

Supplementary Far-Red Light for Photosynthetic Active Radiation Differentially Influences the Photochemical Efficiency and Biomass Accumulation in Greenhouse-Grown Lettuce

Abstract
1. Introduction
2. Results
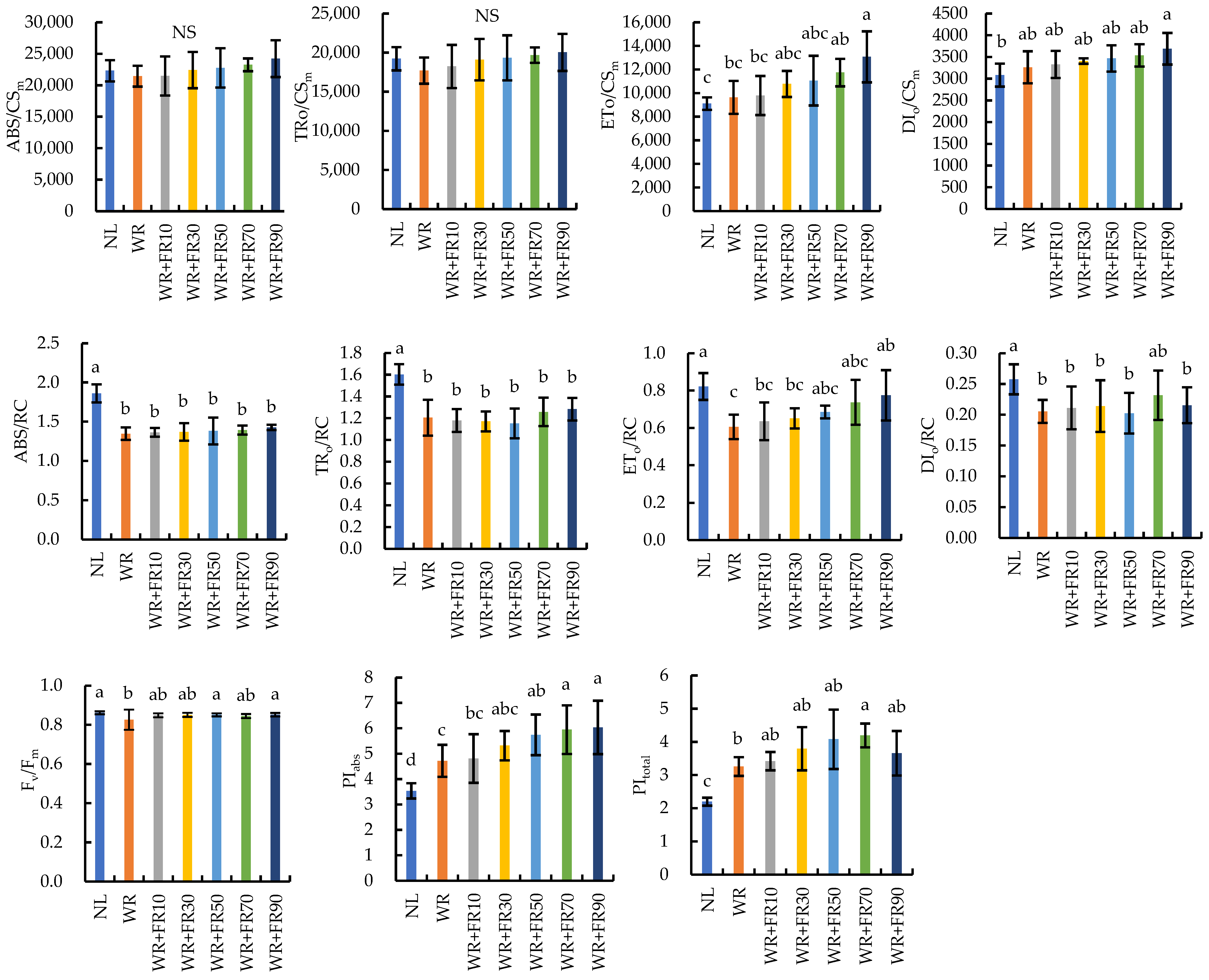
2.1. Chlorophyll: Fluorescence Parameters of Greenhouse-Grown Lettuce
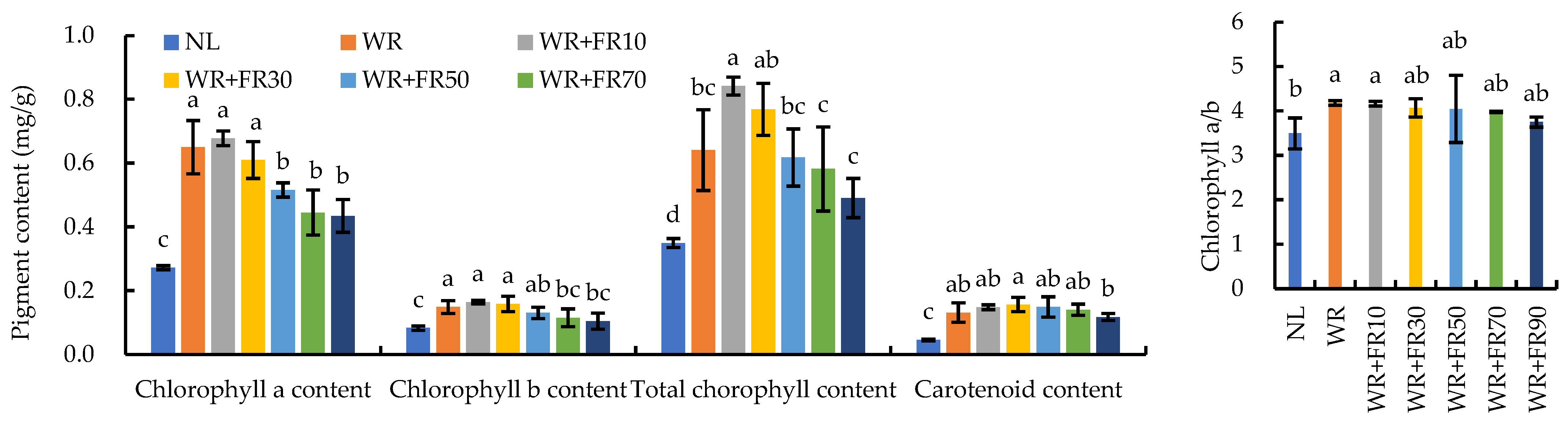
2.2. Photosynthetic Characteristics and Chlorophyll Content of Greenhouse-Grown Lettuce
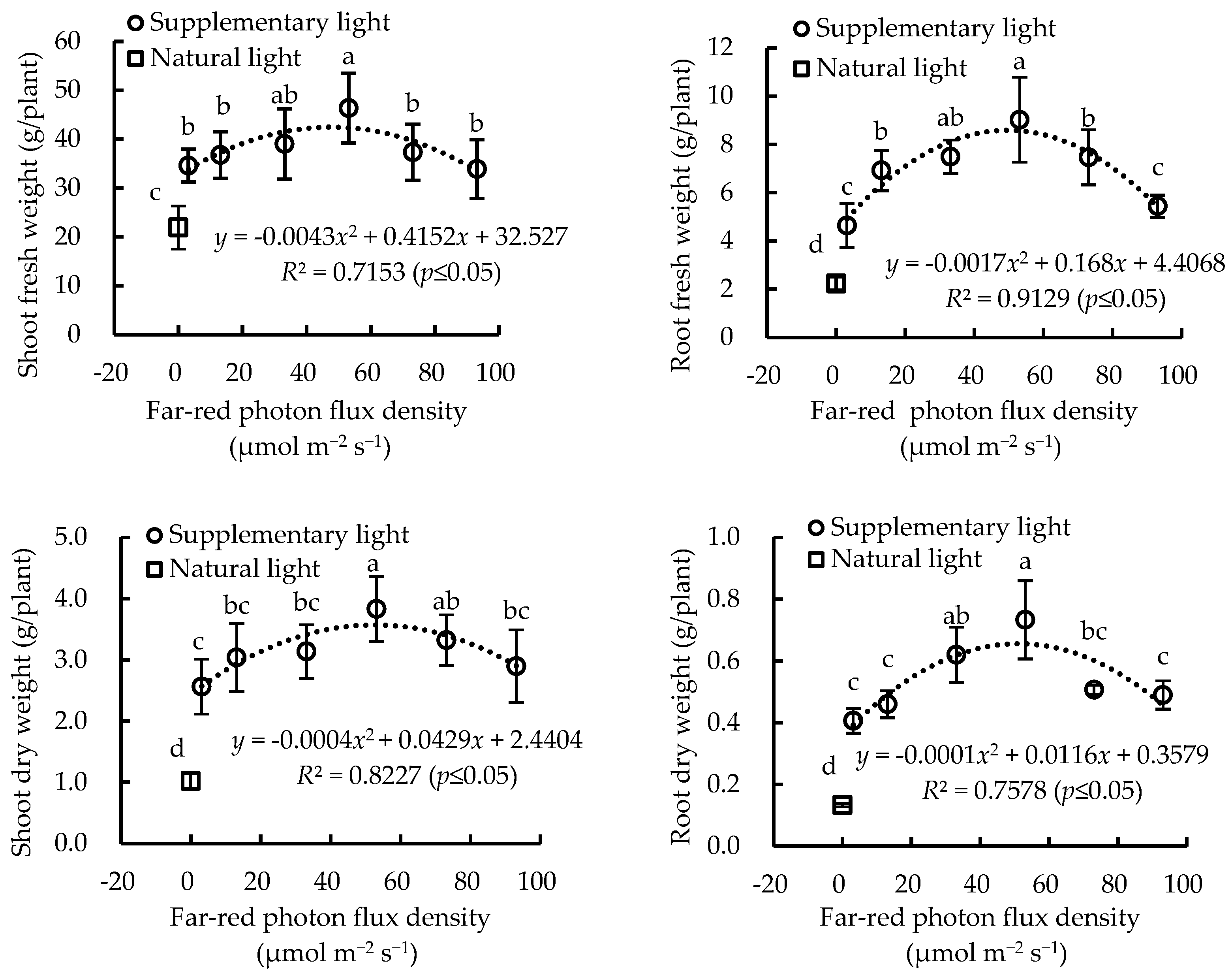
2.3. Growth and Biomass Accumulation of Greenhouse-Grown Lettuce
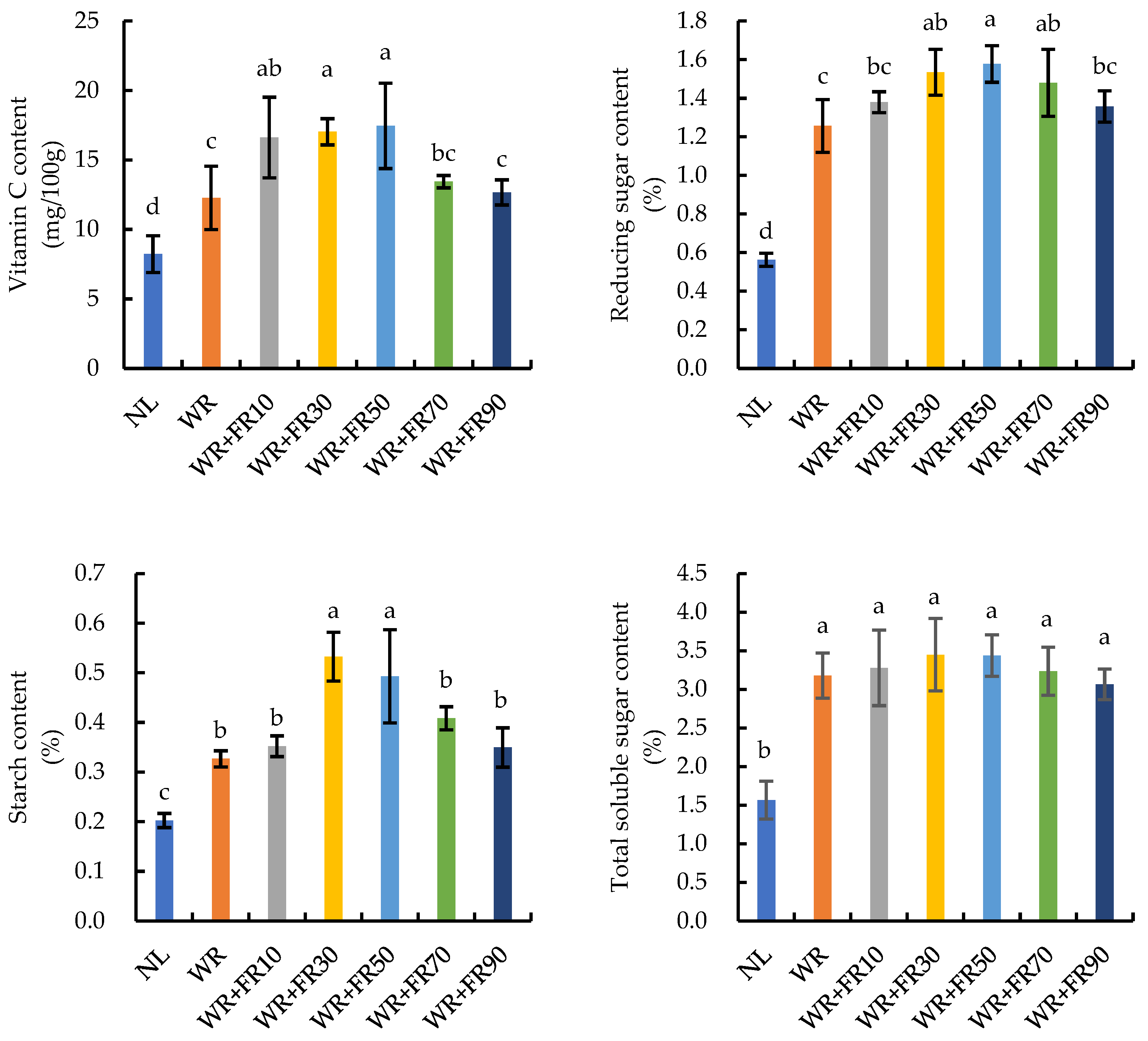
2.4. Vitamin C, Sugar, and Starch Contents of Greenhouse-Grown Lettuce
3. Discussion
4. Materials and Methods
4.1. Plant Materials
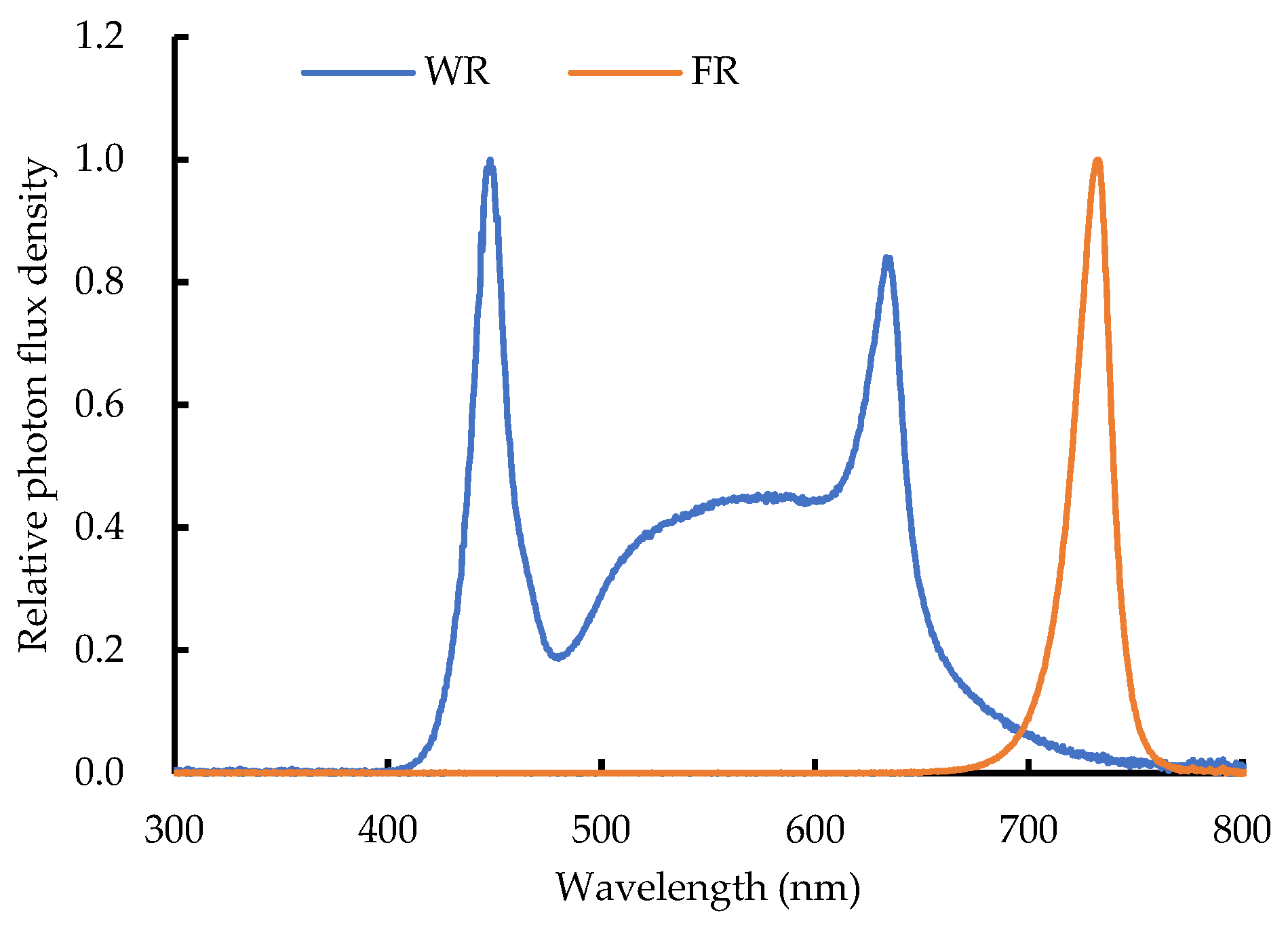
4.2. Light Treatment Design
4.3. Growth Measurement
4.3.1. Photosynthetic Characteristics, Pigments, and Chlorophyll Fluorescence
4.3.2. Plant Morphology and Growth Characteristics
4.3.3. Vitamin C, Reducing Sugar, Soluble Sugar, and Starch Content
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light to blue light. Front Plant. Sci. 2016, 10, 250. [Google Scholar]
- Dou, H.; Niu, G.; Gu, M.; Masabni, J.G. Responses of sweet basil to different daily light integrals in photosynthesis, morphology, yield, and nutritional quality. HortScience 2018, 53, 496–503. [Google Scholar] [CrossRef]
