


The Influence of Logging-Related Soil Disturbance on Pioneer Tree Regeneration in Mixed Temperate Forests
 ,
,  , ,
, ,  ,
,  and
and
Abstract
1. Introduction
- The changes over time in the soil physical and chemical characteristics that influence the growth of seedlings.
- The ability of alder and maple seedlings to grow on compact forest soils and penetrate them with their roots.
- The potential use of alder and maple planting to enhance the structural regeneration of compacted forest soils.
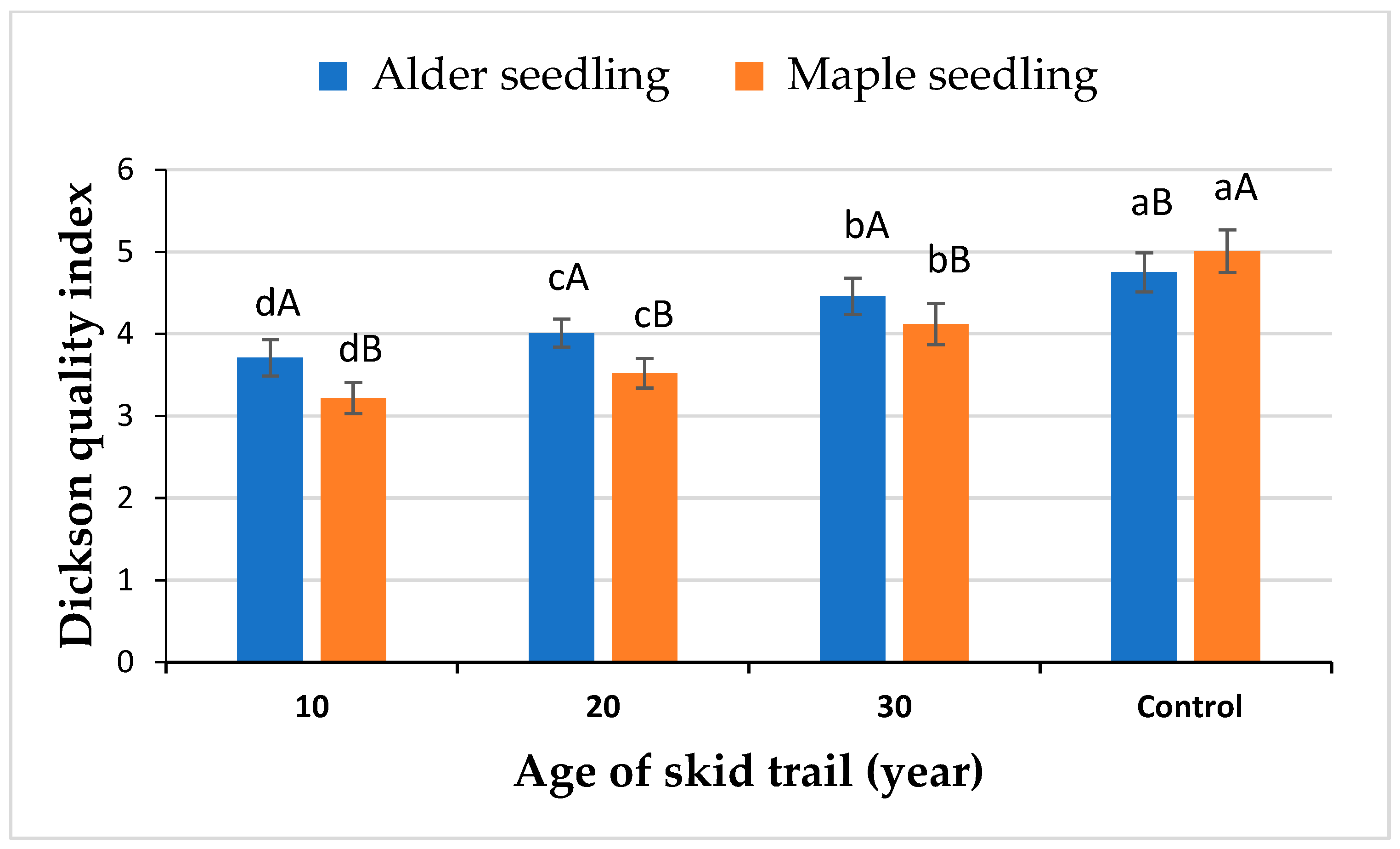
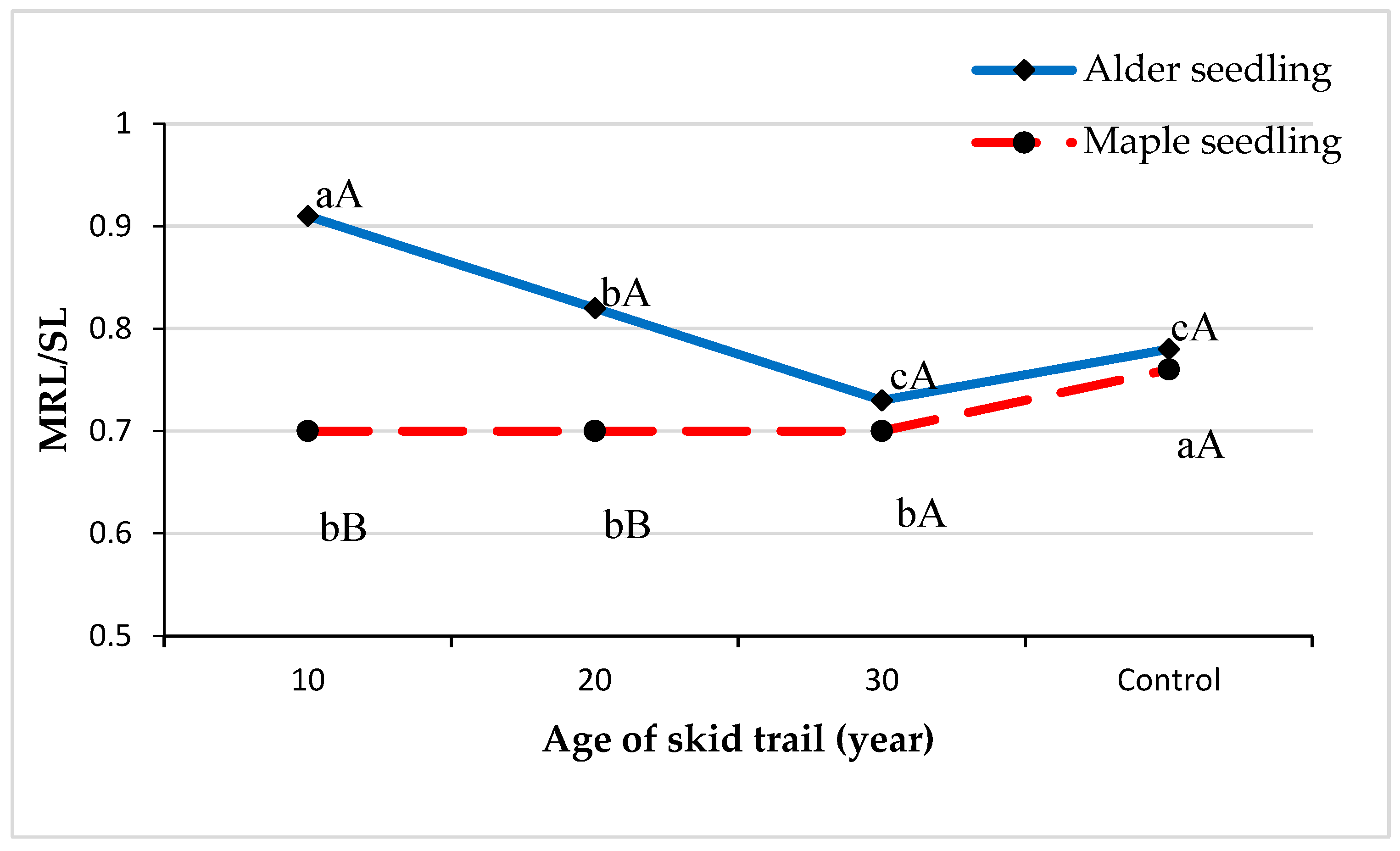
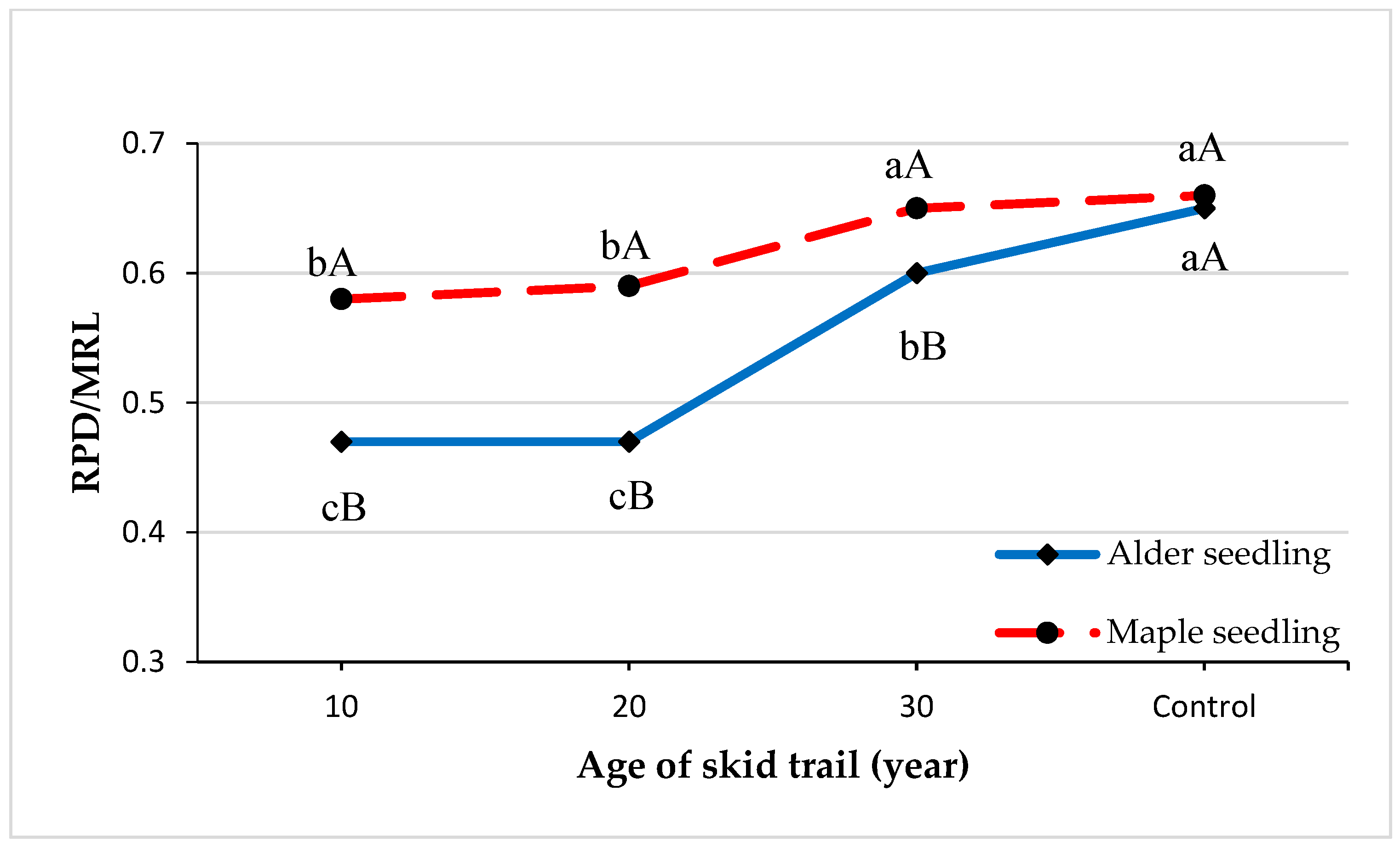
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Area
4.2. Research Design and Data Collection
4.2.1. Soil Sampling
4.2.2. Seedling Sampling
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cambi, M.; Hoshika, Y.; Mariotti, B.; Paoletti, E.; Picchio, R.; Venanzi, R.; Marchi, E. Compaction by a forest machine affects soil quality and Quercus robur L. seedling performance in an experimental field. For. Ecol. Manag. 2017, 384, 406–414. [Google Scholar] [CrossRef]
- Robinson, D.; Emmett, B.; Reynolds, B.; Rowe, E.; Spurgeon, D.; Keith, A.; Lebron, I.; Hockley, N.; Hester, R.; Harrison, R. Soil Natural Capital and Ecosystem Service Delivery in a World of Global Soil Change. Soils Food Secur. 2012, 35, 41–68. [Google Scholar]
- Picchio, R.; Mederski, P.S.; Tavankar, F. How and how much, do harvesting activities affect forest soil, regeneration and stands? Curr. For. Rep. 2020, 6, 115–128. [Google Scholar] [CrossRef]
- Schweier, J.; Magagnotti, N.; Labelle, E.R.; Athanassiadis, D. Sustainability impact assessment of forest operations: A review. Curr. For. Rep. 2019, 5, 101–113. [Google Scholar] [CrossRef]
- Ganatsios, H.P.; Tsioras, P.A.; Papaioannou, A.G.; Blinn, C.R. Short term impacts of harvesting operations on soil chemical properties in a mediterranean oak ecosystem. Croat. J. For. Eng. 2021, 42, 463–476. [Google Scholar] [CrossRef]
- Page-Dumroese, D.; Jurgensen, M.; Elliot, W.; Rice, T.; Nesser, J.; Collins, T.; Meurisse, R. Soil quality standards and guidelines for forest sustainability in northwestern North America. For. Ecol. Manag. 2000, 138, 445–462. [Google Scholar] [CrossRef]
- Hartmann, M.; Niklaus, P.A.; Zimmermann, S.; Schmutz, S.; Kremer, J.; Abarenkov, K.; Lüscher, P.; Widmer, F.; Frey, B. Resistance and resilience of the forest soil microbiome to logging-associated compaction. ISME J. 2014, 8, 226–244. [Google Scholar] [CrossRef]
- Grigal, D.F. Effects of Extensive Forest Management on Soil Productivity. For. Ecol. Manag. 2000, 138, 167–185. [Google Scholar] [CrossRef]
