Abstract
This study presents the phytochemical composition, antioxidant (hydrogen atom and single-atom transfer mechanisms), and digestive enzyme inhibitory (alpha-amylase, alpha-glucosidase, and pancreatic lipase) activities of ethanol-based extractions and traditional preparations (infusion) of the leaves of wild-edible Allium species (A. kharputense, A. affine, A. shirnakiense, and A. akaka) from the highlands of Eastern Anatolia. Among the eight extracts analyzed, ethanol extractions of the A. kharputense and A. akaka leaves exhibited better biotherapeutic activities and had the highest bioactive content. The dominant bioactive profile was composed of mainly allicin and phenolic compounds (chlorogenic acid, hesperidin, rutin, isoquercitrin, and quercetin) with small amounts of fatty acids. These data were similar to the biological activities and chemical composition of common Allium species and suggest the utilization of the extracts of wild-edible Allium species in the development of Allium-based biotherapeutics or nutraceuticals.
1. Introduction
Plant-based natural materials are extensively used as food in Eastern Anatolia, which represents a rich traditional knowledge of food plants due to its rich biodiversity, and local flora and fauna. Also, their herbal infusions have been used as local medicines to treat various ailments, such as inflammation, diabetes, stomachache, hemorrhoids, colds, and cancer [1]. Of them, a high number of Allium species have been used as food and medicine due to their common vegetation potential in the wild. Ethnobotanical studies revealed the extensive uses of leaves and bulbs of Allium species as food and medicine [2,3], and their therapeutic and nutritive potentials were later reported [4,5,6].
It has been reported that more than 600 Allium species grow in the wild [7] and only a limited number of these species, including onion (Allium cepa) and garlic (Allium sativum), has been cultivated and consumed by several ethnic groups across the world [3], which contain significant nutraceuticals and phytochemicals useful for the treatment or management of metabolic disorders [8], cancer, and cardiovascular diseases, and exhibit numerous biological activities, including antimicrobial, antiviral, antioxidant, antispasmodic, antimutagenic, neuroprotective, and immunomodulatory activities [9,10]. However, it was reported that local Allium species showed comparable biological activities to cultivated Allium species, including onion and garlic [11].
Due to its tribal and semi-nomadic lifestyle, local foods have represented a high scale of plant foods in Eastern Anatolia until now. Allium species are not only significant flavoring agents and food bioresources, but also significant sources of medicinal practices in Eastern Anatolia. In addition to commonly cultivated Allium species (onion and garlic), a great number of Allium species have been used in the preparation of food (herbal cheese, omelets, sauces etc.) and medicine (particularly diabetes) in the region. Of them, A. kharputense, A. affine, A. shirnakiense, and A. akaka are commonly collected from the highlands and stored as dried for later use [2,12]. The bulbs and leaves of these species are generally utilized as an infusion for medicine or eaten raw for food. Though they have extensive utilization, scientific reports regarding their biological activities and chemical composition are limited.
Plants can produce many diverse bioactive compounds, such as phenolics, carotenoids, and tocopherols, which have medicinal properties and nutritional value. High concentrations of phytochemicals accumulate in plant organs, which act as natural antioxidants and enzyme inhibitors. To extract phytochemicals from the plant matrix, various extraction methods, including solvent-based, microwave-assisted, enzyme hydrolysis, supercritical fluids, and ultrasonic-assisted methods, are applied. Of them, the solvent-based extraction method has been extensively utilized because of its effectiveness, ease of use, and wide applicability. One of the most important advantages of the solvent-based extraction method is the polarity index of the solvent used, which provides to the extract targeted phytochemicals from the plant matrix [13,14,15]. Therefore, we hypothesized that traditional preparations (infusion) and ethanol-based extracts of these species must be analytically analyzed to understand their chemical nature, as well as reveal the beneficiation percentage in the diet for human health and determine the potential of local Allium species as functional foods for the management of enzyme inhibition-related metabolic diseases.
2. Results
2.1. In Vitro Biotherapeutic Potential
2.1.1. Extraction Yields
The extraction yields of the extracts obtained from the leaves of Allium species are presented in Table 1. Among all the extracts analyzed, the A. affine and A. akaka plant materials gave the highest yields through the infusion method both for leaves. The results showed that the use of water and the application of heat treatment showed a better extraction of phytochemicals from the plant matrix.

Table 1.
Yields, total phenolic contents, and antioxidant activities.
2.1.2. Total Phenolic Contents and Antioxidant Activities
Total phenolic contents and antioxidant activities through the single-electron transfer mechanism, which was represented by the FRAP assay, showed that the ethanol extract of Allium akaka leaf sample (48 mg gallic acid E and 872 μmol Fe2+/g extract, respectively) had the highest results. With regard to the ORAC assay, which represented the hydrogen atom transfer mechanism, ethanol extracts of the Allium akaka and Allium kharputense leaf samples showed better antioxidant activities. Also, the ethanol solvent had a higher capacity for extracting antioxidant compounds than the water solvent (Table 1). These findings are comparable with several Allium species, such as Allium cepa [16], Allium roseum [17], and Allium scorodoprasum [18].
2.1.3. Enzyme Inhibition Activities
The extracts showed moderate inhibitory activities against alpha-glucosidase and pancreatic lipase, but not amylase. However, the extracts were found as weaker inhibitory agents according to the positive controls: acarbose and orlistat. Like antioxidant capacities, the ethanol extracts were stronger than infusion preparations (Table 2). Among all samples tested, Allium akaka showed the highest inhibitory activities (IC50: 500 and 195 μg/mL for α-glucosidase and pancreatic lipase, respectively). The inhibitory activities of the extracts against α-glucosidase were high and against α-amylase were low, which was like the inhibitory activities of Allium cepa extracts [16]. In agreement, Nickavar and Yousefian [19] reported that Allium species are weak inhibitors of α-amylase.
Table 2.
Enzyme inhibitory activities.
2.1.4. Phytochemical Composition
The phytochemical compounds identified using HPLC-PDA-MS/MS are shown in Table 3. Six dominant compounds were identified as described below. The first compound was identified as chlorogenic acid according to a maximum absorption at 326 nm and a charged molecular ion of 353 m/z in the negative ion mode with the detection of a fragment ion of 191 m/z in the SRM mode. This compound was detected at very high levels in all samples, excluding A. shirnakiense.
