


Adaptation of the Invasive Plant Sphagneticola trilobata (L.) Pruski to Drought Stress


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Phenotypic Characteristics
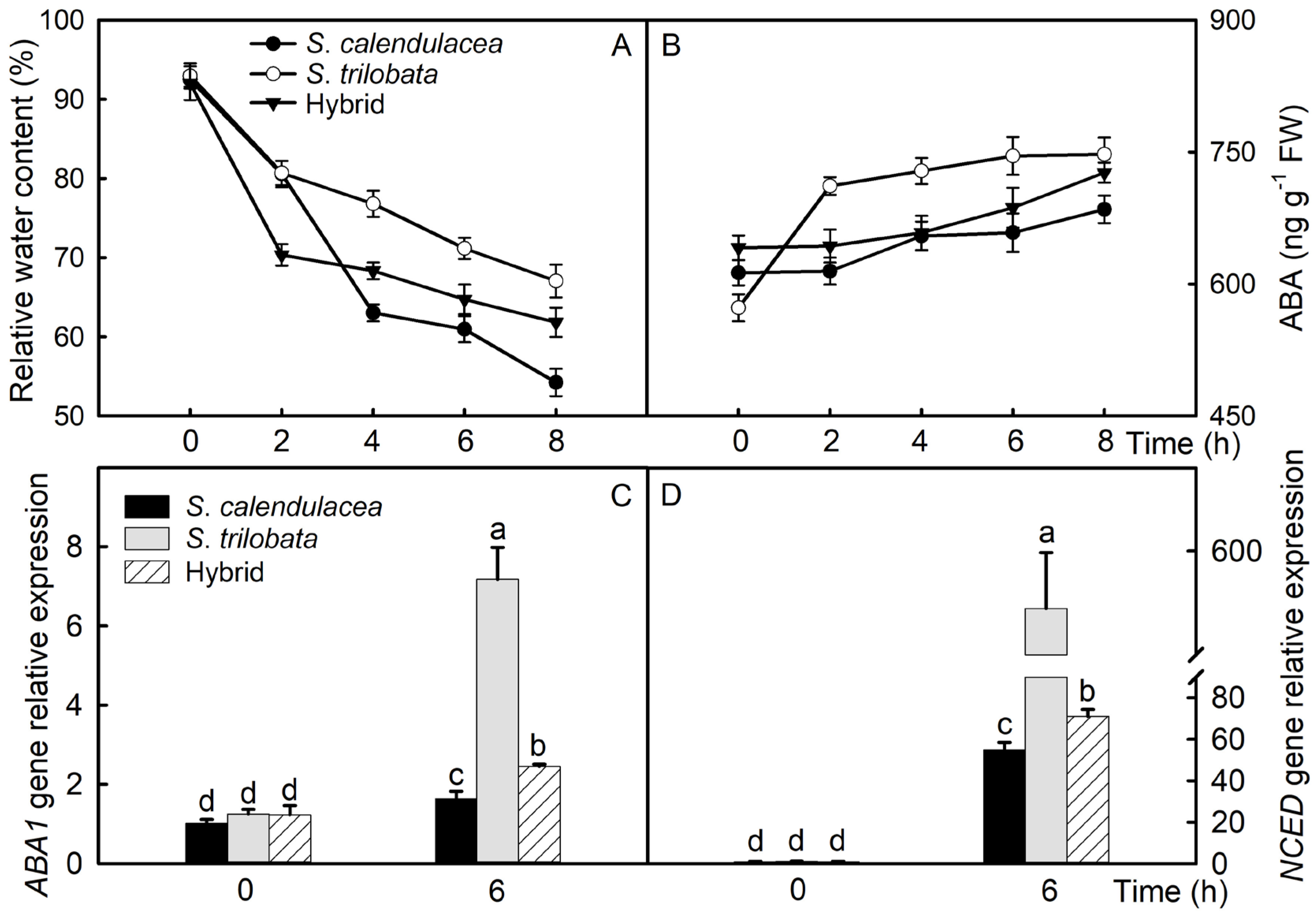
2.2. Leaf Water and ABA Content, as Well as Expression of Related Genes
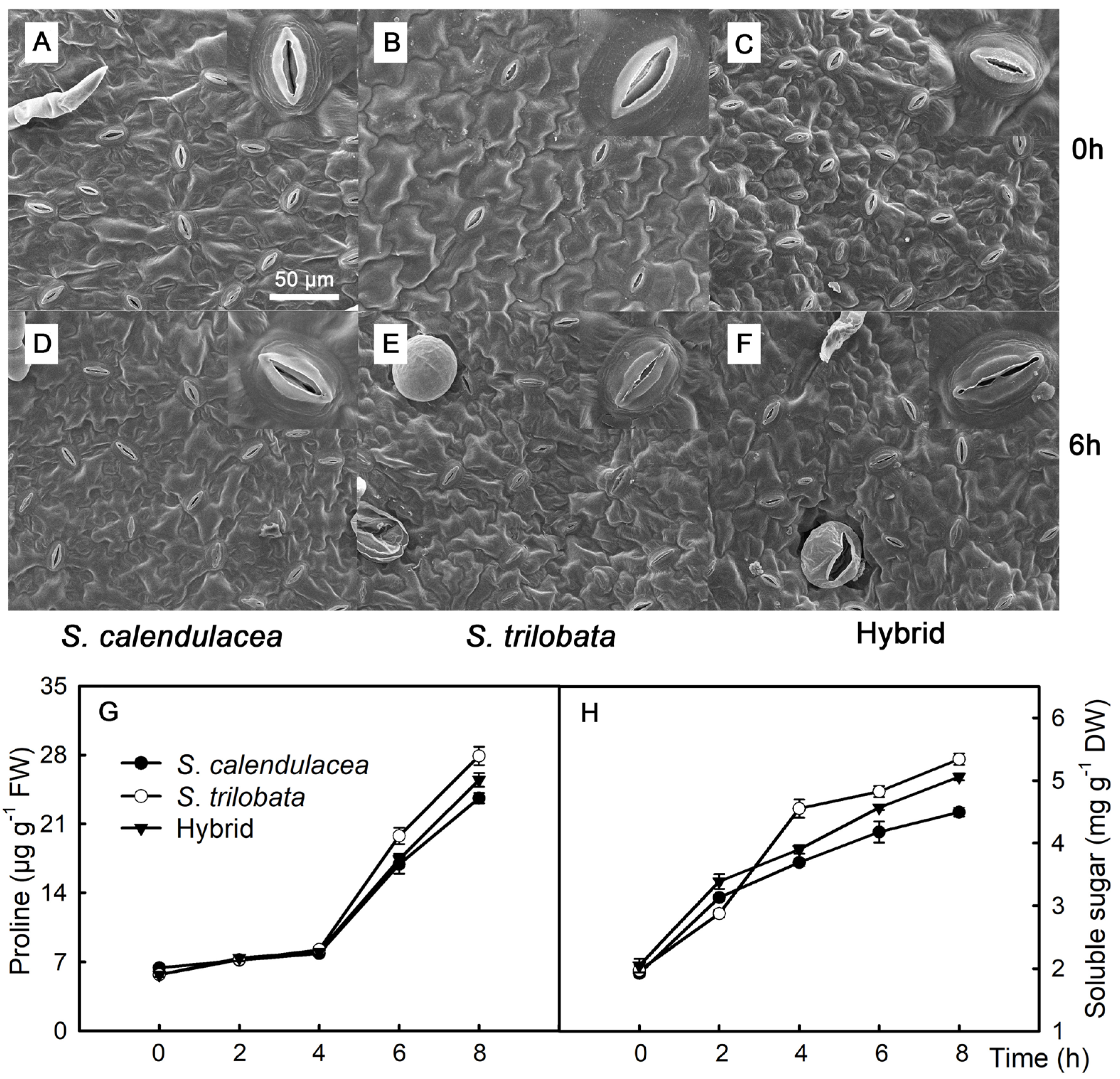
2.3. Leaf Stomata, Proline, and Soluble Sugar
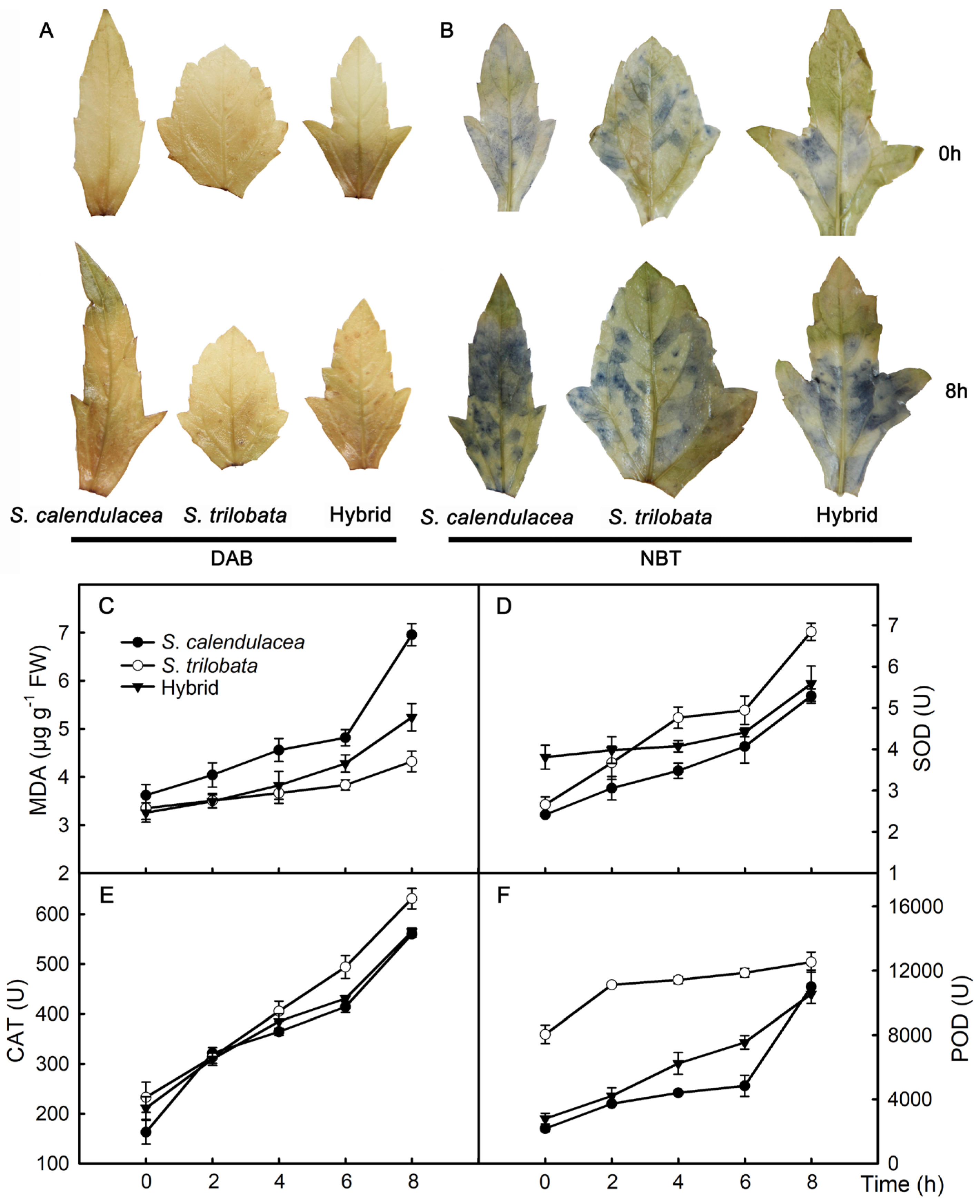
2.4. Hydrogen Peroxide, Superoxide Anion, MDA, and Enzyme Activity
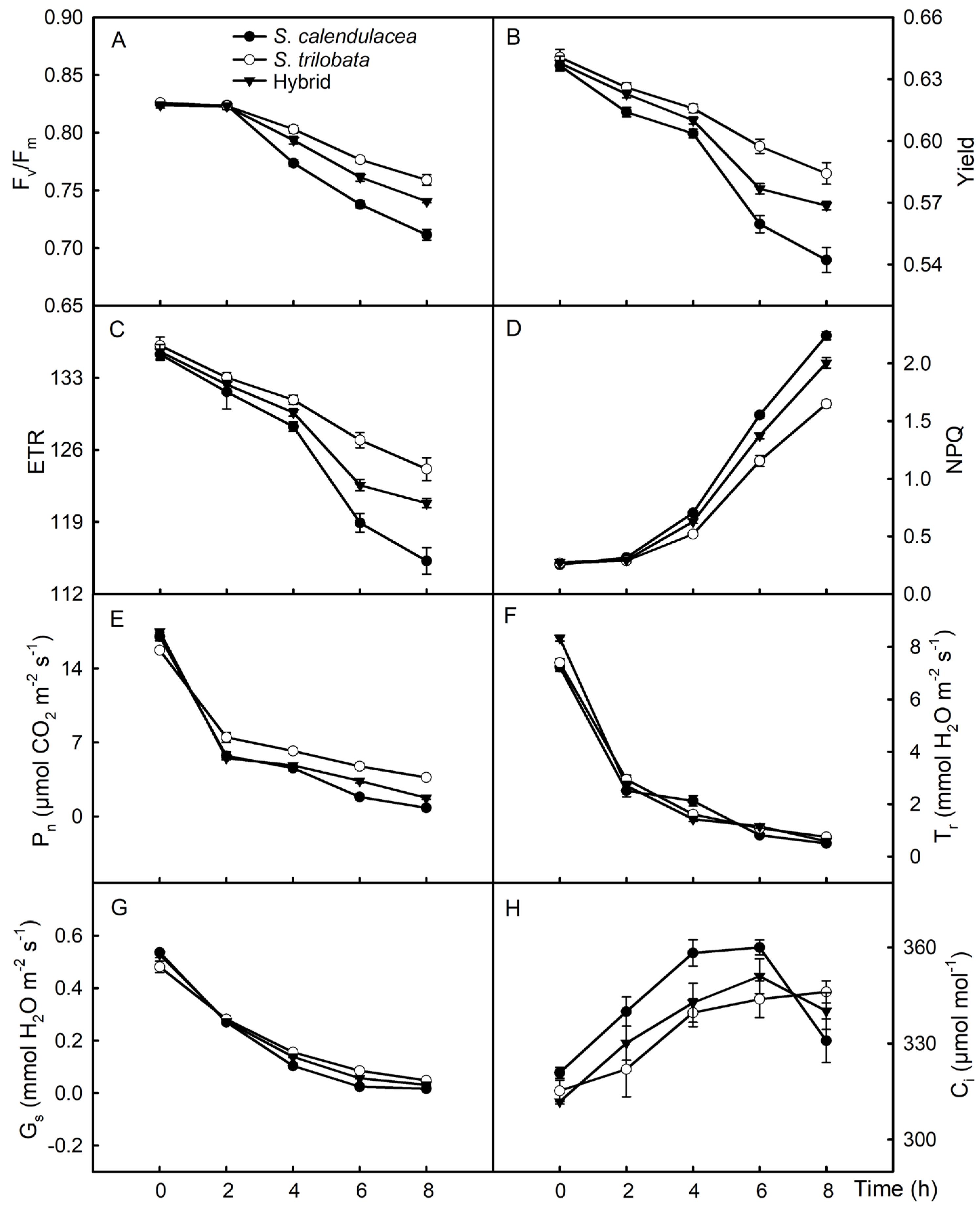
2.5. Chlorophyll Fluorescence Parameters and Gas Exchange Parameters
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Relative Water Content of Leaves
4.3. Abscisic Acid, Proline, and Soluble Sugar
4.4. Enzyme Activity
4.5. Malondialdehyde, Hydrogen Peroxide, and Superoxide Anion
4.6. Gas Exchange and Fluorescence Parameters
4.7. Stomatal Observations
4.8. Gene Expression Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gurvich, D.E.; Tecco, P.A.; Díaz, S. Plant invasions in undisturbed ecosystems: The triggering attribute approach. J. Veg. Sci. 2005, 16, 723–728. [Google Scholar] [CrossRef]
- Daleo, P.; Alberti, J.; Iribarne, O. Biological invasions and the neutral theory. Divers. Distrib. 2009, 15, 547–553. [Google Scholar] [CrossRef]
- Kueffer, C.; Pysek, P.; Richardson, D.M. Integrative invasion science: Model systems, multi-site studies, focused meta-analysis and invasion syndromes. New Phytol. 2013, 200, 615–633. [Google Scholar] [CrossRef]