- Marshall, V.G. Impacts of Forest Harvesting on Biological Processes in Northern Forest Soils. For. Ecol. Manag. 2000, 133, 43–60. [Google Scholar] [CrossRef]
- Uusitalo, J.; Ala-Ilomäki, J.; Lindeman, H.; Toivio, J.; Siren, M. Predicting rut depth induced by an 8-wheeled forwarder in fine-grained boreal forest soils. Ann. For. Sci. 2020, 77, 42. [Google Scholar] [CrossRef]
- Labelle, E.R.; Lemmer, K.J. Selected environmental impacts of forest harvesting operations with varying degree of mechanization. Croat. J. For. Eng. 2019, 40, 239–257. [Google Scholar] [CrossRef]
- Mederski, P.S.; Borz, S.A.; Đuka, A.; Lazdiņš, A. Challenges in forestry and forest engineering–Case studies from four countries in East Europe. Croat. J. For. Eng. J. Theory Appl. For. Eng. 2021, 42, 117–134. [Google Scholar]
- Agherkakli, B.; Najafi, A.; Sadeghi, S.H. Ground based operation effects on soil disturbance by steel tracked skidder in a steep slope of forest. J. For. Sci. 2010, 56, 278–284. [Google Scholar] [CrossRef]
- Ampoorter, E.; Van Nevel, L.; De Vos, B.; Hermy, M.; Verheyen, K. Assessing the effects of initial soil characteristics, machine mass and traffic intensity on forest soil compaction. Forest Ecol. Manag. 2010, 260, 1664–1676. [Google Scholar] [CrossRef]
- Eliasson, L. Effects of forwarder tyre pressure on rut formation and soil compaction. Silva Fenn. Monogr. 2005, 39, 549–557. [Google Scholar] [CrossRef]
- Mohieddinne, H.; Brasseur, B.; Spicher, F.; Gallet-Moron, E.; Buridant, J.; Kobaissi, A.; Horen, H. Physical recovery of forest soil after compaction by heavy machines, revealed by penetration resistance over multiple decades. For. Ecol. Manag. 2019, 449, 117472. [Google Scholar] [CrossRef]
- Håkansson, R.C. Reeder Subsoil compaction by vehicles with high axle load—Extent, persistence and crop response. Soil Tillage Res. 1994, 29, 277–304. [Google Scholar] [CrossRef]
- Proto, A.R.; Macrì, G.; Sorgonà, A.; Zimbalatti, G. Impact of skidding operations on soil physical properties in southern Italy. Contemp. Eng. Sci. 2016, 9, 1095–1104. [Google Scholar] [CrossRef]
- Eroğlu, H.; Sariyildiz, T.; Küçük, M.; Sancal, E. The effects of different logging techniques on the physical and chemical characteristics of forest soil. Balt. For. 2016, 22, 139–147. [Google Scholar]
- Nazari, M.; Eteghadipour, M.; Zarebanadkouki, M.; Ghorbani, M.; Dippold, M.A.; Bilyera, N.; Zamanian, K. Impacts of logging-associated compaction on forest soils: A meta-analysis. Front. For. Glob. Change 2021, 4, 780074. [Google Scholar] [CrossRef]