- Pennisi, G.; Orsini, F.; Blasioli, S.; Cellini, A.; Crepaldi, A.; Braschi, I.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Stanghellini, C. Resource use efficiency of indoor lettuce (Lactuca sativa L.) cultivation as affected by red:blue ratio provided by LED lighting. Nat. Sci. Rep. 2019, 9, 14127. [Google Scholar]
- Spalholz, H.; Perkins-Veazie, P.; Hernández, R. Impact of sun-simulated white light and varied blue:red spectrums on the growth, morphology, development, and phytochemical content of green- and red-leaf lettuce at different growth stages. Sci. Hortic. 2020, 264, 109195. [Google Scholar] [CrossRef]
- McCree, K.J. The action spectrum, absorptance and quantum yield of photosynthesis in crop plants. Agric. Meteorol. 1972, 9, 191–216. [Google Scholar] [CrossRef]
- Zou, J.; Fanourakis, D.; Tsaniklidis, G.; Cheng, R.; Yang, Q.; Li, T. Lettuce growth, morphology and critical leaf trait responses to far-red light during cultivation are low fluence and obey the reciprocity law. Sci. Hortic. 2021, 289, 110455. [Google Scholar] [CrossRef]
- Jin, W.; Urbina, J.L.; Heuvelink, E.; Marcelis, L.F.M. Adding far-red to red-blue light-emitting diode light promotes yield of lettuce at different planting densities. Front. Plant Sci. 2021, 11, 609977. [Google Scholar] [CrossRef] [PubMed]