- Mallet, J. Hybridization as an invasion of the genome. Trends Ecol. Evol. 2005, 20, 229–237. [Google Scholar] [CrossRef]
- Schierenbeck, K.A.; Ellstrand, N.C. Hybridization and the evolution of invasiveness in plants and other organisms. Biol. Invasions 2009, 11, 1093–1105. [Google Scholar] [CrossRef]
- Zhang, Q.L.; Chen, G.X.; Shao, L.; Gu, X.Q.; Huang, J.D.; Peng, C.L. The hybridization between Sphagneticola trilobata (L.) Pruski and Sphagneticola calendulacea (L.) Pruski improved the tolerance of hybrid to cadmium stress. Chemosphere 2020, 249, 126540. [Google Scholar] [CrossRef]
- Mooney, H.A.; Cleland, E.E. The evolutionary impact of invasive species. Proc. Natl. Acad. Sci. USA 2001, 98, 5446–5451. [Google Scholar] [CrossRef]
- Yi, C.X.; Wei, S.H.; Hendrey, G. Warming climate extends dryness-controlled areas of terrestrial carbon sequestration. Sci. Rep. 2014, 4, 5472. [Google Scholar] [CrossRef]
- Abreha, K.B.; Enyew, M.; Carlsson, A.S.; Vetukuri, R.R.; Feyissa, T.; Motlhaodi, T.; Ng’uni, D.; Geleta, M. Sorghum in dryland: Morphological, physiological, and molecular responses of sorghum under drought stress. Planta 2022, 255, 20. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Richards, C.L.; Valladares, F.; et al. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000Research 2016, 5, 1554. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhou, G.; Shimizu, H. Plant responses to drought and rewatering. Plant Signal. Behav. 