- Basset, C.; Abou Najm, M.; Ghezzehei, T.; Hao, X.; Daccache, A. How does soil structure affect water infiltration? A meta-data systematic review. Soil. Tillage Res. 2023, 226, 105577. [Google Scholar] [CrossRef]
- Malvar, M.C.; Silva, F.C.; Prats, S.A.; Vieira, D.C.S.; Coelho, C.O.A.; Keizer, J.J. Short-term effects of post-fire salvage logging on runoff and soil erosion. For. Ecol. Manag. 2017, 400, 555–567. [Google Scholar] [CrossRef]
- Tavankar, F.; Picchio, R.; Nikooy, M.; Jourgholami, M.; Latterini, F.; Venanzi, R. Effect of soil moisture on soil compaction during skidding operations in poplar plantation. Int. J. For. Eng. 2021, 32, 128–139. [Google Scholar] [CrossRef]
- Bens, O.; Wahl, N.A.; Fischer, H.; Hüttl, R.F. Water infiltration and hydraulic conductivity in sandy cambisols: Impacts of forest transformation on soil hydrological properties. Eur. J. For. Res. 2007, 126, 101–109. [Google Scholar] [CrossRef]
- Wagenbrenner, J.W.; Robichaud, P.R.; Brown, R.E. Rill erosion in burned and salvage logged western montane forests: Effects of logging equipment type, traffic level, and slash treatment. J. Hydrol. 2016, 541, 889–901. [Google Scholar] [CrossRef]
- Hansson, L.; Šimůnek, J.; Ring, E.; Bishop, K.; Gärdenäs, A.I. Soil compaction effects on root-zone hydrology and vegetation in boreal forest clearcuts. Soil Sci. Soc. Am. J. 2019, 83, 105–115. [Google Scholar] [CrossRef]
- Jourgholami, M. Effects of soil compaction on growth variables in Cappadocian maple (Acer cappadocicum) seedlings. J. For. Res. 2018, 29, 601–610. [Google Scholar] [CrossRef]
- Tavankar, F.; Nikooy, M.; Ezzati, S.; Jourgholami, M.; Latterini, F.; Venanzi, R.; Picchio, R. Long-term assessment of soil physicochemical properties and seedlings establishment after skidding operations in mountainous mixed hardwoods. Eur. J. For. Res. 2022, 141, 571–585. [Google Scholar] [CrossRef]
- Jourgholami, M.; Soltanpour, S.; Etehadi Abari, M.; Zenner, E.K. Influence of slope on physical soil disturbance due to farm tractor forwarding in a Hyrcanian forest of northern Iran. iForest. 2014, 7, 342–348. [Google Scholar] [CrossRef]
- Warlo, H.; Zimmermann, S.; Lang, F.; Schack-Kirchner, H. Characteristics of Soil Structure and Greenhouse Gas Fluxes on Ten-Year Old Skid Trails with and without Black Alders (Alnus glutinosa (L.) Gaertn.). Soil Syst. 2022, 6, 43. [Google Scholar] [CrossRef]
- Schäffer, J. Recovery of Soil Structure and Fine Root Distribution in Compacted Forest Soils. Soil Syst. 2022, 6, 49. [Google Scholar] [CrossRef]