- Zhen, S.; Van Iersel, M.W. Far-red light is needed for efficient photochemistry and photosynthesis. J. Plant Physiol. 2017, 209, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Zhen, S.; Bugbee, B. Far-red photons have equivalent efficiency to traditional photosynthetic photons: Implications for redefining photosynthetically active radiation. Plant Cell Environ. 2020, 43, 1259–1272. [Google Scholar] [CrossRef] [PubMed]
- Legendre, R.; van Iersel, M.W. Supplemental far-red light stimulates lettuce growth: Disentangling morphological and physiological effects. Plants 2021, 10, 166. [Google Scholar] [CrossRef] [PubMed]
- Kusuma, P.; Bugbee, B. On the contrasting morphological response to far-red at high and low photon fluxes. Front. Plant Sci. 2023, 14, 1185622. [Google Scholar] [CrossRef]
- Ji, Y.; Ouzounis, T.; Courbier, S.; Kaiser, E.; Nguyen, P.T.; Schouten, H.J.; Visser, R.G.F.; Pierik, R.; Marcelis, L.F.M.; Heuvelink, E. Far-red radiation increases dry mass partitioning to fruits but reduces Botrytis cinerea resistance in tomato. Environ. Exp. Bot. 2019, 168, 103889. [Google Scholar] [CrossRef]
- Zou, J.; Zhang, Y.; Zhang, Y.; Bian, Z.; Fanourakis, D.; Yang, Q.; Li, T. Morphological and physiological properties of indoor cultivated lettuce in response to additional far-red light. Sci. Hortic. 2019, 257, 108725. [Google Scholar] [CrossRef]