2010, 5, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Li, Y.; Yin, Z.G.; Jiang, J.H.; Zhang, M.H.; Guo, X.; Ye, Z.J.; Zhao, Y.; Xiong, H.Y.; Zhang, Z.Y.; et al. OsASR5 enhances drought tolerance through a stomatal closure pathway associated with ABA and H2O2 signalling in rice. Plant Biotechnol. J. 2017, 15, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Murata, N.; Allakhverdiev, S.I.; Nishiyama, Y. The mechanism of photoinhibition: Re-evaluation of the roles of catalase, α-tocopherol, non-photochemical quenching, and electron transport. Bba-Bioenergetics 2012, 1817, 1127–1133. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.J.; Zheng, J.; Yu, Z.C.; Gu, X.Q.; Tian, X.S.; Peng, C.L.; Chow, W.S. Variations in photoprotective potential along gradients of leaf development and plant succession in subtropical forests under contrasting irradiances. Environ. Exp. Bot. 2018, 154, 23–32. [Google Scholar] [CrossRef]
- Xu, C.B.; Li, X.M.; Zhang, L.H. The Effect of calcium chloride on growth, photosynthesis, and antioxidant responses of under drought conditions. PLoS ONE 2013, 8, e68214. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, H.; Rashid, U.; Hassan, M.N.; Nosheen, A.; Naz, R.; Ilyas, N.; Sajjad, M.; Azmat, A.; Alyemeni, M.N. Volatile organic compounds produced by alleviated drought stress by modulating defense system in Maize (Zea mays L.). Physiol. Plant. 2021, 172, 896–911. [Google Scholar] [CrossRef] [PubMed]