- Bejarano, M.D.; Villar, R.; Murillo, A.M.; Quero, J.L. Effects of soil compaction and light on growth of Quercus pyrenaica Willd. (Fagaceae) seedlings. Soil. Till. Res. 2010, 110, 108–114. [Google Scholar] [CrossRef]
- Picchio, R.; Tavankar, F.; Nikooy, M.; Pignatti, G.; Venanzi, R.; Lo Monaco, A. Morphology, Growth and Architecture Response of Beech (Fagus orientalis Lipsky) and Maple Tree (Acer velutinum Boiss.) Seedlings to Soil Compaction Stress Caused by Mechanized Logging Operations. Forests 2019, 10, 771. [Google Scholar] [CrossRef]
- Jourgholami, M.; Khoramizadeh, A.; Zenner, E.K. Effects of soil compaction on seedling morphology, growth, and architecture of chestnut-leaved oak (Quercus castaneifolia). iForest 2016, 10, 145–153. [Google Scholar] [CrossRef]
- Singh, S.; Malik, Z.A.; Sharma, C.M. Tree species richness, diversity, and regeneration status in different oak (Quercus spp.) dominated forests of Garhwal Himalaya, India. J. Asia-Pac. Biodivers. 2016, 9, 293–300. [Google Scholar] [CrossRef]
- Jamshidi, R.; Jaeger, D.; Dragovich, D. Establishment of pioneer seedling species on compacted skid tracks in a temperate Hyrcanian Forest, northern Iran. Environ. Earth Sci. 2018, 77, 1143. [Google Scholar] [CrossRef]
- Sabeti, H. Forests, Trees and Shrubs of Iran; University of Yazd: Yazd, Iran, 1994; p. 810. [Google Scholar]
- Perez, J.; Salazar, R.C.; Stokes, A. An open access database of plant species useful for controlling soil erosion and substrate mass movement. Ecol. Eng. 2017, 99, 530–534. [Google Scholar] [CrossRef]
- Liu, H.; Mao, Z.; Wang, Y.; Kim, J.H.; Bourrier, F.; Mohamed, A.; Stokes, A. Slow recovery from soil disturbance increases susceptibility of high elevation forests to landslides. For. Ecol. Manag. 2021, 485, 118891. [Google Scholar] [CrossRef]
- Ezzati, S.; Najafi, A.; Rab, A.; Zenner, E.K. Recovery of Soil Bulk Density, Porosity and Rutting from Ground Skidding Over a 20-Year Period after Timber Harvesting in Iran. Silva Fenn. 2012, 46, 521–538. [Google Scholar] [CrossRef]
- Klaes, B.; Struck, J.; Schneider, R.; Schüler, G. Middle-term effects after timber harvesting with heavy machinery on a fine-textured forest soil. Eur. J. Forest Res. 2016, 135, 1083–1095. [Google Scholar] [CrossRef]
- Pousse, N.; Bonnaud, P.; Legout, A.; Darboux, F.; Ranger, J. Forest Soil Penetration Resistance Following Heavy Traffic: A 10-YearField Study. Soil Use Manag. 2022, 38, 815–835. [Google Scholar] [CrossRef]
- Kabzems, R.; Haeussler, S. Soil properties, aspen, and white spruce responses 5 years after organic matter removal and compaction treatments. Can. J. For. Res. 2005, 35, 2045–2055. [Google Scholar] [CrossRef]