- Wong, C.; Teo, Z.W.N.; Shen, L.; Yu, H. Seeing the lights for leafy greens in indoor vertical farming. Trends Food Sci. Technol. 2020, 106, 48–63. [Google Scholar] [CrossRef]
- Kong, Y.; Nemali, K. Blue and far-red light affect area and number of individual leaves to influence vegetative growth and pigment synthesis in lettuce. Front. Plant Sci. 2021, 12, 667407. [Google Scholar] [CrossRef]
- Zou, J.; Fanourakis, D.; Tsaniklidis, G.; Woltering, E.J.; Cheng, R.; Li, T. Far-red radiation during indoor cultivation reduces lettuce nutraceutical quality and shortens the shelf-life when stored at supra optimal temperatures. Postharvest Biol. Technol. 2023, 198, 112269. [Google Scholar] [CrossRef]
- Gao, H.; Gong, L.; Ni, J.; Li, Q. Metabolomics analysis of lettuce (Lactuca sativa L.) affected by low potassium supply. Agriculture 2022, 12, 1153. [Google Scholar]
- Zhao, J.; Li, H.; Chen, C.; Pang, Y.; Zhu, X. Detection of water content in lettuce canopies based on hyperspectral imaging technology under outdoor conditions. Agriculture 2022, 12, 1796. [Google Scholar] [CrossRef]
- Li, Q.; Gao, H.; Zhang, X.; Ni, J.; Mao, H. Describing lettuce growth using morphological features combined with nonlinear models. Agronomy 2022, 12, 860. [Google Scholar] [CrossRef]
- THECORNERPLOT. Available online: https://thecornerplot.blog/2023/07/28/exploring-the-global-production-of-lettuce-where-does-it-grow/ (accessed on 15 March 2024).