Table 3.
Phytochemical composition using HPLC-MS/MS.
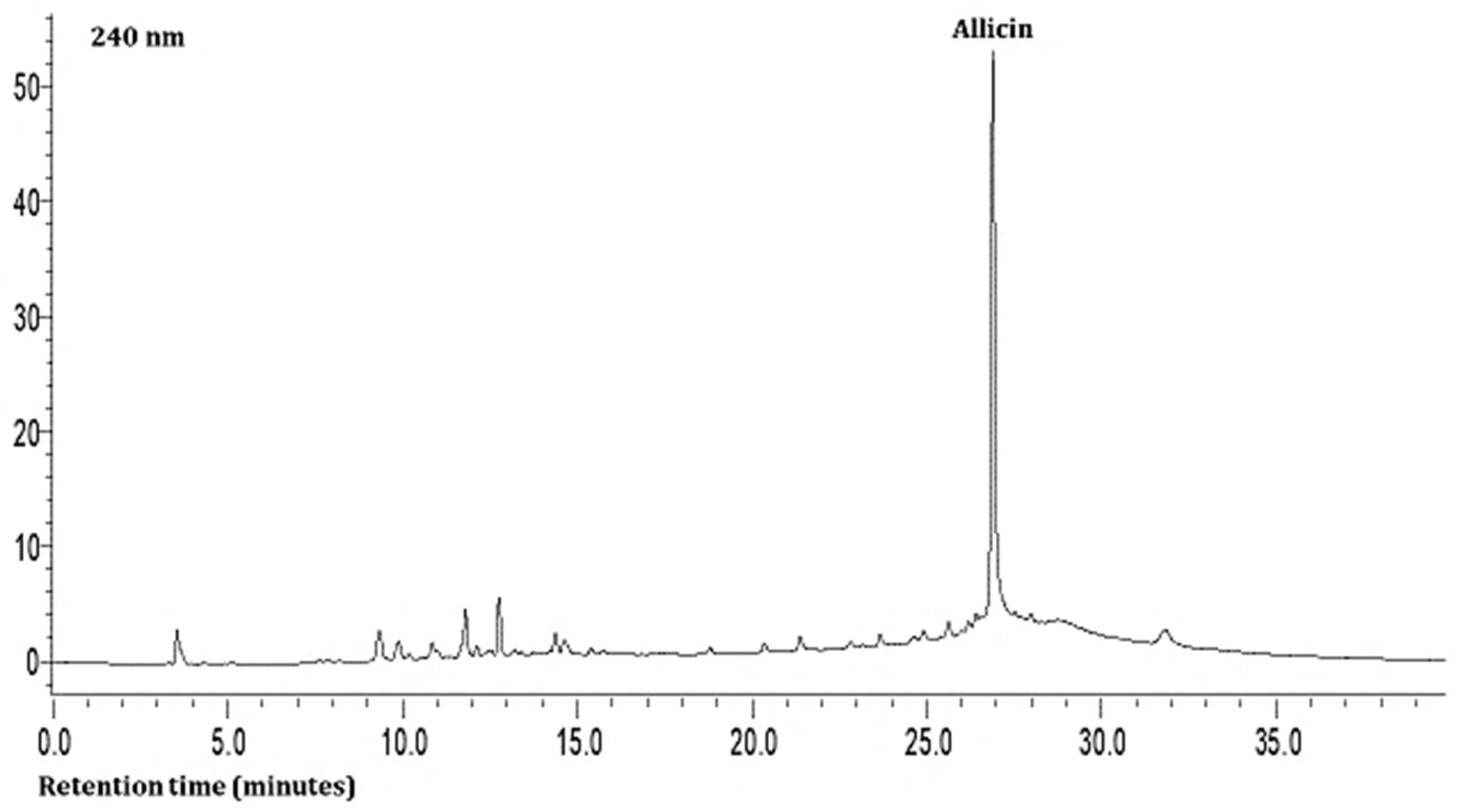
The second compound was characterized as hesperidin based on a charged molecular ion of 609 m/z in the negative ion mode with the detection of fragment ions of 449 and 431 m/z in the SRM mode. This was found in all the leaf samples at low levels (Table 3). The third, fourth, and fifth compounds present in the extracts were quercetin and its glycosides (rutin and isoquercitrin) at very high levels based on the MS data. The seventh compound was identified as allicin based on a charged molecular ion of 163 m/z in the positive ion mode with the detection of a fragment ion of 121 m/z in the SRM mode (Table 3; Figure 1). This compound was found in all the samples at high levels.
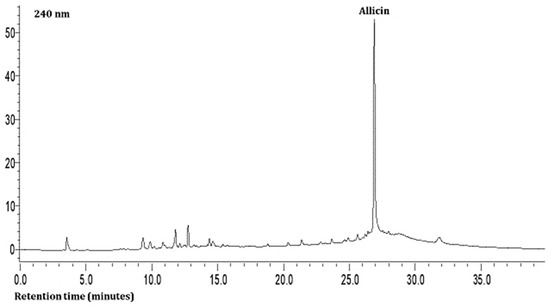
Figure 1.
Representative HPLC chromatogram of Allium leaves at 240 nm.