- Maloney, K.O.; Garten, C.T.; Ashwood, T.L. Changes in soil properties following 55 years of secondary Forest succession at fort Benning, Georgia, U.S.A. Restor. Ecol. 2008, 16, 503–510. [Google Scholar] [CrossRef]
- Sohrabi, H.; Jourgholami, M.; Tavankar, F.; Venanzi, R.; Picchio, R. Post-Harvest Evaluation of Soil Physical Properties and Natural Regeneration Growth in Steep-Slope Terrains. Forests 2019, 10, 1034. [Google Scholar] [CrossRef]
- Kiumarsi, F.; Jourgholami, M.; Jafari, M.; Lo Monaco, A.; Venanzi, R.; Picchio, R. Restoring soil properties in the Hyrcanian forests from machine induced compaction: Reforestation of N2-fixing black alder (Alnus glutinosa (L.) Gaertn.). Land Degrad. Dev. 2024, 35, 2084–2096. [Google Scholar] [CrossRef]
- Alexander, A.B. Soil compaction on skid trails after selective logging in moist evergreen forest of Ghana. Agr. Biol. J. N. Am. 2012, 3, 262–264. [Google Scholar] [CrossRef]
- Gomez, A.; Powers, R.F.; Singer, M.J.; Horwath, W.R. Soil compaction effects on growth of young ponderosa pine following litter removal in California’s Sierra Nevada. Soil Sci. Soc. Am. J. 2002, 66, 1334–1343. [Google Scholar] [CrossRef]
- Brais, S. Persistence of soil compaction and effects on seedling growth in Northwestern Quebec. Soil Sci. Soc. Am. J. 2001, 65, 1263–1271. [Google Scholar] [CrossRef]
- Alameda, D.; Villar, R. Linking root traits to plant physiology and growth in Fraxinus angustifolia Vahl. seedlings under soil compaction conditions. Environ. ExBot 2012, 79, 49–57. [Google Scholar] [CrossRef]
- Bassett, I.E.; Simcock, R.C.; Mitchell, N.D. Consequences of soil compaction for seedling establishment: Implications for natural regeneration and restoration. Austral Ecol. 2005, 30, 827–833. [Google Scholar] [CrossRef]
- Flores Fernández, L.; Rubin, P.; Hartmann, H.; Puhlmann, K. von Wilpert, Initial recovery of soil structure of a compacted forest soil can be enhanced by technical treatments and planting. For. Ecol. Manag. 2019, 431, 54–62. [Google Scholar] [CrossRef]
- Jourgholami, M.; Khoramizadeh, A.; Lo Monaco, A.; Venanzi, R.; Latterini, F.; Tavankar, F.; Picchio, R. Evaluation of Leaf Litter Mulching and Incorporation on Skid Trails for the Recovery of Soil Physico-Chemical and Biological Properties of Mixed Broadleaved Forests. Land 2021, 10, 625. [Google Scholar] [CrossRef]
- Battigelli, J.P.; Spence, J.R.; Langor, D.W.; Berch, S.M. Short-term impact of forest soil compaction and organic matter removal on soil mesofauna density and oribatid mite diversity. Can. J. For. Res. 2004, 34, 1136–1149. [Google Scholar] [CrossRef]
