
Two Carya Species, Carya hunanensis and Carya illinoinensis, Used as Rootstocks Point to Improvements in the Heat Resistance of Carya cathayensis

Abstract
:1. Introduction
2. Results
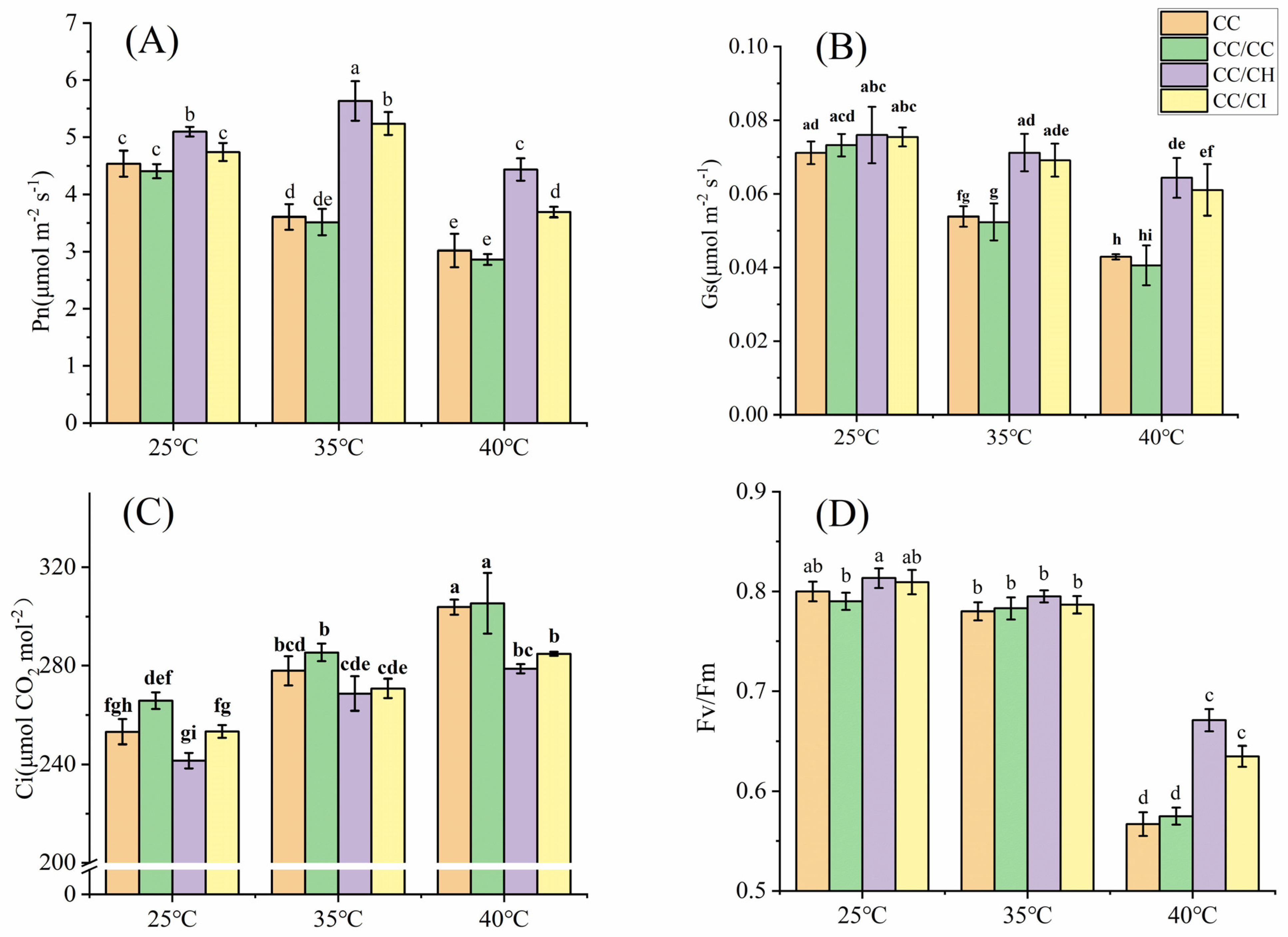
2.1. Effects of Heat Stress on Gas Exchange and Chlorophyll Fluorescence Parameters
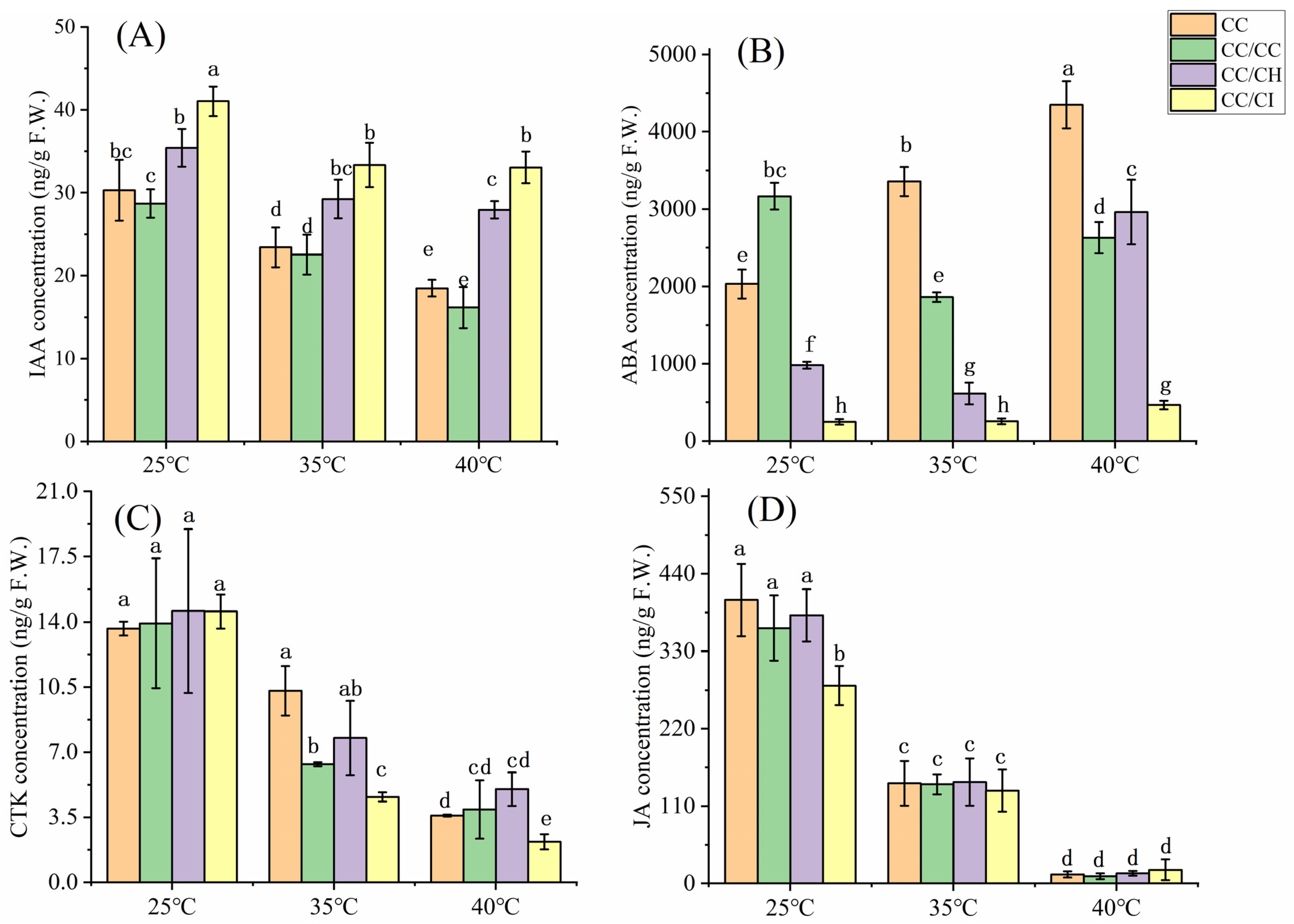
2.2. Effects of Heat Stress on Hormone Contents
2.3. Identification of Differentially Expressed Genes
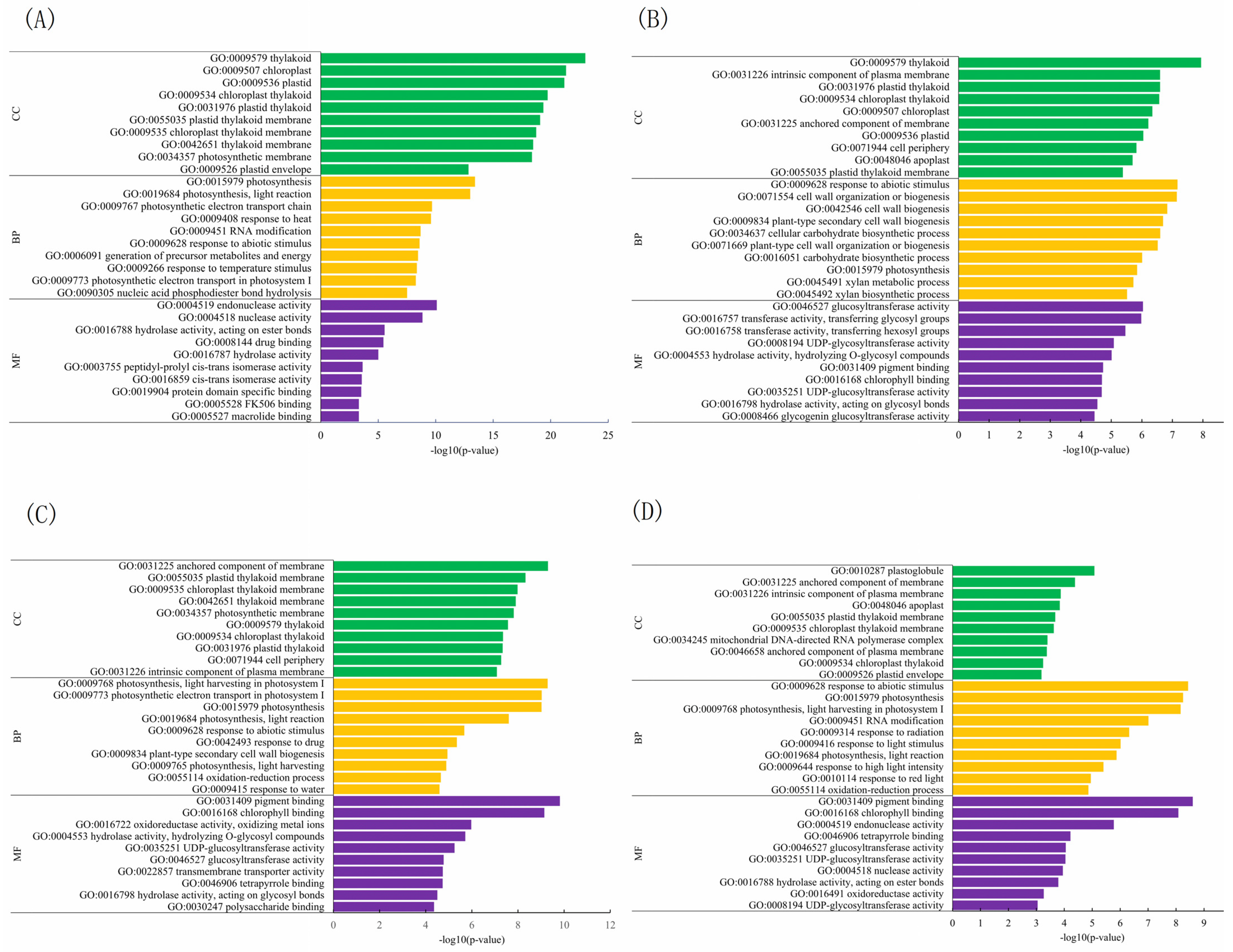
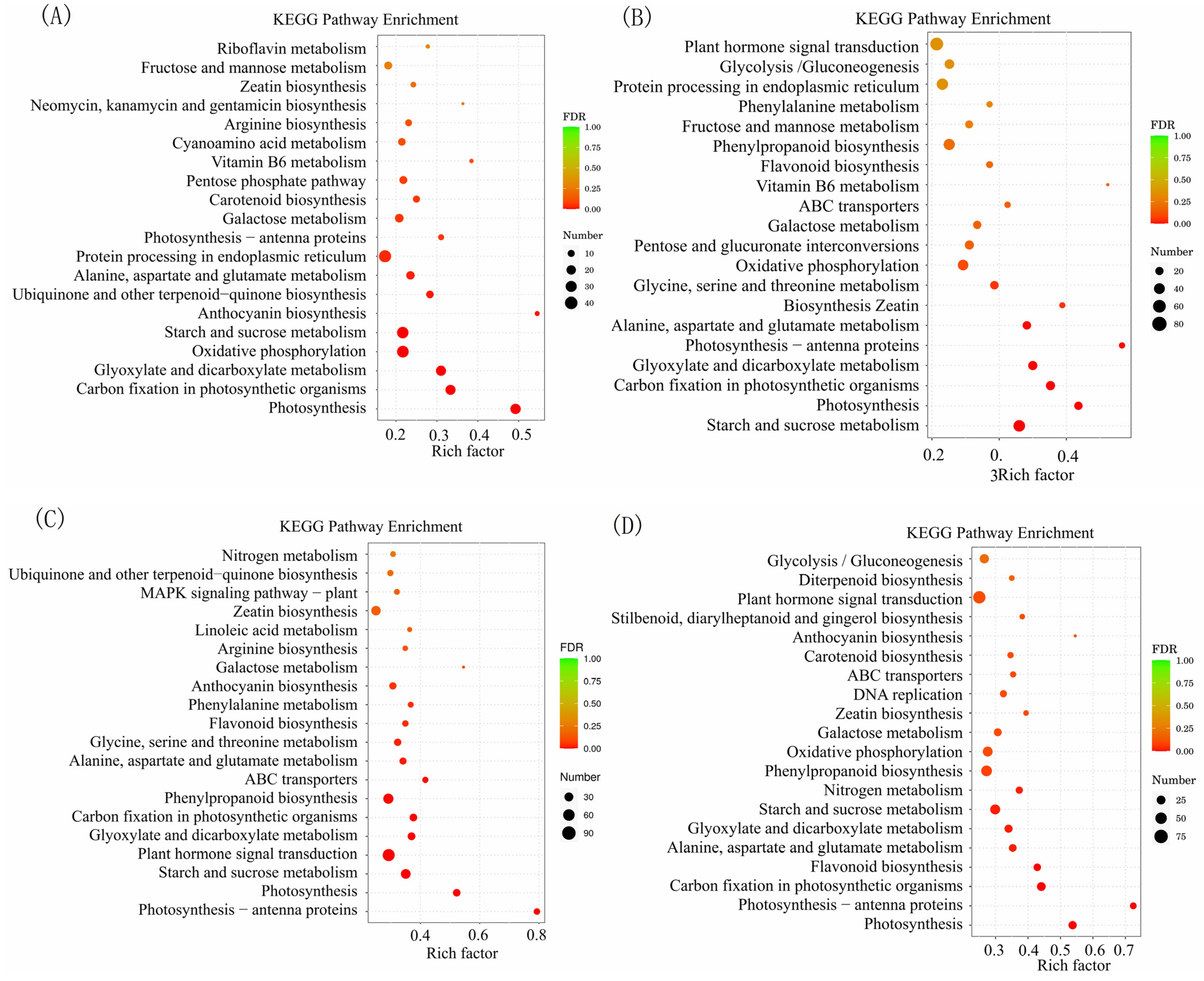
2.4. GO Enrichment and KEGG Pathway Analysis of Differentially Expressed Genes
2.5. Key Differentially Expressed Genes in Response to Heat Stress
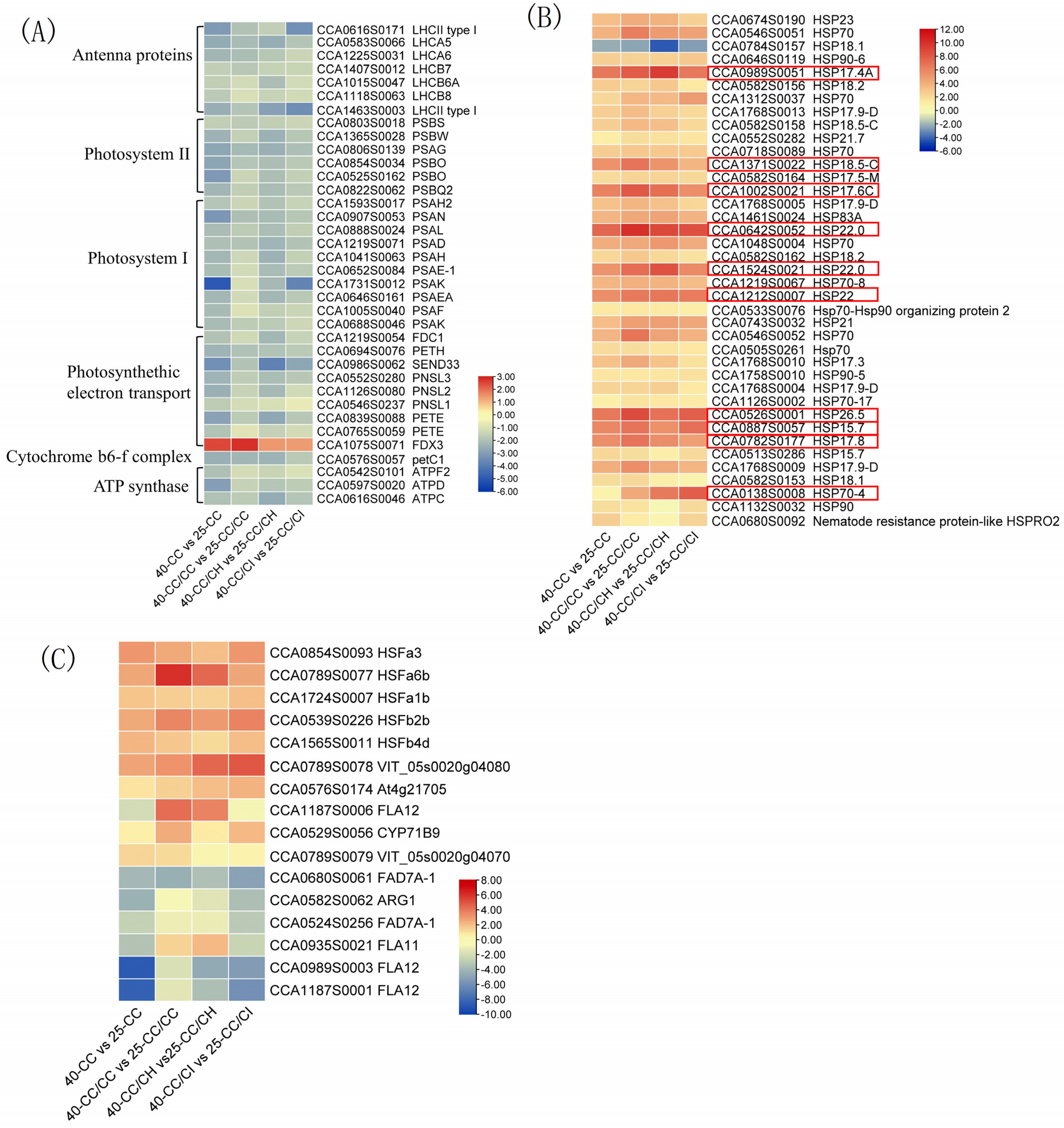
2.5.1. DEGs Involved in Photosynthesis under Heat Stress
2.5.2. Heat Shock Proteins Activated by HS
2.5.3. Differentially Expressed Transcription Factors under HS
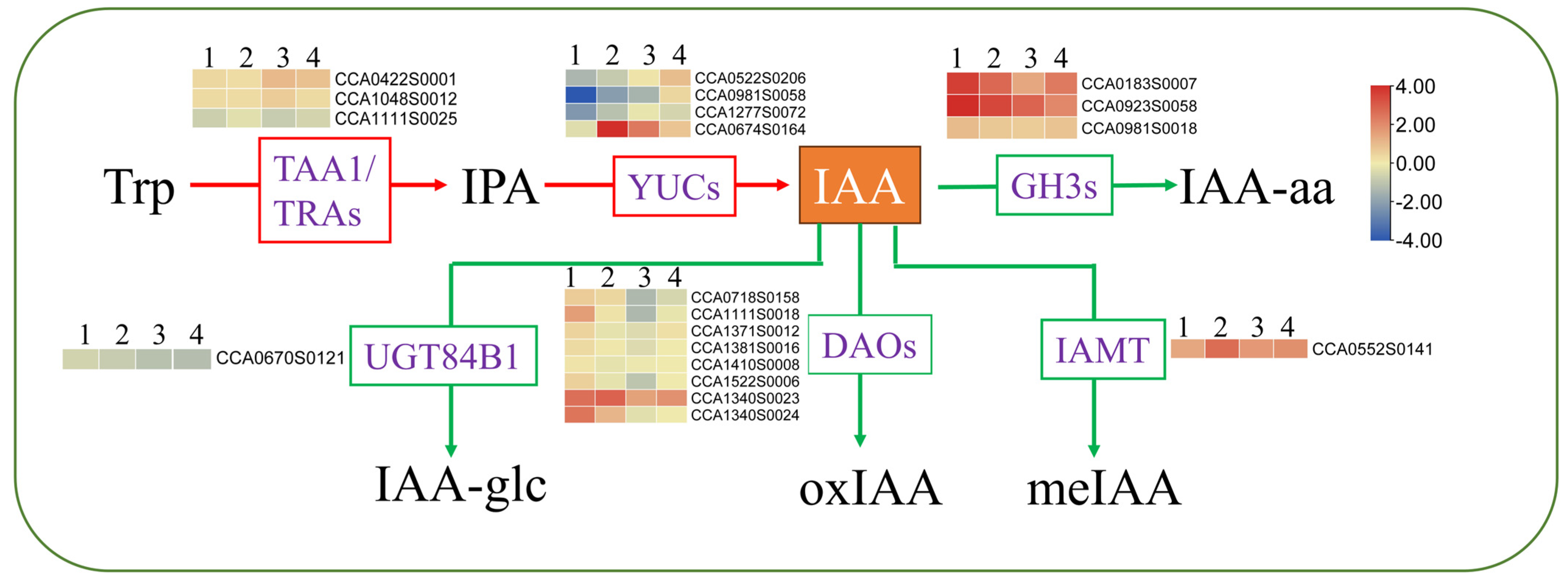
2.5.4. DEGs Involved in Hormones under Heat Stress
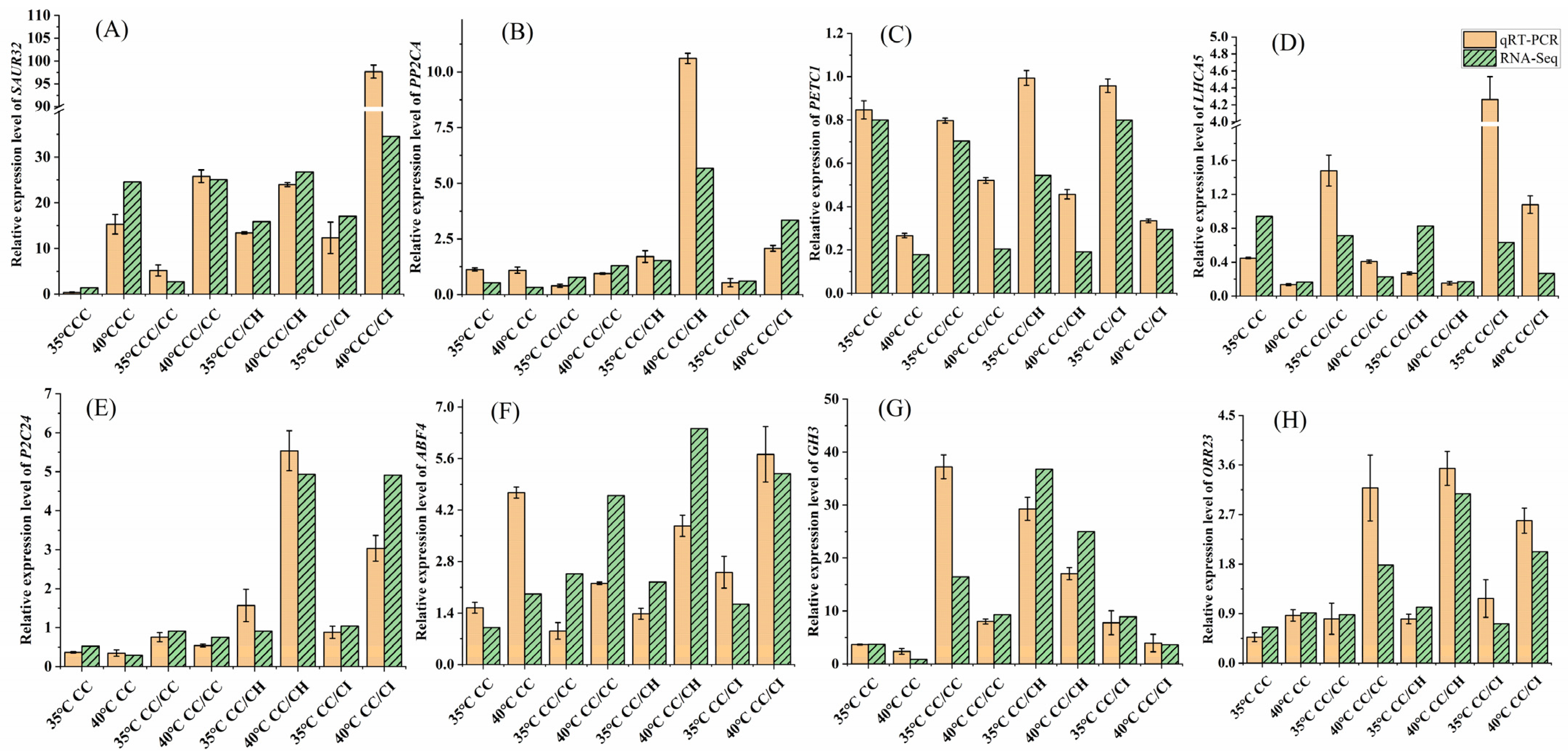
2.6. Validation of Transcription Data Using qRT-PCR
3. Discussion
3.1. Grafting onto C. hunanensis and C. illinoinensis Enhances Photosynthesis of Chinese Hickory under HS
3.2. Grafting onto C. hunanensis and C. illinoinensis Stabilized Phytohormones in Chinese Hickory under HS
3.3. Grafting onto C. hunanensis and C. illinoinensis Activated More Transcription Factors and Heat Shock Proteins in Chinese Hickory under HS
4. Materials and Methods
4.1. Plant Materials and High-Temperature Treatment
4.2. Determination of Gas Exchange and Chlorophyll Fluorescence Parameters
4.3. Phytohormone Extraction and Ultra-High-Performance Liquid Chromatography
4.4. RNA Extraction, cDNA Library Preparation, and Sequencing
4.5. Transcriptome Sequencing (RNA-seq) Data Processing and Analysis
4.6. Quantitative Real-Time PCR
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, N.; Wang, E.L.; Wang, J.; Zhang, J.P.; Zheng, B.R.; Huang, Y.; Tan, M. Modelling maize phenology, biomass growth and yield under contrasting temperature conditions. Agric. For. Meteorol. 2018, 250, 319–329. [Google Scholar] [CrossRef]
- Xu, X.; Wang, L.; Sun, D.; Liu, L.; Banson, K.E. The impact of climate change on yield potential of maize across China. Int. J. Plant Prod. 2017, 11, 47–63. [Google Scholar] [CrossRef]
- Zha, Q.; Xia, X.J.; He, Y.N.; Jiang, A.L. Transcriptomic analysis of the leaves of two grapevine cultivars under high temperature stress. Sci. Hortic. 2020, 265, 109265. [Google Scholar] [CrossRef]