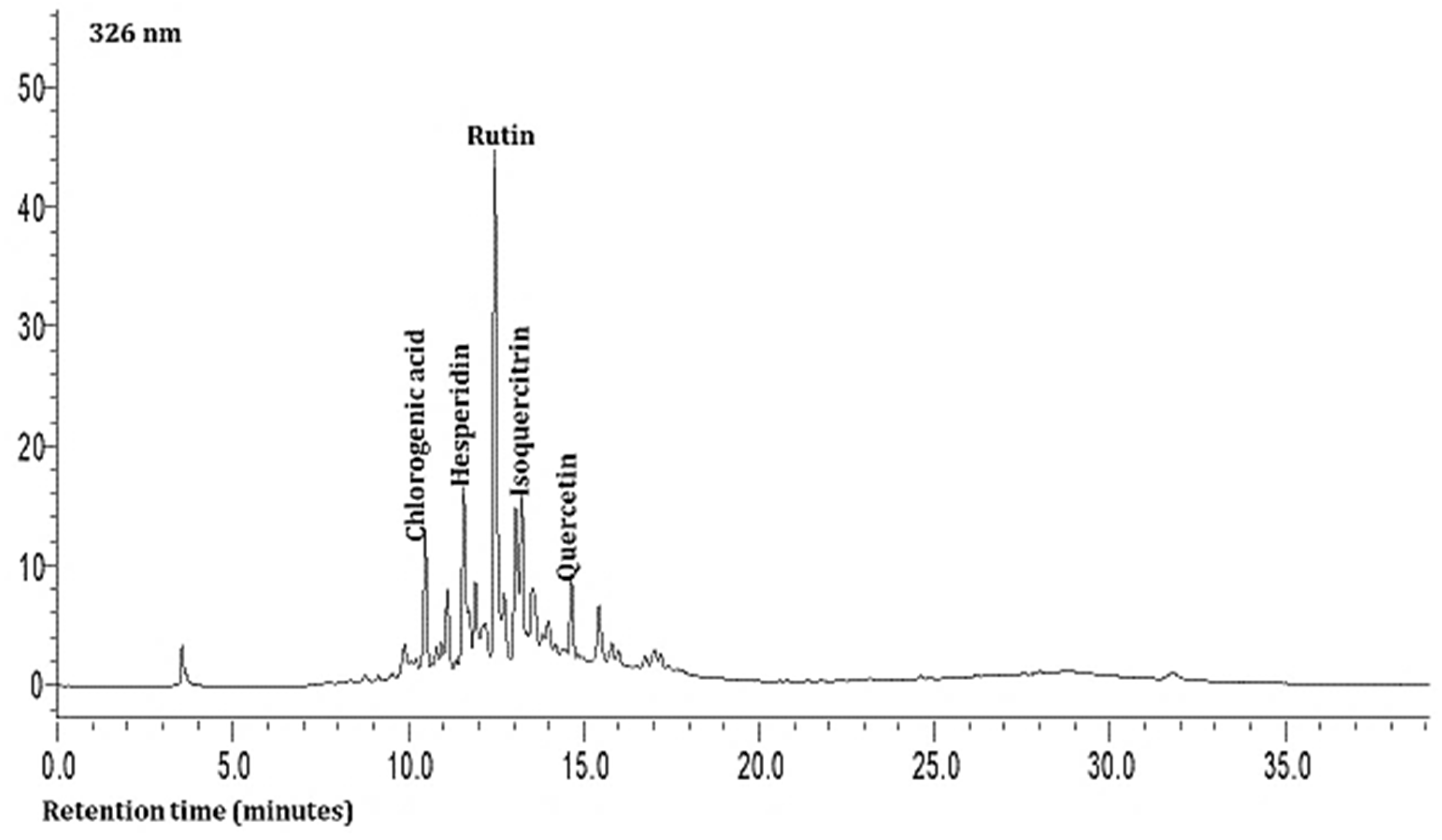
As shown in Table 3 and Figure 1 and Figure 2, the hydrophilic composition of the infusions and ethanol-based extracts mainly consisted of allicin (Figure 1), an organosulphur compound, and various phenolic compounds, including chlorogenic acid, rutin, isoquercetin, and quercetin (Figure 2). It was found that the Allium akaka and Allium kharputense samples suggested rich sources of phenolics. Most phenolic compounds consisted of flavonols with the domination of quercetin and its derivatives. The representative compound of phenolic acid was found as chlorogenic acid.
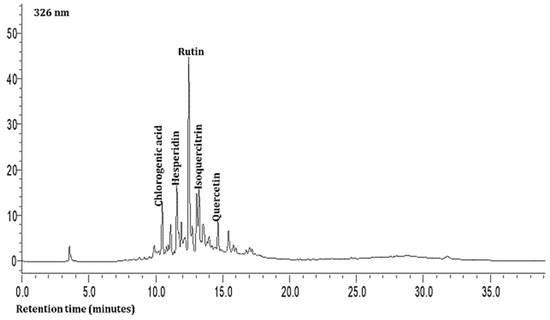
Figure 2.
Representative HPLC chromatogram of Allium leaves at 326 nm.
On the other hand, fatty acids (palmitic, stearic, linoleic, and linolenic acids) dominated the lipophilic composition of the extracts (Table 4). The major fatty acids were found to be oleic, palmitic, and linolenic acids.
Table 4.
Fatty acid composition using GC-MS.
3. Discussion
The solubility of phytochemicals is governed by the chemical nature of the plant sample, as well as the polarity of the solvents used. The choice of the solvent in the extraction procedure affects the recovery of phytochemicals from the plant matrix. For example, a low polarity-indexed solvent, such as n-hexane or chloroform, can extract fatty acids, waxes, and terpenoids, while a medium polarity-indexed acetone can extract less polar and polar flavonoids, tannins, terpenoids, and glycosides, based on the chemical nature of the plant material extracted [13]. The chromatographic studies revealed that those solvents (water and ethanol) used in this study extracted the same compounds at different concentrations. The ethanol solvent had a higher capability of extracting hesperidin, rutin, isoquercitrin, quercetin, allicin, and chlorogenic acid. Also, it can be suggested that the heating procedure used in the infusion method had no remarkable effect on the extraction of phytochemicals from the plant matrix (Table 3). Kim et al. [16] reported that using ethanol is more convenient to recover phytochemicals, particularly quercetin, from onion skin. Scientific reports showed that the major phytochemicals present in Allium species differ according to the plant material studied, such as quercetin in Allium cepa [16]; quercetin, luteolin, and eriodictyol in Allium scorodoprasum [18]; kaempferol and its glycosides in Allium tuberosum [20] and Allium roseum [17]; allicin and isoquercitrin in Allium schoenoprasum; rutin in Allium senescens [21]; rutin, quercetin, and kaempferol in Allium flavum [22]; and quercetin 3,4′-diglucoside, quercetin 4′-monoglucoside, quercetin aglycone, isorhamnetin, and kaempferol in Allium cornutum [10]. Moreover, Schmidt et al. [11] investigated the phytochemical composition of 35 Allium species and found quercetin to be a representative phenolic compound of the extracts, which agrees with our findings. Like our findings, allicin was found in 33 Allium accessions representing 14 species, which makes it a taxonomic key compound [23]. Rutin, chlorogenic acid, isoquercitrin, quercetin, and allicin were found to be major phytochemicals of Allium species in this study, which agrees with previous studies conducted in common and local Allium species [18,21,22,24]. Also, the presence of quercetin and its glycosides, which are among the most common phenolics of wild plants, showed the adaptation abilities of Allium species in the wild.