- Sohrabi, H.; Jourgholami, M.; Lo Monaco, A.; Picchio, R. Effects of Forest Harvesting Operations on the Recovery of Earthworms and Nematodes in the Hyrcanain Old-Growth Forest: Assessment, Mitigation, and Best Management Practice. Land 2022, 11, 746. [Google Scholar] [CrossRef]
- Thomas, G.W. Soil pH and soil acidity. In Methods of Soil Analysis, Part 3—Chemical Methods; Sparks, D.L., Ed.; Soil Science Society of America: Madison, WI, USA, 1996; pp. 475–490. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total Carbone, organic Carbone, and organic matter. In Methods of Soil Analysis, Part 3—Chemical Methods; Sparks, D.L., Ed.; Soil Science Society of America: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Bremner, J.M. Nitrogen-total. In Methods of Soil Analysis, Part 3—Chemical Methods; Sparks, D.L., Ed.; Soil Science Society of America: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Kuhlemeier, C. Phyllotaxis. Trends Plant Sci. 2007, 12, 143–150. [Google Scholar] [CrossRef]
- Nyoka, B.I.; Kamanga, R.; Njoloma, J.; Jamnadass, R.; Mng’omba, S.; Muwanje, S. Quality of Tree Seedlings Produced in Nurseries in Malawi: An Assessment of Morphological Attributes. For. Trees Livelihoods 2018, 27, 103–117. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Soil Properties | Age of Skid Trail (Year) | F Value | |||
|---|---|---|---|---|---|
| 10 | 20 | 30 | Control | ||
| Bulk density (g/cm3) | 1.26 ± 0.05 a | 1.20 ± 0.04 a | 1.10 ± 0.04 b | 1.02 ± 0.05 b | 136.4 ** |
| Porosity (%) | 52.4 ± 1.65 b | 54.7 ± 1.82 b | 58.5 ± 1.94 a | 61.5 ± 3.01 a | 107.3 ** |
| Soil penetration resistance (MPa) | 0.46 ± 0.05 a | 0.38 ± 0.06 b | 0.29 ± 0.04 c | 0.26 ± 0.05 c | 215.1 ** |
| Moisture (%) | 37.8 ± 1.70 b | 40.1 ± 1.88 a b | 44.0 ± 2.02 a | 44.5 ± 2.10 a | 33.9 ** |
| Organic matter (%) | 2.64 ± 1.08 d | 2.96 ± 0.07 c | 3.70 ± 0.07 b | 4.08 ± 0.11 a | 59.9 ** |
| Nitrogen (%) | 0.22 ± 0.04 c | 0.26 ± 0.03 c | 0.39 ± 0.11 b | 0.51 ± 0.05 a | 45.7 ** |
| C/N ratio | 12.00 ± 0.07 a | 11.38 ± 0.06 a | 9.49 ± 0.06 b | 8.00 ± 0.08 c | 68.4 ** |
| pH (1:1 H2O) | 5.50 ± 0.06 b | 5.56 ± 0.06 b | 5.79 ± 0.11 a | 5.88 ± 0.10 a | 65.7 ** |
| Seedling Growth Characteristics | Seedling Species | Age of Skid Trail (Year) | F Value | |||
|---|---|---|---|---|---|---|
| 10 | 20 | 30 | Control | |||
| Stem length (cm) | Alder | 49.0 ± 2.5 cA | 57.0 ± 3.3 bA | 68.5 ± 3.3 aA | 69.0 ± 3.1 aA | 77.0 ** |
| Maple | 48.8 ± 3.0 bA | 50.2 ± 3.3 bB | 57.6 ± 3.5 aB | 60.2 ± 2.7 aB | 79.4 * | |