- Wu, J.C.; Lu, Y.; Zhou, B.Y.; Hu, Z.Q. Effects of cytokinin and abscisic acid on heat resistance of Vetiveria zizanioides. Not. Bot. Horti. Agrobo. 2022, 50, 12755. [Google Scholar] [CrossRef]
- Cong, X.; Zhang, Q.; Li, H.T.; Jiang, Z.L.; Cao, R.F.; Gao, S.S.; Tian, W. Puerarin ameliorates heat stress–induced oxidative damage and apoptosis in bovine Sertoli cells by suppressing ROS production and upregulating Hsp72 expression. Theriogenology 2017, 88, 215–227. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Li, L.; Xu, H.; Xi, J.; Cao, X.; Xu, H.; Rong, S.; Dong, Y.; Wang, C.; Chen, R.; et al. A rice jacalin-related mannose-binding lectin gene, OsJRL, enhances Escherichia coli viability under high salinity stress and improves salinity tolerance of rice. Plant Biol. 2017, 19, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Ohama, N.; Sato, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional regulatory network of plant heat stress response. Trends Plant Sci. 2017, 22, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Kotak, S.; Larkindale, J.; Lee, U.; Koskull-Döring, P.; Vierling, E.; Scharf, K.D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Hayer-Hartl, M.; Hartl, U. Molecular chaperones in the cytosol: From nascent chain to folded protein. Science 2002, 295, 1852–1858. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, X.; Goatley, M.; Ervin, E. Heat shock proteins in relation to heat stress tolerance of creeping bentgrass at different N levels. PLoS ONE 2014, 9, e102914. [Google Scholar] [CrossRef]
- Kazan, K. Diverse roles of jasmonates and ethylene in abiotic stress tolerance. Trends Plant Sci. 2015, 20, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.H.; Kumar, V.; Shriram, V.; Sah, S.K. Phytohormones and their metabolic engineering for abiotic stress tolerance in crop plants. Crop J. 2016, 4, 162–176. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, B.; Zaid, A.; Sadiq, Y.; Bashir, S.; Wani, S.H. Role of selective exogenous elicitors in plant responses to abiotic stress tolerance. In Plant Abiotic Stress Tolerance; Springer: Berlin/Heidelberg, Germany, 2019; pp. 273–290. [Google Scholar]
- Sakata, T.; Oshino, T.; Miura, S.; Tomabechi, M.; Tsunaga, Y.; Higashitani, N.; Miyazawa, Y.; Takahashi, H.; Watanabe, M.; Higashitani, A. Auxins reverse plant male sterility caused by high temperatures. Proc. Natl. Acad. Sci. USA 2010, 107, 8569–8574. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Zheng, B.S. Molecular Responses during Plant Grafting and Its Regulation by Auxins, Cytokinins, and Gibberellins. Biomolecules 2019, 9, 397. [Google Scholar] [CrossRef] [PubMed]