- Zarrinabadi, I.G.; Razmjoo, J.; Mashhadi, A.A.; Mojeni, H.K.; Boroomand, A. Physiological response and productivity of pot marigold (Calendula officinalis) genotypes under water deficit. Ind. Crop. Prod. 2019, 139, 111488. [Google Scholar] [CrossRef]
- Gao, J.M.; Xiao, Q.; Ding, L.P.; Chen, M.J.; Yin, L.; Li, J.Z.; Zhou, S.Y.; He, G.Y. Differential responses of lipid peroxidation and antioxidants in and subjected to drought stress. Plant Growth Regul. 2008, 56, 89–95. [Google Scholar] [CrossRef]
- Wu, W.; Zhou, R.C.; Ni, G.Y.; Shen, H.; Ge, X.J. Is a new invasive herb emerging? Molecular confirmation and preliminary evaluation of natural hybridization between the invasive Sphagneticola trilobata (Asteraceae) and its native congener S. calendulacea in South China. Biol. Invasions 2013, 15, 75–88. [Google Scholar] [CrossRef]
- Ni, G.Y.; Zhao, P.; Wu, W.; Lu, X.K.; Zhao, X.H.; Zhu, L.W.; Niu, J.F. A hybrid of the invasive plant has similar competitive ability but different response to nitrogen deposition compared to parent. Ecol. Res. 2014, 29, 331–339. [Google Scholar] [CrossRef]
- Sun, Z.Y.; Chen, Y.Q.; Schaefer, V.; Liang, H.M.; Li, W.H.; Huang, S.Q.; Peng, C.L. Responses of the hybrid between Sphagneticola trilobata and Sphagneticola calendulacea to low temperature and weak light characteristic in South China. Sci. Rep. 2015, 5, 16906. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.L.; Chen, G.X.; Ke, W.Q.; Peng, C.L. Adaptation of the invasive plant Sphagneticola trilobata to flooding stress by hybridization with native relatives. Int. J. Mol. Sci. 2024, 25, 6738. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Xu, Z.; He, W.; Yang, H.; Li, B.; Ding, W.; Lei, Y.; Abbas, A.; Hameed, R.; Wang, C.; et al. The cooperation regulation of antioxidative systemand hormone contents on physiological responses of Wedelia trilobata and Wedelia chinensis under simulated drought environment. Plants 2024, 13, 472. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, T.; Zhang, J.; Ma, Y.L.; Li, Y.S.; Zhang, F.; Zhang, Y.; Liang, Y. Overexpression of a mitogen-activated protein kinase SlMAPK3 positively regulates tomato tolerance to cadmium and drought stress. Molecules 2019, 24, 556. [Google Scholar] [CrossRef] [PubMed]
- Kosar, F.; Alshallash, K.S.; Akram, N.A.; Sadiq, M.; Ashraf, M.; Alkhalifah, D.H.M.; Latef, A.A.H.A.; Elkelish, A. Trehalose-induced regulations in nutrient status and secondary metabolites of drought-stressed sunflower (Helianthus annuus L.) plants. Plants 2022, 11, 2780. [Google Scholar] [CrossRef]
- Song, L.Y.; Chow, W.S.; Sun, L.L.; Li, C.H.; Peng, C.L. Acclimation of photosystem II to high temperature in two Wedelia species from different geographical origins: Implications for biological invasions upon global warming. J. Exp. Bot. 2010, 61, 4087–4096. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Shen, F.Y.; Abbas, A.; Wang, H.; Du, Y.Z.; Du, D.L.; Hussain, S.; Javed, T.; Alamri, S. Effects of different nitrogen forms and competitive treatments on the growth and antioxidant system of Wedelia trilobata and Wedelia chinensis under high nitrogen concentrations. Front. Plant Sci. 2022, 13, 851099. [Google Scholar] [CrossRef]