One of the most effective therapeutic approaches to manage postprandial hyperglycemia is to limit the activity of the carbohydrate digestive enzymes involved in the release of glucose from the daily diet. Alpha-amylase and alpha-glucosidase are among the key enzymes in the digestive process that are responsible for converting polysaccharides to disaccharides and monosaccharides, which results in the increasing of the blood glucose level. α-Amylase breaks down large polysaccharides into sugars in the saliva. It is the key enzyme that degrades the polymeric substrate into shorter oligomers by catalyzing the hydrolysis of the α-1,4-glucan linkages present in starch, maltodextrins, and other related carbohydrates [25]. α-Glucosidase is an enzyme that catalyzes the final step of glucose absorption in the intestine during the digestive process of carbohydrates, hence α-glucosidase inhibitors could retard the rapid hydrolysis of dietary carbohydrates and suppress postprandial hyperglycemia. Dietary starch and other related carbohydrates are digested by α-amylase to a large quantity of maltose, which is further digested by α-glucosidase to glucose to be absorbed in the human intestine [26]. Natural α-amylase and α-glucosidase inhibitors from plant sources offer an attractive strategy to control postprandial hyperglycemia. Especially these, which have a lower inhibitory activity against α-amylase and a stronger inhibitory activity against α-glucosidase, can be used as an effective therapy for postprandial hyperglycemia with minimal side effects [27]. This is because the simultaneous inhibition of both enzymes would result in abnormal bacterial fermentation in the colon due to the presence of undigested carbohydrates, which cause unwanted side effects such as abdominal distention, flatulence, meteorism, and possibly diarrhea [28]. Among the Allium species evaluated in this study, A. akaka and A. kharputense fulfill the condition of possessing a high α-glucosidase inhibitory activity with a low inhibition of α-amylase, suggesting a desirable feature for candidates of nutraceuticals and biotherapeutics. Besides, the extracts showed moderate levels of the pancreatic lipase enzyme, which reduces the formation of lipids in the body and provides a desirable feature to the plant-based foods or nutraceuticals [29].
The scientific reports revealed that allicin- and phenolic-rich Allium species were weak alpha-amylase [19] inhibitors, but strong alpha-glucosidase [11] inhibitors. Also, it was found that rutin is an effective agent of antilipase activity [27,28,29,30]. Kim et al. [16] and Schmidt et al. [11] found a positive correlation between the level of quercetin and the antioxidant activity and α-glucosidase activity in several extracts obtained from Allium cepa. Nishimura et al. [31] found that consumption of quercetin-rich onion had beneficial effects in preventing obesity and improving liver function through low HDL-C and ALT levels in vivo. Compatibly, there is a positive correlation between the levels of quercetin and its glycosides and biological activities in this study, which suggests that quercetin and its glycosides are among the major contributors of the antilipase and antioxidant activities. Kamalakkanan and Price [32] reported that the oral administration of rutin to diabetic rats significantly decreased fasting plasma glucose and glycosylated hemoglobin, and increased insulin, C-peptide, hemoglobin, thiobarbituric acid reactive substances, and lipid hydroperoxide levels, and increased the non-enzymic antioxidants significantly. The authors also found that the treatment of normal rats with rutin did not significantly alter any of the parameters studied. These findings showed that rutin, a glycoside of quercetin, showed a high antihyperglycemic and antioxidant activity in diabetic rats, but not in healthy ones, which suggests a relationship between oxidative stress and metabolic diseases and the targeted therapeutic effect of the rutin compound. The presence of chlorogenic acid [33,34], rutin [34,35], and allicin [36] in plant extracts provided effective antioxidant abilities, which suggests they are major contributors of antioxidant abilities in Allium species grown in Eastern Anatolia. The lipophilic compounds present in the extracts were fatty acids, which were reported as weak antioxidant agents [37], indicating their limited contribution to antioxidant activity.
Chlorogenic acid is a phenolic compound from the hydroxycinnamic acid family, which possesses many biological activities, and it was reported that chlorogenic acid-rich foods had preventive effects such as the suppression of an increase in body fat and sugar cataracts, and a reduction in glucose absorption and therapeutic effects, including improved fasting glucose, reduction in weight, acceleration of diabetic wound healing, and improved symptoms of diseases over metabolic syndromes [38]. Allicin, which is considered a specific representative compound of Allium species [39], possesses broad antimicrobial and health-enhancing properties, which ensure they are natural and safe agents against synthetic preservatives and enzyme inhibitory compounds [23].
Due to a higher affinity to specific amino acids of the enzyme’s protein, phenolic compounds can suppress the activity of the enzymes related to metabolic diseases [40]. The phytochemicals present in the extracts were among the most effective enzyme inhibitors. For example, quercetin bound to Asp80, Arg257, Val260, and Ala261 amino acids in pancreatic lipase. The size and structural rigidity of quercetin could be responsible for its binding characteristics (more polar interactions) and, consequently, its higher pancreatic lipase inhibitory potency [41].
The antioxidant and enzyme inhibitory activities of quercetin and its glycosides are mainly based on their structural features, which can be summarized as the presence of (i) a catechol structure in the B ring, (ii) a double bond between C2 and C3 in the C ring, and (iii) the number of aromatic and hydroxyl groups. According to the abovementioned criteria, quercetin is one of the most effective antioxidant and enzyme inhibitory compounds among the flavonoids [41], which explains the positive correlation between biological activities and the levels of quercetin and its glycosides (rutin and isoquercitrin) in Allium species.
The Allium species analyzed within this study had high antioxidant and moderate enzyme inhibitory (alpha-glucosidase and pancreatic lipase) activities and contained phenolic compounds (chlorogenic acid, hesperidin, rutin, isoquercitrin, and quercetin) and allicin as major chemical compounds. These data were similar to previous reports regarding the chemical composition and biological activities of common Allium species and can contribute to the scientific database of Allium species.