- Paz, M.; Fisher, P.R.; Gómez, C. Minimum light requirements for indoor gardening of lettuce. Urban Agric. Reg. Food Syst. 2019, 4, 1–10. [Google Scholar] [CrossRef]
- Zhang, M.; Whitman, C.M.; Runkle, E.S. Manipulating growth, color, and taste attributes of fresh cut lettuce by greenhouse supplemental lighting. Sci. Hortic. 2019, 252, 274–282. [Google Scholar] [CrossRef]
- Yan, Z.; Wang, C.; Li, Z.; Li, X.; Cheng, F.; Lin, D.; Yang, Y. Supplementary white, UV-A, and far-red radiation differentially regulates growth and nutritional qualities of greenhouse lettuce. Plants 2023, 12, 3234. [Google Scholar] [CrossRef]
- Misra, A.N.; Misra, M.; Singh, R. Chlorophyll fluorescence in plant biology. Biophysics 2012, 7, 171–192. [Google Scholar]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. Probing Photosynth. Mech. Regul. Adapt. 2000, 25, 445–483. [Google Scholar]
- Zhang, C.; Akhlaq, M.; Yan, H.; Ni, Y.; Liang, S.; Zhou, J.; Xue, R.; Li, M.; Adnan, M.R.; Li, J. Chlorophyll fluorescence parameter as a predictor of tomato growth and yield under CO2 enrichment in protective cultivation. Agricul. Water Manag. 2023, 284, 108333. [Google Scholar] [CrossRef]
- Chen, X.; Li, Y.; Wang, L.; Yang, Q.; Guo, W. Responses of butter leaf lettuce to mixed red and blue light with extended light/dark cycle period. Sci. Rep. 2022, 12, 6924. [Google Scholar] [CrossRef] [PubMed]
- Shamsabad, M.R.M.; Esmaeilizadeh, M.; Roosta, H.R.; Dehghani, M.R.; Dąbrowski, P.; Kalaji, H.M. The effect of supplementary light on the photosynthetic apparatus of strawberry plants under salinity and alkalinity stress. Sci. Rep. 2022, 12, 13257. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Ma, C.; Feng, Y.; Zhang, J.; Yang, F.; Li, Y. Response of chlorophyll fluorescence transient in leaves of wheats with different drought resistances to drought stresses and rehydration. Plant Physiol. J. 2018, 54, 1119–1129. [Google Scholar]
- Khudyakova, A.Y.; Kreslavski, V.D.; Shmarev, A.N.; Lyubimov, V.Y.; Shirshikova, G.N.; Pashkovskiy, P.P.; Kuznetsov, V.V.; Allakhverdiev, S.I. Impact of UV-B radiation on the photosystem II activity, pro-/antioxidant balance and expression of light-activated genes in Arabidopsis thaliana hy4 mutants grown under light of different spectral composition. J. Photochem. Photobiol. B Biol. 2019, 194, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Stefanov, D.; Petkova, V.; Denev, I.D. Screening for heat tolerance in common bean (Phaseolus vulgaris L.) lines and cultivars using JIP-test. Sci. Hortic. 2011, 128, 1–6. [Google Scholar]
- Wang, Q.; Ning, Z.; Awan, S.A.; Gao, J.; Chen, J.; Lei, Y.; Tan, X.; Wu, X.; Wu, Y.; Liu, C.; et al. Far-red light mediates light energy capture and distribution in soybeans (Glycine max L.) under the shade. Plant Physiol. Biochem. 2023, 204, 108130. [Google Scholar]
- Kono, M.; Kawaguchi, H.; Mizusawa, N.; Yamori, W.; Suzuki, Y.; Terashima, I. Far-red light accelerates photosynthesis in the low-light phases of fluctuating light. Plant Cell Physiol. 2019, 61, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Zhen, S.; van Iersel, M.W.; Bugbee, B. Photosynthesis in sun and shade: The surprising importance of far-red photons. New Phytol. 2022, 236, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, Z.; Li, Y.; Lu, X.; Li, G.; Qi, S.; Khan, I.; Li, G.; Dai, Z.; Du, D. The degradability of microplastics may not necessarily equate to environmental friendliness: A case study of cucumber seedlings with disturbed photosynthesis. Agriculture 2024, 14, 53. [Google Scholar] [CrossRef]