- Jha, U.C.; Nayyar, H.; Siddique, K.H.M. Role of Phytohormones in Regulating Heat Stress Acclimation in Agricultural Crops. J. Plant Growth Regul. 2022, 41, 1041–1064. [Google Scholar] [CrossRef]
- Huang, Y.; Xiao, L.; Zhang, Z.; Zhang, R.; Wang, Z.; Huang, C.; Huang, R.; Luan, Y.; Fan, T.; Wang, J.; et al. The genomes of pecan and Chinese hickory provide insights into Carya evolution and nut nutrition. Giga Sci. 2019, 8, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Augstein, F.; Kareem, A.; Melnyk, C.W. Plant grafting: Molecular mechanisms and applications. Mol. Plant 2023, 17, 75–91. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.C.; Zhao, L.; Mei, J.Q.; Abbas, F.; Xie, X.T.; Yang, Y.; Huang, Q.; Wang, J.; Yuan, H.; Sharma, A.; et al. Genome-Wide Identifcation and Expression Analysis of Response Regulators Family Genes in Chinese Hickory (Carya cathayensis) Suggests Their Potential Roles during Grafting. J. Plant Growth Regul. 2023, 42, 5099–5115. [Google Scholar] [CrossRef]
- Zhou, D.; Fei, L.; Huang, R.; Ma, L.; Zhou, X.; Su, X. Analysis of Carya cathayensis resistance to Botryosphaeria dothidea using grafting on pecan in China. J. Plant Pathol. 2023, 105, 955–962. [Google Scholar] [CrossRef]
- Schwarz, D.; Rouphael, Y.; Colla, G.; Venema, J.H. Grafting as a tool to improve tolerance of vegetables to abiotic stresses: Thermal stress, water stress and organic pollutants. Sci. Hortic. 2010, 127, 162–171. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.; Wang, Z.; Guo, X.; Wang, F.; Xia, X.J.; Zhou, J.; Shi, K.; Yu, J.-Q.; Zhou, Y.-H. Microarray and genetic analysis reveals that csa-miR159b plays a critical role in abscisic acid-mediated heat tolerance in grafted cucumber plants. Plant Cell Environ. 2016, 39, 1790–1804. [Google Scholar] [CrossRef] [PubMed]
- Penella, C.; Landi, M.; Guidi, L.; Nebauer, S.G.; Pellegrini, E.; San Bautista, A.; Remorini, D.; Nali, C.; López-Galarza, S.; Calatayud, A. Salt-tolerant rootstock increases yield of pepper under salinity through maintenance of photosynthetic performance and sinks strength. J. Plant Physiol. 2016, 193, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rivero, R.M.; Ruiz, J.M.; Sánchez, E.; Romero, L. Does grafting provide tomato plants an advantage against H2O2 production under conditions of thermal shock? Physiol. Plant. 2003, 117, 44–50. [Google Scholar] [CrossRef]