4. Materials and Methods
4.1. Plant Materials
The leaves of four wild-edible Allium species (Table 5) were collected from the highlands of Eastern Anatolia, Turkey on 12 May 2018. The scientific identification was performed at the Van Pharmaceutical Herbarium (VPH), Pharmacy Faculty, Van Yuzuncu Yil University (Van, Turkey). The plant materials were air-dried in the dark at room temperature (22 ± 2 °C) for 96 h. Then, they were ground into a fine powder using a grinding mill (Isolab laboratory mill 602, Interlab, İstanbul, Turkey) and stored at −20 °C for a maximum of 4 weeks until analyzed.
Table 5.
Botanical and ethnobotanical knowledge of wild-edible Allium taxa from the highlands of Eastern Anatolia.
4.2. Chemicals
Unless otherwise stated, all chemicals were purchased from Sigma-Aldrich, Inc. (St. Louis, MO, USA) and were of analytical or HPLC grade.
4.3. Preparation of Samples for Analysis
4.3.1. Ethanol Extract
The ethanol extracts were prepared as described previously [42]. Briefly, fine powders of the plant materials were mixed with a 10-fold volume of ethanol (80%, obtained from 95%), and shaken for 2 h at room temperature at 15,300× g (10,000 rpm in a Beckman JA14 (137 mm) rotor, Sorvall RC-5B centrifuge, Wilmington, DE, USA) at 4 °C for 20 min with the supernatant collected. Subsequently, the supernatants were evaporated under a reduced pressure at 37 °C using a rotary evaporator (Rotavapor R-205, Buchi, Flawil, Switzerland) and then freeze-dried using a lyophilizator (Alpha 1-2 LDplus, Christ, Osterode am Harz, Germany) under a vacuum at −51 °C to fine lyophilized powders. The lyophilized extracts were stored at −20 °C for a maximum of 4 week until analyzed.
4.3.2. Herbal Infusion Extract
The lyophilized herbal infusions were prepared according to Baytop [43]. Briefly, the plant materials were mixed with a 10-fold volume (g/mL) of pre-boiled distilled water and incubated for 10 min. Then, the mixtures were filtered using cotton and vacuum filtering. The filtrates were evaporated and freeze-dried as described previously.
4.4. Antioxidant Capacity
4.4.1. Folin-Ciocalteu Reducing (FCR)
The FCR levels of the extracts were measured as described previously [44]. Briefly, 25 µL of the extracts were mixed with 125 µL Folin–Ciocalteu reagent in 96-well microplates and shaken for 3 min. The absorbance was measured at 600 nm for ascorbic acid correction. Subsequently, 125 µL of 6% Na2CO3 were added and the microplate was shaken for 12 min. The absorbance was then measured at 600 nm. The results were expressed as mg gallic acid (GA) E/g dw based on a GA standard curve. The analyses were conducted in triplicate.
4.4.2. Ferric Reducing Antioxidant Power (FRAP)
The FRAP values of the extracts were determined as described previously [44]. Briefly, 10 µL of the extracts were placed in 96-well microplate and 200 µL of the FRAP reagent was added, followed by 15 s of shaking. After incubation for 8 min, the absorbance was measured at 595 nm using a spectrophotometer and the reducing capacities of the extracts were expressed as mmol of iron (Fe2+)/g dw based on an iron sulphate standard (Fe2SO4) curve against a blank control. The analyses were performed in triplicate.
4.4.3. Oxygen Radical Absorbance Capacity (ORAC)
The ORAC assay was carried out as previously described [44]. Briefly, 20 µL of the extracts were added to a microplate with a 20 min incubation at 37 °C. Subsequently, 20 µL AAPH was added to each well and the fluorescence measurements initiated immediately. Fluorescence (excitation wavelength 495 nm and emission wavelength 515 nm) was measured every min until the fluorescence reached zero and a kinetic curve was obtained using the Mars data analysis software 5.0 (POLARstar Omega, BMG Labtech, Offenburg, Germany). The area under the curve (AUC) was integrated and standardized against a blank control (without AAPH). The measurements were carried out in triplicate. The ORAC levels of the samples were expressed as mmol Trolox equivalents (E)/g dw based on a Trolox standard curve.
4.5. Enzyme Inhibitory Activities
4.5.1. α-Glucosidase Inhibitory Activity
The inhibition of the α-glucosidase enzyme was determined as described previously [42]. Briefly, 20 µL of the sample solutions were mixed with 20 µL of 2% sucrose (w/v) in a maleate buffer. The enzymatic reaction was initiated by adding 20 µL of enzyme solution prepared in 1 mL of 100 mmol maleate buffer and the mixture was incubated at 37 °C for 60 min. The enzymatic reaction was terminated by heating the mixture at 100 °C for 10 min and the absorbance was measured at 505 nm. The relative α-glucosidase inhibition was calculated using the formula: % Inhibition = [((ACB − AC) − (ASB − AS))/(ACB − AC)] × 100, where AS and AC were the absorbance of the sample and the negative control, and where ASB and ACB were the absorbance of the sample blank and the control blank, respectively. Acarbose was used as a positive control.
4.5.2. α-Amylase Inhibitory Activity
The inhibition of α-amylase was performed using the Caraway–Somogyi iodine/potassium iodide (IKI) method as described previously [42]. The assay was conducted with α-amylase Type 1-A: DFP treated from porcine pancreas (Sigma Aldrich, Istanbul, Turkiye) and the Amylase-Test Wako kit (Wako Pure Chemical Institute, Tokyo, Japan).