- Franklin, K.A. Shade avoidance. New Phytol. 2010, 179, 930–944. [Google Scholar] [CrossRef]
- Park, Y.; Runkle, E.S. Far-red radiation promotes growth of seedlings by increasing leaf expansion and whole-plant net assimilation. Environ Exp. Bot. 2017, 136, 41–49. [Google Scholar] [CrossRef]
- Meng, Q.; Kelly, N.; Runkle, E. Substituting green or far-red radiation for blue radiation induces shade avoidance and promotes growth in lettuce and kale. Environ. Exp. Bot. 2019, 162, 383–391. [Google Scholar] [CrossRef]
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Meng, Q.; Runkle, E. Far red radiation interacts with relative and absolute blue and red photon flux densities to regulate growth, morphology, and pigmentation of lettuce and basil seedlings. Sci. Hortic. 2019, 255, 269–280. [Google Scholar] [CrossRef]
- Elkins, C.; van Iersel, M.W. Longer photoperiods with the same daily light integral improve growth of Rudbeckia seedlings in a greenhouse. HortScience 2020, 55, 1676–1682. [Google Scholar] [CrossRef]
- Weaver, G.; van Iersel, M.W. Longer photoperiods with adaptive lighting control can improve growth of greenhouse-grown ‘Little Gem’ lettuce (Lactuca sativa). HortScience 2020, 55, 573–580. [Google Scholar] [CrossRef]
- Van de Velde, E.; Steppe, K.; Van Labeke, M.C. Leaf morphology, optical characteristics and phytochemical traits of butterhead lettuce affected by increasing the far-red photon flux. Front. Plant Sci. 2023, 14, 1129335. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; van Iersel, M.W. Far-red light effects on lettuce growth and morphology in indoor production are cultivar specific. Plants 2022, 11, 2714. [Google Scholar] [CrossRef]
- Trupkin, S.A.; Legris, M.; Buchovsky, A.S.; Rivero, M.B.T.; Casal, J.J. Phytochrome B nuclear bodies respond to the low red to far-red ratio and to the reduced irradiance of canopy shade in Arabidopsis. Plant Physiol. 2014, 165, 1698–1708. [Google Scholar] [CrossRef]
- Shmarev, A.; Vereshagin, M.; Pashkovskiy, P.; Kreslavski, V.D.; Allakhverdiev, S.I. Influence of additional far-red light on photosynthetic and growth parameters of lettuce plants and the resistance of the photosynthetic apparatus to high irradiance. Photosynthetica 2024, 62, 180–186. [Google Scholar] [CrossRef]
- Cao, K.; Yu, J.; Xu, D.; Ai, K.; Bao, E.; Zou, Z. Exposure to lower red to far-red light ratios improve tomato tolerance to salt stress. BMC Plant Biol. 2018, 18, 92. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.G.; Smith, H. The function of phytochrome in the natural environment. I. Characterization of daylight for studies in photomorphogenesis and photoperiodism. Photochem. Photobiol. 1977, 25, 533–538. [Google Scholar] [CrossRef]
- Yang, F.; Liu, Q.; Cheng, Y.; Feng, L.; Wu, X.; Fan, Y.; Raza, M.A.; Wang, X.; Yong, T.; Liu, W.; et al. Low red/far-red ratio as a signal promotes carbon assimilation of soybean seedlings by increasing the photosynthetic capacity. BMC Plant Biol. 2020, 20, 148. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Huang, M.; Huang, W.; Hsu, M.H.; Yang, C. The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar]