- Abdelmageed, A.; Gruda, N. Influence of grafting on growth, development and some physiological parameters of tomatoes under controlled heat stress conditions. Eur. J. Hortic. Sci. 2009, 74, 16–20. [Google Scholar] [CrossRef]
- Gisbert-Mullor, R.; Padilla, Y.G.; Martínez-Cuenca, M.; Lopez-Galarza, S.; Calatayud, A. Suitable rootstocks can alleviate the effects of heat stress on pepper plants. Sci. Hortic. 2021, 290, 110529. [Google Scholar] [CrossRef]
- Tao, M.Q.; Jahan, M.S.; Hou, K.; Shu, S.; Wang, Y.; Sun, J.; Guo, S.R. Bitter melon (Momordica charantia L.) rootstock improves the heat tolerance of cucumber by regulating photosynthetic and antioxidant defense pathways. Plants 2020, 9, 692. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.Q.; Zhang, C.L.; Wang, W.J.; Cao, Z.; Li, S.; Li, H.; Zhu, W.; Huang, Y.; Bao, M.; He, Y.; et al. Comparative transcriptome analysis of different heat stress responses between self-root grafting line and heterogeneous grafting line in rose. Hortic. Plant J. 2021, 7, 243–255. [Google Scholar] [CrossRef]
- Goncalves, B.; Moutinho-Pereira, J.; Santos, A.; Solva, P.A.; Bacelar, E.; Correia, C.; Rosa, E. Scion–rootstock interaction affects the physiology and fruit quality of sweet cherry. Tree Physiol. 2005, 26, 93–104. [Google Scholar] [CrossRef]
- Luis, I.S.; Theodore, M.D. The effect of root pressurization on water relations, shoot growth, and leaf gas exchange of peach (Prunus persica) trees on rootstocks with differing growth potential and hydraulic conductance. J. Exp. Bot. 2006, 57, 1981–1989. [Google Scholar] [CrossRef]
- Sean, M.B.; Fiona, M.W.; Peter, H.; Andrew, L.P.; Stephen, J.C.; David, J.J. Modification of gibberellin biosynthesis in the grafted apple scion allows control of tree height independent of the rootstock. Plant Biotechnol. J. 2005, 3, 215–223. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, J.; Xu, Y.; Abbas, F.; Xu, D.; Tao, S.; Xie, X.; Song, F.; Huang, Q.; Sharma, A.; et al. Genome-wide identification and expression analysis of AUX/LAX family genes in Chinese hickory (Carya cathayensis Sarg.). Front. Plant Sci. 2023, 13, 1060965. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Mei, J.; Chen, J.; Yang, Y.; Gu, Y.; Tang, X.; Lu, H.; Yang, K.; Sharma, A.; Wang, X.; et al. Expression analysis of PIN family genes in Chinese hickory reveals their potential roles during grafting and salt stress. Front. Plant Sci. 2022, 13, 999990. [Google Scholar] [CrossRef] [PubMed]