A sample of starch (200 μL, 0.4 mg/mL) and an extract (100 μL, 20 mg/mL of lyophilized powder) in the phosphate buffer (20 mmol NaH2PO4 and 6.7 mmol NaCl, pH 6.9) were added to a clean cuvette at room temperature. Subsequently, 50 μL of pancreatic porcine α-amylase solution (1 U/1 mL in the phosphate buffer) was added to the sample mixture, after which the phosphate buffer was added to obtain a final volume of 500 μL. The mixture was incubated at room temperature for 3 min to allow the enzymatic reaction to occur. The reaction was terminated by adding hydrochloric acid (1 mL, 0.1%). Subsequently, an iodine reagent (Wako amylase kit) was added to the mixture. The decrease in starch concentration due to the activity of α-amylase was measured at 660 nm using a Shimadzu 1601 spectrophotometer (Tokyo, Japan). The enzyme inhibitory activities of the extracts were calculated as a percentage of the control using the following equation:
where ACB is the absorbance of the control blank, AC is the absorbance of the control, ASB is the absorbance of the sample blank, and AS is the absorbance of the sample. Acarbose was used as a positive control.
% Inhibition = (ACB − AC) − (ASB − AS)/(ACB − AC) × 100
4.5.3. Pancreatic Lipase Inhibitory Activity
The lipase inhibitory activity was assayed as described previously [42]. A total of 25 µL of sample solutions prepared using a citric/phosphate buffer (1 mL of 100 mmol), 50 µL of 4-Muo, and 25 µL of the enzyme solution were placed in a 96-well microplate, inserted into the microplate reader and incubated for 20 min at 37 °C. Subsequently, the reaction was terminated by adding 50 µL of 1 mol hydrochloric acid and 100 µL of sodium citrate, respectively. The relative lipase inhibition activity was calculated using the formula: % Inhibition = (1 − (FS − FSB)/(FC − FCB)) × 100, where FS and FC were the values of the samples and the negative control measured fluorometrically at an emission wavelength of 460 nm and an excitation of 320 nm with slit widths of 5 nm using the microplate reader, and where FSB and FCB were the fluorescence readings of the sample blank and the control blank, respectively. Orlistat was used as positive control.
4.6. Phytochemical Profile
4.6.1. Identification and Quantification of Phenolic Compounds
The identification and quantification of phenolic compounds using a high performance liquid chromatography-diode array detector (HPLC-DAD) and liquid chromatography-photo-diode array–mass spectrometry (LC-DAD–MS/MS) on a Quantum triple-stage quadrupole mass spectrometer, equipped with a quaternary solvent delivery system, a column oven, a photo-diode array detector, and an auto sampler (Thermo Fisher Scientific, Waltham, MA, USA), were performed as described previously [42]. An aliquot (3 μL) of each sample solution was chromatographed on a 150 × 2.1 mm, 5 μm Luna Synergy Hydro column (Phenomenex, Torrance, CA, USA), which was heated to 30 °C. The analytes were separated using 0.5% formic acid and acetonitrile (200 µL/min flow rate). The photodiode array detector was used to obtain data from 190–520 nm. The composition of phenolic compounds was identified based on their UV spectrum, retention time, co-chromatography with commercial standards, when available, and MS/MS fragmentation patterns. Mass spectrometry data were obtained using an electrospray source in either the positive (quercetin 3-glucoside) or the negative (chlorogenic acid) modes. MS experiments in the full scan (parent and product-specific) and the selected reaction monitoring (SRM) mode were performed. The quantification of phenolic compounds was performed using the HPLC-DAD system, which consisted of two LC-10ADVP pumps, an SPD-M10ADVP diode array detector, a CTO-1-ADVP column oven, a DGU-12A degasser, an SIL-10ADVP auto injector, and an SCL-10A system controller equipped with an Atlantis column (dC18, 4.6 × 100 mm, 5 µm particle size) (Waters Associates, Milford, MA, USA). Analytical HPLC was run at 30 °C and monitored at 250, 280, 320, 370, and 520 nm. The injection volume was 10 µL. The levels of the phenolic compounds were quantified as an authentic standard E/g dw based on a calibration curve.
4.6.2. Identification and Quantification of Fatty Acid (FA) Compounds
The FA compounds present in lyophilized extracts were measured using gas chromatography mass spectrometry (GC/MS) (3800 Varian GC, Agilent Technologies, Istanbul, Turkey) using a headspace solid-phase microextraction as described previously by Uzun et al. [45]. Briefly, the extracts were chromatographed on a 60 × 0.25 mm i.d., 0.2 μm HP 88 column (Agilent Technologies, Istanbul, Turkey). The oven temperature was programmed as follows: 80 °C at the start, followed by an increase to 190 °C at a rate of 5 °C/min, held at 190 °C for 3 min, followed by an increase at a rate of 5 °C/min to 260 °C, and held for 3 min. The subsequent temperature increase was at a rate of 2 °C/min to 275 °C and held for 12.5 min. The carrier gas was helium, used at a constant pressure of 10 psi; the transfer line temperature was 250 °C; electron impact (EI); acquisition ion range, 40 to 200 m/z; scan rate.
4.7. Data Analysis
A one-way ANOVA with the Bonferroni post hoc test was used to measure differences between the samples at p < 0.05 using Graphpad Prism 5 (Graphpad Software, San Diego, CA, USA). All the IC50 values were determined through the corresponding dose-inhibition curves according to their best-fit shapes.
5. Conclusions
This is the first report of the biological activities and phytochemical composition of Allium species, including A. akaka, A. affine, A. kharputense, and A. shirnakiense, grown and consumed as food in daily life in the highlands of Eastern Anatolia. Th phytochemical compounds detected within this study (allicin and various phenolics) were among the major contributors to the antioxidant and enzyme inhibitory activities, with the contribution of fatty acids. Catechol moiety-containing compounds rich in ethanol-based leaf extracts had a superior antioxidant capacity both for the HAT and SET mechanisms and the inhibition of of α-glucosidase and pancreatic lipase. Ethanol extracts and infusion preparations of Allium akaka and Allium kharputense were found to be potent sources of functional foods and/or biotherapeutics, which contain significant phytochemicals.