- Woltering, E.J.; Witkowska, I.M. Effects of pre- and postharvest lighting on quality and shelf life of fresh-cut lettuce. Acta. Hortic. 2016, 1134, 357–366. [Google Scholar] [CrossRef]
- Ballaré, C.L. Light regulation of plant defense. Annu. Rev. Plant Biol. 2014, 65, 335–363. [Google Scholar] [CrossRef] [PubMed]
- Mickens, M.A.; Skoog, E.J.; Reese, L.E.; Barnwell, P.L.; Spencer, L.E.; Massa, G.D.; Wheeler, R.M. A strategic approach for investigating light recipes for ‘Outredgeous’ red romaine lettuce using white and monochromatic LEDs. Life Sci. Space Res. 2018, 19, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, T.; Endo, R.; Hayashi, N.; Kitaya, Y. High-light-like photosynthetic responses of Cucumis sativus leaves acclimated to fluorescent illumination with a high red:far-red ratio: Interaction between light quality and quantity. Photosynthetic 2012, 50, 623–629. [Google Scholar] [CrossRef]
- Yan, Z.; He, D.; Niu, G.; Zhou, Q.; Qu, Y. Growth, nutritional quality and energy use efficiency of hydroponic lettuce as influenced by daily light integrals exposed to white versus white plus red light-emitting diodes. HortScience 2019, 54, 1737–1744. [Google Scholar] [CrossRef]
- Ye, Z.; Suggett, D.; Robakowski, P.; Kang, H. A mechanistic model for the photosynthesis light response based on the photosynthetic electron transport of photosystem II in C3 and C4 species. New Phytol. 2013, 199, 110–120. [Google Scholar] [CrossRef]
- Li, Q.; Chen, L.; Jiang, H.; Tang, N.; Yang, L.; Lin, Z.; Li, Y. Effects of manganese-excess on CO2 assimilation, ribulose-1,5-bisphosphate carboxylase/oxygenase, carbohydrates and photosynthetic electron transport of leaves, and antioxidant systems of leaves and roots in Citrus grandis seedlings. BMC. Plant Biol. 2010, 10, 42. [Google Scholar] [CrossRef]
- Klinkovsky, T.; Naus, J. Sensitivity of the relative Fpl level of chlorophyll fluorescence induction in leaves to the heat stress. Photosynth. Res. 1994, 39, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Wellburn, A. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 603, 591–592. [Google Scholar] [CrossRef]
- Shyamala, B.J.; Jamuna, P. Nutritional content and antioxidant properties of pulp waste from Daucus carota and Beta vulgaris. Malays. J. Nutr. 2010, 16, 397–408. [Google Scholar] [PubMed]
- Song, J.; Huang, H.; Hao, Y.; Song, S.; Liu, H. Nutritional quality, mineral and antioxidant content in lettuce affected by interaction of light intensity and nutrient solution concentration. Sci. Rep. 2020, 10, 2796. [Google Scholar] [CrossRef]
- Zhan, L.; Hu, J.; Ai, Z.; Pang, L.; Li, Y.; Zhu, M. Light exposure during storage preserving soluble sugar and l-ascorbic acid content of minimally processed romaine lettuce (Lactuca sativa L.var. longifolia). Food Chem. 2012, 136, 273–278. [Google Scholar]
- Takahashi, K.; Fujino, K.; Kikuta, Y.; Koda, Y. Involvement of the accumulation of sucrose and the synthesis of cell wall polysaccharides in the expansion of potato cells in response to jasmonic acid. Plant Sci. 1995, 111, 11–18. [Google Scholar] [CrossRef]