- Goldschmidt, E.E. Plant grafting: New mechanisms, evolutionary implications. Front. Plant Sci. 2014, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Guigón-López, C. Differential development of wilt and stem rot diseases in grafted bell pepper (Capsicum annuum L.). Eur. J. Plant Pathol. 2019, 154, 347–357. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Sun, X.Z.; Zhang, S.; Yang, J.H.; Liu, F.F.; Fan, J. Comprehensive transcriptome analysis of grafting onto Artemisia scoparia W.to affect the aphid resistance of chrysanthemum (Chrysanthemum morifolium). BMC Genom. 2019, 20, 776. [Google Scholar] [CrossRef]
- Fita, A.; Picó, B.; Roig, C.; Nuez, F. Performance of Cucumis melo ssp. agrestis as a rootstock for melon. J. Hortic. Sci. Biotechnol. 2015, 82, 184–190. [Google Scholar] [CrossRef]
- Rafael, D.; Alejandro, P.; Ruiz-Sánchez, M. Physiological responses of apricot plants grafted on two different rootstocks to flooding conditions. J. Plant Physiol. 2002, 159, 725–732. [Google Scholar]
- Yang, Y.; Huang, Q.Y.; Wang, X.F.; Mei, J.Q.; Sharma, A.; Tripathi, D.K.; Yuan, H.; Zheng, B. Genome-wide identification and expression profiles of ABCB gene family in Chinese hickory (Carya cathayensis Sarg.) during grafting. Plant Physiol. Biochem. 2021, 168, 477–487. [Google Scholar] [CrossRef]
- Mathur, S.; Agrawal, D.; Jajoo, A. Photosynthesis: Response to high temperature stress. J. Photochem. Photobiol. B Biol. 2014, 137, 116–126. [Google Scholar] [CrossRef]
- Li, H.; Wang, F.; Chen, X.J.; Shi, K.; Xia, X.J.; Considine, M.J.; Yu, J.-Q.; Zhou, Y.-H. The sub/supra-optimal temperature-induced inhibition of photosynthesis and oxidative damage in cucumber leaves are alleviated by grafting onto figleaf gourd/luffa rootstocks. Physiol. Plant 2014, 152, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yuan, Y.; Du, N.; Wang, Y.; Shu, S.; Sun, J.; Guo, S. Proteomic analysis of heat stress resistance of cucumber leaves when grafted onto Momordica rootstock. Hortic. Res. 2018, 5, 53. [Google Scholar] [CrossRef] [PubMed]
- Faria, T.; Wilkins, D.; Besford, R.T.; Vaz, M.; Pereira, J.S.; Chaves, M.M. Growth at elevated CO2 leads to down-regulation of photosynthesis and altered response to high temperature in Quercus suber L. seedlings. J. Exp. Bot. 1996, 47, 1755–1761. [Google Scholar] [CrossRef]