| Stem height (cm) | Alder | 46.1 ± 2.4 cA | 54.5 ± 2.6 bA | 65.6 ± 2.7 aA | 66.7 ± 3.2 aA | 67.2 ** |
| Maple | 45.0 ± 2.1 cA | 47.3 ± 2.3 bB | 54.5 ± 3.0 aB | 57.2 ± 2.8 aB | 96.1 ** | |
| Stem diameter (mm) | Alder | 3.75 ± 0.6 dA | 4.10 ± 0.5 cA | 4.69 ± 0.7 aA | 4.71 ± 0.5 aA | 29.7 ** |
| Maple | 3.71 ± 0.6 dA | 3.91 ± 0.6 cB | 4.31 ± 0.8 bB | 4.45 ± 0.5 aB | 61.5 ** | |
| Main root length (cm) | Alder | 44.7 ± 2.2 cA | 46.9 ± 2.8 cA | 50.0 ± 2.2 bA | 54.3 ± 2.5 aA | 39.5 ** |
| Maple | 34.7 ± 2.6 cB | 35.1 ± 2.5 cB | 40.2 ± 2.5 bB | 45.6 ± 2.3 aB | 23.7 ** | |
| Main root diameter (mm) | Alder | 3.80 ± 0.5 dA | 4.20 ± 0.6 cA | 4.72 ± 0.6 bA | 4.88 ± 0.7 aA | 99.4 ** |
| Maple | 3.81 ± 0.6 dA | 4.07 ± 0.5 cB | 4.50 ± 0.5 bB | 4.62 ± 0.7 aB | 112.4 ** | |
| Lateral root length (cm) | Alder | 49.3 ± 2.6 bA | 50.5 ± 3.2 bA | 57.1 ± 3.3 aA | 59.0 ± 3.2 aA | 26.9 ** |
| Maple | 36.4 ± 2.5 bB | 37.9 ± 2.2 bB | 50.9 ± 2.7 aB | 51.5 ± 3.1 aB | 23.7 ** | |
| Root penetration length (cm) | Alder | 20.9 ± 2.7 cA | 21.7 ± 2.2 cA | 30.1 ± 2.5 bA | 35.2 ± 2.0 aA | 44.0 ** |
| Maple | 20.2 ± 2.6 cA | 20.6 ± 2.5 cA | 26.1 ± 2.4 bB | 30.0 ± 2.0 aB | 31.5 ** | |
| Biomass of Seedlings | Seedling Species | Age of Skid Trail (Year) | F Value | |||
|---|---|---|---|---|---|---|
| 10 | 20 | 30 | Control | |||
| Stem biomass (g) | Alder | 33.0 ± 2.0 bA | 35.2 ± 2.2 bA | 41.5 ± 2.4 aA | 43.2 ± 2.3 aA | 21.4 ** |
| Maple | 35.5 ± 2.1 bA | 34.0 ± 2.6 bA | 35.2 ± 2.1 bB | 42.4 ± 2.5 aA | 17.5 ** | |
| Root biomass (g) | Alder | 22.2 ± 1.7 cA | 24.0 ± 2.1 b cA | 26.8 ± 2.0 bA | 30.7 ± 2.0 aA | 28.2 ** |
| Maple | 21.0 ± 1.5 bA | 22.8 ± 1.6 bA | 23.2 ± 1.8 bA | 29.2 ± 2.0 aA | 14.8 ** | |
| Total biomass (g) | Alder | 55.5 ± 2.4 dA | 59.2 ± 2.6 cA | 68.3 ± 3.1 bA | 73.9 ± 3.1 aA | 54.7 ** |
| Maple | 53.5 ± 2.3 cA | 56.8 ± 2.5 bA | 58.4 ± 2.6 bB | 71.6 ± 2.8 aA | 51.0 ** | |
| Soil Physical and Chemical Properties | Seedling Species | SH (cm) | SL (cm) | SD (mm) | MRL (cm) | MRD (mm) | LRL (cm) | RPD (cm) | SDB (g) | RDB (g) | TDB (g) | DQI |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BD (g/cm3) | Alder | −0.49 ** | −0.48 ** | −0.45 ** | −0.83 ** | −0.58 ** | −0.56 ** | −0.72 ** | −0.49 ** | −0.51 ** | −0.59 ** | −0.66 ** |
| Maple | −0.55 ** | −0.58 ** | −0.48 ** | −0.85 ** | −0.58 ** | −0.55 ** | −0.79 ** | −0.53 ** | −0.55 ** | −0.64 ** | −0.70 ** | |
| TP (%) | Alder | 0.55 ** | 0.40 * | 0.39 * | 0.52 ** | 0.33 * | 0.46 ** | 0.69 ** | 0.59 ** | 0.66 ** | 0.70 ** | 0.63 ** |