Author Contributions
Conceptualization, methodology, software, validation, formal analysis, investigation, resources, data curation, writing—original draft preparation, writing—review and editing, visualization, supervision, project administration, and funding acquisition, M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Acknowledgments
The author thanks Abdullah Dalar from Mardin Artuklu University for his supervision during the investigation and help in writing—original draft preparation.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Dalar, A.; Mukemre, M.; Unal, M.; Ozgokce, F. Traditional medicinal plants of Ağrı Province, Turkey. J. Ethnopharmacol. 2018, 226, 56–72. [Google Scholar] [CrossRef] [PubMed]
- Fırat, M. The Ethnobotanical Usage of Some East Anatolian (Turkey) Allium L. Species. Manas J. Agric. Vet. Life Sci. 2015, 5, 80–86. [Google Scholar]
- Ekşi, G.; Özkan, A.M.; Koyuncu, M. Garlic and onions: An eastern tale. J. Ethnopharmacol. 2020, 253, 112675. [Google Scholar] [CrossRef]
- Rahman, M.S. Allicin and Other Functional Active Components in Garlic: Health Benefits and Bioavailability. Int. J. Food Prop. 2007, 10, 245–268. [Google Scholar] [CrossRef]
- Corzo-Martínez, M.; Corzo, N.; Villamiel, M. Biological properties of onions and garlic. Trends Food Sci. Technol. 2007, 18, 609–625. [Google Scholar] [CrossRef]
- Santhosha, S.G.; Jamuna, P.; Prabhavathi, S. Bioactive components of garlic and their physiological role in health maintenance: A review. Food Biosci. 2013, 3, 59–74. [Google Scholar] [CrossRef]
- Poojary, M.M.; Putnik, P.; Kovačević, D.; Barba, F.; Lorenzo, J.; Dias, D.; Shpigelman, A. Stability and extraction of bioactive sulfur compounds from Allium genus processed by traditional and innovative technologies. J. Food Compos. Anal. 2017, 61, 28–39. [Google Scholar] [CrossRef]
- Lanzotti, V. The analysis of onion and garlic. J. Chromatogr. 2006, 1112, 3–22. [Google Scholar] [CrossRef]
- Putnik, P.; Gabrić, D.; Roohinejad, S.; Barba, F.; Granato, D.; Mallikarjunan, K.; Lorenzo, J.; Kovačević, D.B. An overview of organosulfur compounds from Allium spp.: From processing and preservation to evaluation of their bioavailability, antimicrobial, and anti-inflammatory properties. Food Chem. 2019, 276, 680–691. [Google Scholar] [CrossRef]
- Fredotović, Ž.; Puizina, J.; Nazlić, M.; Maravić, A.; Ljubenkov, I.; Soldo, B.; Vuko, E.; Bajić, D. Phytochemical Characterization and Screening of Antioxidant, Antimicrobial and Antiproliferative Properties of Allium × cornutum Clementi and Two Varieties of Allium cepa L. Peel Extracts. Plants 2021, 10, 832. [Google Scholar] [CrossRef]
- Schmidt, J.S.; Nyberg, N.; Staerk, D. Assessment of constituents in Allium by multivariate data analysis, high-resolution α-glucosidase inhibition assay and HPLC-SPE-NMR. Food Chem. 2014, 161, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Dalar, A.; Mükemre, M. Traditional Medicinal Plants of Van Province, Eastern Anatolia; NOVA Science Publishers Inc.: New York, NY, USA, 2020; pp. 51–162. [Google Scholar]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Altemimi, A.; Lakhssassi, N.; Baharlouei, A.; Watson, D.G.; Lightfoot, D.A. Phytochemicals: Extraction, isolation, and identification of bioactive compounds from plant extracts. Plants 2017, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.P.; Li, Y.; Meng, X.; Zhou, T.; Zhou, Y.; Zheng, J.; Li, H.B. Natural antioxidants in foods and medicinal plants: Extraction, assessment and resources. Int. J. Mol. Sci. 2017, 18, 96. [Google Scholar] [CrossRef]
- Kim, M.; Jo, S.; Jang, H.; Lee, M.; Kwon, Y. Antioxidant activity and α-glucosidase inhibitory potential of onion (Allium cepa L.) extracts. Food Sci. Biotechnol. 2010, 19, 159–164. [Google Scholar] [CrossRef]
- Snoussi, M.; Trabelsi, N.; Dehmeni, A.; Benzekri, R.; Bouslama, L.; Hajlaoui, B.; Al-sieni, A.; Papetti, A. Phytochemical analysis, antimicrobial and antioxidant activities of Allium roseum var. odoratissimum (Desf.) Coss extracts. Ind. Crops Prod. 2016, 89, 533–542. [Google Scholar] [CrossRef]
- Mollica, A.; Zengin, G.; Locatelli, M.; Picot-Allain, C.; Mahomoodally, M.F. Multidirectional investigations on different parts of Allium scorodoprasum L. subsp. rotundum (L.) Stearn: Phenolic components, in vitro biological, and in silico propensities. Food Res. Int. 2018, 108, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Nickavar, B.; Yousefian, N. Inhibitory Effects of Six Allium Species on α-Amylase Enzyme Activity. Iran. J. Pharm. Res. 2009, 8, 53–57. [Google Scholar]
- Tang, X.; Olatunji, O.; Zhou, Y.; Hou, X. Allium tuberosum: Antidiabetic and hepatoprotective activities. Food Res. Int. 2017, 102, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Vlase, L.; Pârvu, M.; Parvu, E.A.; Toiu, A. Chemical constituents of three Allium species from Romania. Molecules 2012, 18, 114–127. [Google Scholar] [CrossRef]
- Simin, N.; Orčić, D.; Četojević-Simin, D.; Mimica-Dukić, N.; Anačkov, G.; Beara, I.; Mitic-Culafic, D.; Božin, B. Phenolic profile, antioxidant, anti-inflammatory and cytotoxic activities of small yellow onion (Allium flavum L. subsp. flavum, Alliaceae). LWT Food Sci. Technol. 2013, 54, 139–146. [Google Scholar] [CrossRef]
- Singh, P.; Mahajan, V.; Shabeer, T.P.; Banerjee, K.; Jadhav, M.; Kumar, P.; Gopal, J. Comparative evaluation of different Allium accessions for allicin and other allyl thiosulphinates. Ind. Crops Prod. 2020, 147, 112215. [Google Scholar] [CrossRef]