- Eamus, D.; Taylor, D.T.; Macinnis, C.M.O.; Shanahan, S.; De Silva, L. Comparing model predictions and experimental data for the response of stomatal conductance and guard cell turgor to manipulations of cuticular conductance, leaf-to-air vapour pressure difference and temperature: Feedback mechanisms are able to account for all observations. Plant Cell Environ. 2008, 31, 269–277. [Google Scholar] [PubMed]
- Law, R.D.; Crafts-Brandner, S.J. Inhibition and acclimation of photosynthesis to heat stress is closely correlated with activa-tion of ribulose-1,5-bisphosphate carboxylase/oxygenase. Plant Physiol. 1999, 120, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Havaux, M. Characterization of thermal damage to the photosynthetic electron transport system in potato leaves. Plant Sci. 1993, 94, 19–33. [Google Scholar] [CrossRef]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef]
- Jin, S.H.; Li, X.Q.; Jia, X.L. Genotypic differences in the responses of gas exchange, chlorophyll fluorescence, and antioxidant enzymes to aluminum stress in Festuca arundinacea. Russ. J. Plant Physiol. 2011, 58, 560–566. [Google Scholar] [CrossRef]
- Ren, S.X.; Ma, K.B.; Lu, Z.G.; Chen, G.; Cui, J.W.; Tong, P.X.; Wang, L.; Jin, B.; Chen, G.; Teng, N. Transcriptomic and Metabolomic Analysis of the Heat-Stress Response of Populus tomentosa Carr. Forests 2019, 10, 383. [Google Scholar] [CrossRef]
- Zheng, Z.; Guo, Y.; Novák, O.; Chen, W.; Ljung, K.; Noel, J.P.; Chory, J. Local auxin metabolism regulates environment-induced hypocotyl elongation. Nat. Plants 2016, 2, 16025. [Google Scholar] [CrossRef]
- Chang, B.; Ma, K.B.; Lu, Z.G.; Lu, J.K.; Cui, J.W.; Wang, L.; Jin, B. Physiological, transcriptomic, and metabolic responses of Ginkgo biloba L. to drought, salt, and heat stresses. Biomolecules 2020, 10, 1635. [Google Scholar] [CrossRef] [PubMed]
- Gray, W.M.; Ostin, A.; Sandberg, G.; Romano, C.P.; Estelle, M. High temperature promotes auxin-mediated hypocotyl elongation in Arabidopsis. Proc. Natl. Acad. Sci. USA 1998, 95, 7197–7202. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Dalal, M.; Verma, R.K.; Kumar, S.V.V.; Yadav, S.K.; Pushkar, S.; Kushwaha, S.R.; Bhowmik, A.; Chinnusamy, V. Auxin protects spikelet fertility and grain yield under drought and heat stresses in rice. Environ. Exp. Bot. 2018, 150, 9–24. [Google Scholar] [CrossRef]
- Ivanov, A.G.; Kitcheva, M.I.; Christov, A.M.; Popova, L.P. Effects of abscisic acid treatment on the thermostability of the photosynthetic apparatus in barley chloroplasts. Plant Physiol. 1992, 98, 1228–1232. [Google Scholar] [CrossRef] [PubMed]