| Maple | 0.57 ** | 0.41 * | 0.37 * | 0.54 ** | 0.35 * | 0.53 ** | 0.75 ** | 0.60 ** | 0.72 ** | 0.76 ** | 0.65 ** | |
| SPR (KPa) | Alder | −0.61 ** | −0.53 ** | −0.41 * | −0.84 ** | −0.57 ** | −0.88 ** | −0.89 ** | −0.76 ** | −0.81 ** | −0.85 ** | −0.74 ** |
| Maple | −0.63 ** | −0.59 ** | −0.44 * | −0.88 ** | −0.59 ** | −0.86 ** | −0.90 ** | −0.77 ** | −0.80 ** | −0.86 ** | −0.72 ** | |
| MC (%) | Alder | 0.40 * | 0.40 * | 0.39 * | 0.51 ** | 0.37 * | 0.37 * | 0.43 * | 0.56 ** | 0.59 ** | 0.60 ** | 0.48 ** |
| Maple | 0.41 * | 0.43 * | 0.38 * | 0.49 ** | 0.34 * | 0.41 * | 0.45 * | 0.57 ** | 0.58 ** | 0.61 ** | 0.51 ** | |
| OC (%) | Alder | 0.45 * | 0.40 * | 0.38 * | 0.44 * | 0.37 * | 0.53 ** | 0.59 ** | 0.53 ** | 0.57 ** | 0.62 ** | 0.56 ** |
| Maple | 0.47 * | 0.39 * | 0.39 * | 0.46 * | 0.36 * | 0.55 ** | 0.60 ** | 0.55 ** | 0.60 ** | 0.64 ** | 0.59 ** | |
| N (%) | Alder | 0.14 | 0.10 | 0.22 | 0.12 | 0.19 | 0.37 * | 0.11 | 0.18 | 0.36 * | 0.22 | 0.19 |
| Maple | 0.12 | 0.13 | 0.20 | 0.11 | 0.17 | 0.35 * | 0.14 | 0.15 | 0.35 * | 0.19 | 0.21 | |
| pH | Alder | 0.08 | 0.10 | 0.11 | 0.37 * | 0.10 | 0.33 * | 0.10 | 0.14 | 0.37 * | 0.18 | 0.20 |
| Maple | 0.09 | 0.08 | 0.10 | 0.35 * | 0.13 | 0.39 * | 0.09 | 0.15 | 0.35 * | 0.14 | 0.15 |
| Seedling Architecture Indices | Ratio of MRL to SL | Ratio of LRL to MRL | Ratio of RPD to MRL | Ratio of Root Biomass to Total Biomass |
|---|---|---|---|---|
| Alder seedling | 0.000 ** | 0.342 | 0.000 ** | 0.507 |
| Maple seedling | 0.045 * | 0.359 | 0.000 ** | 0.482 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tavankar, F.; Venanzi, R.; Nikooy, M.; Lo Monaco, A.; Picchio, R.; Naghdi, R. The Influence of Logging-Related Soil Disturbance on Pioneer Tree Regeneration in Mixed Temperate Forests. Plants 2024, 13, 2149. https://doi.org/10.3390/plants13152149
Tavankar F, Venanzi R, Nikooy M, Lo Monaco A, Picchio R, Naghdi R. The Influence of Logging-Related Soil Disturbance on Pioneer Tree Regeneration in Mixed Temperate Forests. Plants. 2024; 13(15):2149. https://doi.org/10.3390/plants13152149
Chicago/Turabian StyleTavankar, Farzam, Rachele Venanzi, Mehrdad Nikooy, Angela Lo Monaco, Rodolfo Picchio, and Ramin Naghdi. 2024. "The Influence of Logging-Related Soil Disturbance on Pioneer Tree Regeneration in Mixed Temperate Forests" Plants 13, no. 15: 2149. https://doi.org/10.3390/plants13152149
APA StyleTavankar, F., Venanzi, R., Nikooy, M., Lo Monaco, A., Picchio, R., & Naghdi, R. (2024). The Influence of Logging-Related Soil Disturbance on Pioneer Tree Regeneration in Mixed Temperate Forests. Plants, 13(15), 2149. https://doi.org/10.3390/plants13152149