yield | |||
| Mean (SD) | 6120 (2520) | 1390 (665) | 3750 (3000) |
| Median [Min; Max] | 6230 [283; 11,700] | 1340 [0; 8230] | 2240 [0; 11,700] |
| Genotypes | Edozighi GY | Ibadan GY | %Losses |
|---|---|---|---|
| IR 88638-325-1-1-1-1-1-1 | 956 | 1015 | 6 |
| IR 88638-102-1-1-1-1-1-1 | 640 | 685 | 7 |
| IR 88638-34-1-1-1-1-1-1 | 1608 | 1732 | 7 |
| IR 88638-98-1-1-1-1-1-1 | 1904 | 2718 | 30 |
| IR 88638-240-1-1-1-1-1-1 | 1187 | 1765 | 33 |
| IR 88638-13-1-1-1-1-1-1 | 1430 | 2352 | 39 |
| IR 88638-230-1-1-1-1-1-1 | 2855 | 4793 | 40 |
| IR 88638-308-1-1-1-1-1-1 | 1537 | 2765 | 44 |
| IR 88638-39-1-1-1-1-1-1 | 2803 | 5223 | 46 |
| IR 88638-208-1-1-1-1-1-1 | 1299 | 2431 | 47 |
| Suakoko 8 (parent line) | 987 | 2467 | 60 |
| BW348-1 | 1295 | 4035 | 68 |
| WITA4 | 1277 | 4420 | 71 |
| Bao Thai (parent line) | 915 | 4083 | 78 |
| IR64 | 620 | 3439 | 82 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sikirou, M.; Shittu, A.; Moukoumbi, Y.D.; Arouna, A.H.; Zokpon, C.; Bocco, R.; Najimu, A.; Ramaiah, V. Field Evaluation of Rice Lines Derived from Suakoko 8 X Bao Thai for Iron Tolerance in the South Saharan African Farming System. Plants 2024, 13, 1610. https://doi.org/10.3390/plants13121610
Sikirou M, Shittu A, Moukoumbi YD, Arouna AH, Zokpon C, Bocco R, Najimu A, Ramaiah V. Field Evaluation of Rice Lines Derived from Suakoko 8 X Bao Thai for Iron Tolerance in the South Saharan African Farming System. Plants. 2024; 13(12):1610. https://doi.org/10.3390/plants13121610
Chicago/Turabian StyleSikirou, Mouritala, Afeez Shittu, Yonnelle Dea Moukoumbi, Aboudou Hack Arouna, Chédrac Zokpon, Roland Bocco, Adetoro Najimu, and Venuprasad Ramaiah. 2024. "Field Evaluation of Rice Lines Derived from Suakoko 8 X Bao Thai for Iron Tolerance in the South Saharan African Farming System" Plants 13, no. 12: 1610. https://doi.org/10.3390/plants13121610