- Li, G.Y.; Zhang, C.X.; Zhang, G.H.; Fu, W.M.; Feng, B.H.; Chen, T.T.; Peng, S.; Tao, L.; Fu, G. Abscisic acid negatively modulates heat tolerance in rolled leaf rice by increasing leaf temperature and regulating energy homeostasis. Rice 2020, 13, 18. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chang, H.; Hu, S.; Lu, X.; Yuan, C.; Zhang, C.; Wang, P.; Xiao, W.; Xiao, L.; Xue, G.-P.; et al. Plastid casein kinase2 knockout reduces abscisic acid (ABA) sensitivity, thermotolerance, and expression of ABA-and heat-stressresponsive nuclear genes. J. Exp. Bot. 2014, 65, 4159–4175. [Google Scholar] [CrossRef]
- He, P.; Jin, J.Y. Relationships among hormone changes, transmembrane fux of Ca2+ and lipid peroxidation during leaf senescing in spring maize. Acta Bot. Sin. 1999, 41, 1221–1225. [Google Scholar]
- Liu, X.; Huang, B.; Banowetz, G. Cytokinin effects on creeping bent grass response to heat stress: II. leaf senescence and antioxidant metabolism. Crop Sci. 2002, 42, 466–472. [Google Scholar]
- Wu, C.; Cui, K.; Wang, W.; Li, Q.; Fahad, S.; Hu, Q.; Huang, J.; Nie, L.; Mohapatra, P.K.; Peng, S. Heat-induced cytokinin transportation and degradation are associated with reduced panicle cytokinin expression and fewer spikelets per panicle in Rice. Front. Plant Sci. 2017, 8, 371. [Google Scholar] [CrossRef]
- Chou, C.C.; Chen, W.S.; Huang, K.L.; Yu, H.C.; Liao, L.J. Changes in cytokinin levels of Phalaenopsis leaves at high temperature. Plant Physiol. Biochem. 2000, 38, 309–314. [Google Scholar] [CrossRef]
- Skalák, J.; Cerný, M.; Jedelský, P.; Dobrá, J.; Ge, E.; Novák, J.; Hronková, M.; Dobrev, P.; Vanková, R.; Brzobohatý, B. Stimulation of ipt overexpression as a tool to elucidate the role of cytokinins in high temperature responses of Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 2861–2873. [Google Scholar] [CrossRef] [PubMed]
- Dang, F.F.; Wang, Y.N.; Yu, L.; Eulgem, T.; Lai, Y.; Liu, Z.Q.; Wang, X.; Qiu, A.-L.; Zhang, T.-X.; Lin, J.; et al. CaWRKY40, a WRKY protein of pepper, plays an important role in the regulation of tolerance to heat stress and resistance to Ralstonia solanacearum infection. Plant Cell Environ. 2013, 36, 757. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.L.; Yao, J.; Mei, C.S.; Tong, X.H.; Zeng, L.J.; Li, Q.; Xiao, L.-T.; Sun, T.-P.; Li, J.; Deng, X.-W.; et al. Plant hormone jasmonate prioritizes defense over growth by interfering with gibberellin signaling cascade. Proc. Natl. Acad. Sci. USA 2012, 109, E1192–E1200. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, L. Action of jasmonates in plant stress responses and development and applied aspects. Biotechnol. Adv. 2014, 32, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Tao, P.; Guo, W.L.; Li, B.Y.; Wang, W.H.; Yue, Z.C.; Lei, J.L.; Zhong, X.M. Genome-wide identification, classification, and expression analysis of sHSP genes in Chinese cabbage (Brassica rapa ssp pekinensis). Genet. Mol. Res. 2015, 14, 11975–11993. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.J.; Ye, T.T.; Wang, Q.; Cai, B.D.; Feng, Y.Q. A rapid approach to investigate spatiotemporal distribution of phytohormones in rice. Plant Methods 2016, 12, 47. [Google Scholar] [CrossRef]
- Chen, M.L.; Fu, X.M.; Liu, J.Q.; Ye, T.T.; Hou, S.Y.; Huang, Y.Q.; Yuan, B.-F.; Wu, Y.; Feng, Y.-Q. Highly sensitive and quantitative profiling of acidic phytohormones using derivatization approach coupled with nano-LC-ESI-Q-TOF-MS analysis. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2012, 905, 67–74. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparation | Upregulated | Downregulated | Total |
|---|---|---|---|
| 35-CC/CI vs. 25-CC/CI | 1587 | 2104 | 3691 |
| 40-CC/CI vs. 25-CC/CI | 3324 | 3563 | 6887 |
| 40-CC/CC vs. 25-CC/CC | 3446 | 2512 | 5958 |
| 35-CC vs. 25-CC | 345 | 313 | 658 |
| 35-CC/CH vs. 25-CC/CH | 2442 | 1374 | 3816 |
| 35-CC/CC vs. 25-CC/CC | 1435 | 2066 | 3501 |
| 40-CC/CH vs. 25-CC/CH | 3475 | 3583 | 7058 |
| 40-CC vs. 25-CC | 2015 | 1730 | 3745 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.; Xu, Y.; Li, X.; Ye, B.; Jin, S. Two Carya Species, Carya hunanensis and Carya illinoinensis, Used as Rootstocks Point to Improvements in the Heat Resistance of Carya cathayensis. Plants 2024, 13, 1967. https://doi.org/10.3390/plants13141967
Huang S, Xu Y, Li X, Ye B, Jin S. Two Carya Species, Carya hunanensis and Carya illinoinensis, Used as Rootstocks Point to Improvements in the Heat Resistance of Carya cathayensis. Plants. 2024; 13(14):1967. https://doi.org/10.3390/plants13141967
Chicago/Turabian StyleHuang, Shanxia, Yanxia Xu, Xueqin Li, Boyu Ye, and Songheng Jin. 2024. "Two Carya Species, Carya hunanensis and Carya illinoinensis, Used as Rootstocks Point to Improvements in the Heat Resistance of Carya cathayensis" Plants 13, no. 14: 1967. https://doi.org/10.3390/plants13141967
APA StyleHuang, S., Xu, Y., Li, X., Ye, B., & Jin, S. (2024). Two Carya Species, Carya hunanensis and Carya illinoinensis, Used as Rootstocks Point to Improvements in the Heat Resistance of Carya cathayensis. Plants, 13(14), 1967. https://doi.org/10.3390/plants13141967