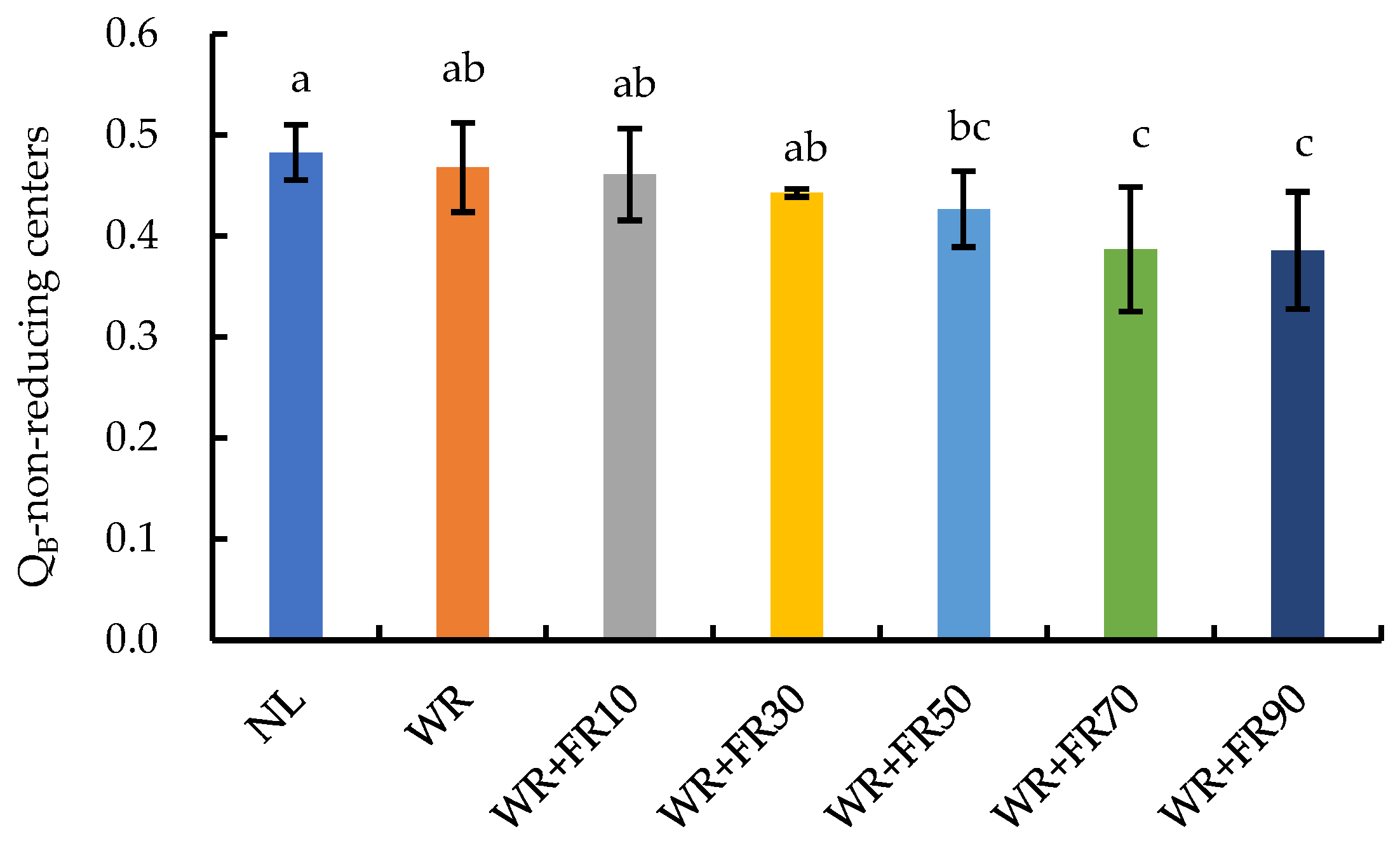
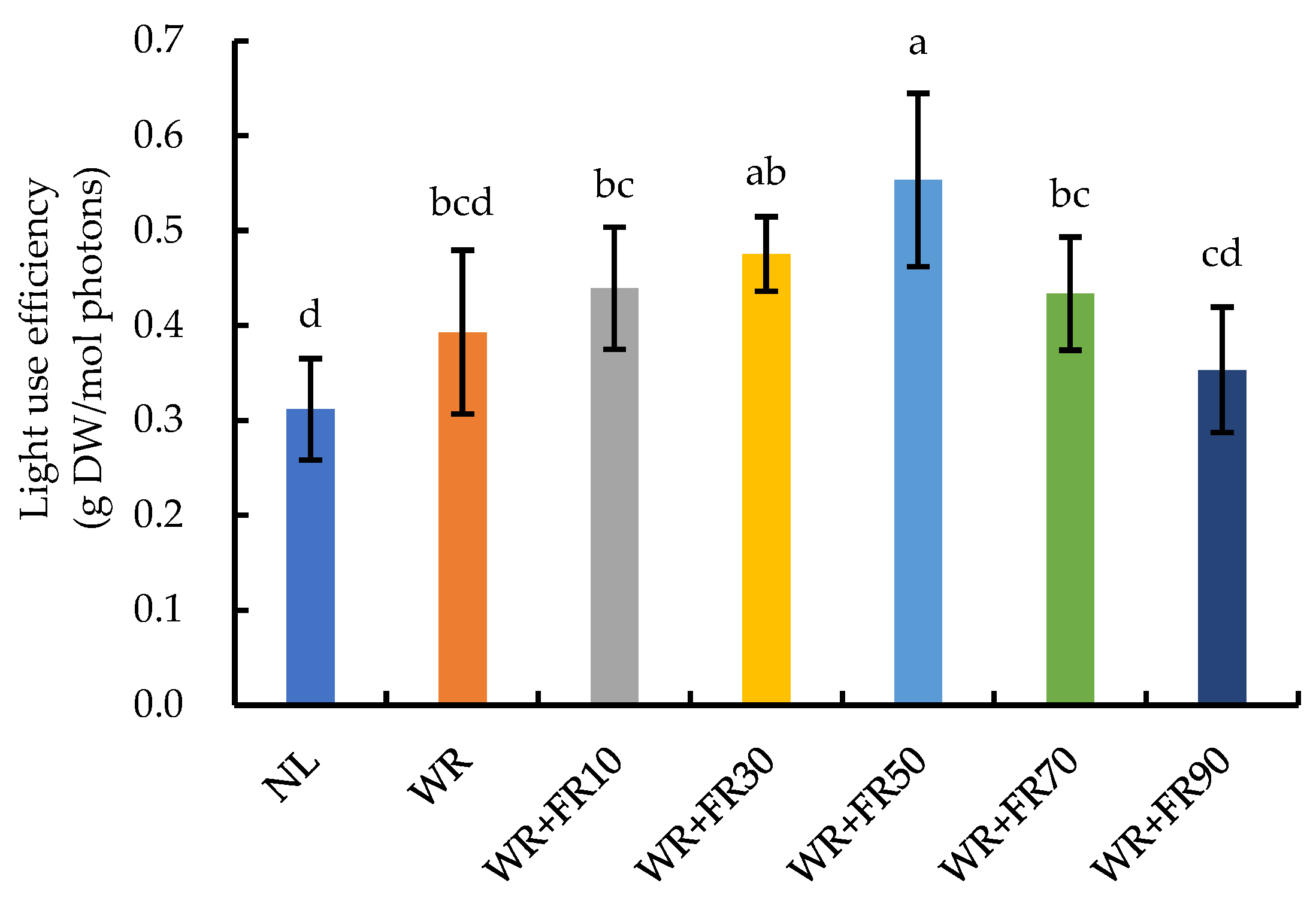

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Pn max (μmol m−2 s−1) | Rd (μmol m−2 s−1) | Lc (μmol m−2 s−1) | |||
|---|---|---|---|---|---|---|
| NL | 6.5 ± 1.1 | c | 0.84 ± 0.09 | b | 12.6 ± 2.3 | c |
| WR | 10.2 ± 0.9 | ab | 0.88 ± 0.16 | b | 12.7 ± 1.8 | c |
| WR + FR10 | 10.7 ± 2.0 | a | 1.01 ± 0.05 | b | 17.2 ± 3.1 | b |
| WR + FR30 | 11.6 ± 2.6 | a | 1.73 ± 0.34 | a | 20.1 ± 4.5 | ab |
| WR + FR50 | 11.7 ± 1.5 | a | 1.53 ± 0.14 | a | 21.3 ± 1.3 | a |
| WR + FR70 | 10.5 ± 0.1 | ab | 1.09 ± 0.14 | b | 16.5 ± 1.7 | bc |
| WR + FR90 | 8.0 ± 1.3 | bc | 1.01 ± 0.18 | b | 16.1 ± 2.9 | bc |
| Treatments | Leaf Number | Leaf Length (cm) | Leaf Width (cm) | Length-Width Ratio | Leaf Area (cm2) | Specific Leaf Area (cm2 g−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NL | 9.0 ± 0.7 | c | 16.8 ± 0.8 | a | 12.0 ± 0.6 | c | 1.40 ± 0.09 | a | 561 ± 76 | d | 1121 ± 97 | a |
| WR | 11.0 ± 1.0 | b | 14.2 ± 0.2 | b | 14.8 ± 1.4 | b | 0.97 ± 0.09 | b | 678 ± 131 | cd | 506 ± 36 | b |
| WR + FR10 | 11.4 ± 0.5 | b | 14.0 ± 0.6 | bc | 15.0 ± 1.1 | b | 0.94 ± 0.09 | bc | 745 ± 40 | c | 469 ± 23 | b |
| WR + FR30 | 11.8 ± 0.8 | b | 12.8 ± 0.4 | d | 15.2 ± 0.6 | ab | 0.85 ± 0.06 | cd | 945 ± 73 | ab | 445 ± 33 | b |
| WR + FR50 | 13.0 ± 1.2 | a | 12.6 ± 1.1 | d | 16.4 ± 0.5 | a | 0.77 ± 0.07 | d | 1031 ± 138 | a | 448 ± 45 | b |
| WR + FR70 | 11.8 ± 0.8 | b | 13.2 ± 0.8 | cd | 14.8 ± 1.3 | b | 0.90 ± 0.11 | bc | 811 ± 89 | bc | 500 ± 17 | b |
| WR + FR90 | 11.4 ± 0.5 | b | 13.3 ± 0.8 | bcd | 14.2 ± 1.2 | b | 0.95 ± 0.07 | bc | 730 ± 37 | c | 507 ± 39 | b |
| Treatments | PPFD (µmol m−2 s−1) | Ratio of B:G:R | FR Photon Flux Density (µmol m−2 s−1) | Ratio of R:FR |
|---|---|---|---|---|
| WR | 151 | 1:1.36:1.14 | 3 | 15.72 |
| WR + FR10 | 13 | 3.75 | ||
| WR + FR30 | 33 | 1.49 | ||
| WR + FR50 | 53 | 0.93 | ||
| WR + FR70 | 73 | 0.67 | ||
| WR + FR90 | 93 | 0.53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dou, H.; Li, X.; Li, Z.; Song, J.; Yang, Y.; Yan, Z. Supplementary Far-Red Light for Photosynthetic Active Radiation Differentially Influences the Photochemical Efficiency and Biomass Accumulation in Greenhouse-Grown Lettuce. Plants 2024, 13, 2169. https://doi.org/10.3390/plants13152169
Dou H, Li X, Li Z, Song J, Yang Y, Yan Z. Supplementary Far-Red Light for Photosynthetic Active Radiation Differentially Influences the Photochemical Efficiency and Biomass Accumulation in Greenhouse-Grown Lettuce. Plants. 2024; 13(15):2169. https://doi.org/10.3390/plants13152169
Chicago/Turabian StyleDou, Haijie, Xin Li, Zhixin Li, Jinxiu Song, Yanjie Yang, and Zhengnan Yan. 2024. "Supplementary Far-Red Light for Photosynthetic Active Radiation Differentially Influences the Photochemical Efficiency and Biomass Accumulation in Greenhouse-Grown Lettuce" Plants 13, no. 15: 2169. https://doi.org/10.3390/plants13152169
APA StyleDou, H., Li, X., Li, Z., Song, J., Yang, Y., & Yan, Z. (2024). Supplementary Far-Red Light for Photosynthetic Active Radiation Differentially Influences the Photochemical Efficiency and Biomass Accumulation in Greenhouse-Grown Lettuce. Plants, 13(15), 2169. https://doi.org/10.3390/plants13152169