- Fratianni, F.; Ombra, M.N.; Cozzolino, A.; Riccardi, R.; Spigno, P.; Tremonte, P.; Coppola, R.; Nazzaro, F. Phenolic constituents, antioxidant, antimicrobial and anti-proliferative activities of different endemic Italian varieties of garlic (Allium sativum L.). J. Funct. Foods 2016, 21, 240–248. [Google Scholar] [CrossRef]
- Truscheit, E.; Frommer, W.; Junge, B.; Müller, L.; Schmidt, D.D.; Wingender, W. Chemistry and biochemistry of microbial α-Glucosidase inhibitors. Angew. Chem. Int. Ed. Engl. 2010, 20, 744–761. [Google Scholar] [CrossRef]
- Vocadlo, D.J.; Davies, G.J. Mechanistic insights into glycosidase chemistry. Curr. Opin. Chem. Biol. 2008, 12, 539–555. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.I.I.; Vattem, D.A.; Shetty, K. Evaluation of clonal herbs of Lamiaceae species for management of diabetes and hypertension. Asia Pac. J. Clin. Nutr. 2006, 15, 107–118. [Google Scholar] [PubMed]
- Exteberria, U.; Garza, A.L.; Campiόn, J.; Martinez, J.A.; Milagro, F.I. Antidiabetic effects of natural plant extracts via inhibition of carbonhydrate hydrolysis enzymes with emphasis on pancreatic alpha amylase. Expert Opin. Ther. Targets 2012, 16, 269–297. [Google Scholar]
- Garza, A.L.; Milagro, F.; Boqué, N.; Campión, J.; Martínez, J. Natural inhibitors of pancreatic lipase as new players in obesity treatment. Planta Medica 2011, 77, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.; Yen, G. Effects of flavonoids and phenolic acids on the inhibition of adipogenesis in 3T3-L1 adipocytes. J. Agric. Food Chem. 2007, 55, 8404–8410. [Google Scholar] [CrossRef]
- Nishimura, M.; Muro, T.; Kobori, M.; Nishihira, J. Effect of Daily Ingestion of Quercetin-Rich Onion Powder for 12 Weeks on Visceral Fat: A Randomised, Double-Blind, Placebo-Controlled, Parallel-Group Study. Nutrients 2020, 12, 91. [Google Scholar] [CrossRef]
- Kamalakkannan, N.; Prince, P. Antihyperglycaemic and antioxidant effect of rutin, a polyphenolic flavonoid, in streptozotocin-induced diabetic wistar rats. Basic Clin. Pharmacol. Toxicol. 2006, 98, 97–103. [Google Scholar] [CrossRef]
- Vuković, S.; Moravčević, D.; Gvozdanović-Varga, J.; Dojčinović, B.; Vujošević, A.; Pećinar, I.; Kilibarda, S.; Kostić, A.Ž. Elemental Profile, General Phytochemical Composition and Bioaccumulation Abilities of Selected Allium Species Biofortified with Selenium under Open Field Conditions. Plants 2023, 12, 349. [Google Scholar] [CrossRef] [PubMed]
- Dávalos, A.; Gómez-Cordovés, C.; Bartolomé, B. Extending applicability of the oxygen radical absorbance capacity (ORAC-fluorescein) assay. J. Agric. Food Chem. 2004, 52, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Tabart, J.; Kevers, C.; Pincemail, J.; Defraigne, J.; Dommès, J. Comparative antioxidant capacities of phenolic compounds measured by various tests. Food Chem. 2009, 113, 1226–1233. [Google Scholar] [CrossRef]
- Okada, Y.; Tanaka, K.; Sato, E.; Okajima, H. Kinetic and mechanistic studies of allicin as an antioxidant. Org. Biomol. Chem. 2006, 422, 4113–4117. [Google Scholar] [CrossRef] [PubMed]
- Ruberto, G.; Baratta, M.T. Antioxidant activity of selected essential oil components in two lipid model systems. Food Chem. 2000, 69, 167–174. [Google Scholar] [CrossRef]
- Santana-Gálvez, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Chlorogenic Acid: Recent Advances on Its Dual Role as a Food Additive and a Nutraceutical against Metabolic Syndrome. Mol. J. Synth. Chem. Nat. Prod. Chem. 2017, 22, 358. [Google Scholar] [CrossRef] [PubMed]
- Iobbi, V.; Parisi, V.; Lanteri, A.P.; Maggi, N.; Giacomini, M.; Drava, G.; Minuto, G.; Minuto, A.; Tommasi, N.D.; Bisio, A. NMR Metabolite Profiling for the Characterization of Vessalico Garlic Ecotype and Bioactivity against Xanthomonas campestris pv. campestris. Plants 2024, 13, 1170. [Google Scholar] [CrossRef] [PubMed]
- Mai, T.T.; Thu, N.N.; Tien, P.G.; Chuyen, N.V. Alpha-glucosidase inhibitory and antioxidant activities of Vietnamese edible plants and their relationships with polyphenol contents. J. Nutr. Sci. Vitaminol. 2007, 53, 267–276. [Google Scholar] [CrossRef]
- Martinez-Gonzalez, A.I.; Alvarez-Parrilla, E.; Diaz-Sanchez, A.G.; de la Rosa, L.A.; Nunez-Gastelum, J.A.; Vazquez-Flores, A.A.; Gonzalez-Aguilar, G.A. In vitro inhibition of pancreatic lipase by polyphenols: A kinetic, fluorescence spectroscopy and molecular docking study. Food Technol. Biotechnol. 2017, 55, 519–530. [Google Scholar] [CrossRef]
- Dalar, A.; Konczak, I. Phenolic contents, antioxidant capacities and inhibitory activities against key metabolic syndrome relevant enzymes of herbal teas from Eastern Anatolia. Ind. Crops Prod. 2013, 44, 383–390. [Google Scholar] [CrossRef]
- Baytop, T. Türkiye’de Bitkiler ile Tedavi (Geçmişte ve Bugün); Istanbul University Press: İstanbul, Türkiye, 1999. [Google Scholar]
- Dalar, A.; Türker, M.; Konczak, I. Antioxidant capacity and phenolic constituents of Malva neglecta Wallr. and Plantago lanceolata L. from Eastern Anatolia region of Turkey. J. Herb. Med. 2012, 2, 42–51. [Google Scholar] [CrossRef]
- Uzun, Y.; Dalar, A.; Konczak, I. Sempervivum davisii: Phytochemical composition, antioxidant and lipase-inhibitory activities. Pharm. Biol. 2017, 55, 532–540. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).