- Guadagno, C.R.; Ewers, B.E.; Speckman, H.N.; Aston, T.L.; Huhn, B.J.; DeVore, S.B.; Ladwig, J.T.; Strawn, R.N.; Weinig, C. Dead or alive? Using membrane failure and chlorophyll a fluorescence to predict plant mortality from drought. Plant Physiol. 2017, 175, 223–234. [Google Scholar] [CrossRef]
- Sun, L.Y.; Li, X.N.; Wang, Z.S.; Sun, Z.W.; Zhu, X.C.; Liu, S.Q.; Song, F.B.; Liu, F.L.; Wang, Y.J. Cold priming induced tolerance to subsequent low temperature stress is enhanced by melatonin application during recovery in wheat. Molecules 2018, 23, 1091. [Google Scholar] [CrossRef]
- Zhang, Q.L.; Chen, G.X.; Huang, J.D.; Peng, C.L. Comparison of the ability to control water loss in the detached leaves of Wedelia trilobata, Wedelia chinensis, and their hybrid. Plants 2020, 9, 1227. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Complex plant responses to drought and heat stress under climate change. Plant J. 2024, 117, 1873–1892. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, G.; Faried, H.N.; Razzaq, K.; Ullah, S.; Wattoo, F.M.; Shehzad, M.A.; Sajjad, Y.; Ahsan, M.; Javed, T.; Dessoky, E.S.; et al. Chitosan-induced physiological and biochemical regulations confer drought tolerance in pot marigold (Calendula officinalis L.). Agronomy 2022, 12, 474. [Google Scholar] [CrossRef]
- Cominelli, E.; Galbiati, M.; Vavasseur, A.; Conti, L.; Sala, T.; Vuylsteke, M.; Leonhardt, N.; Dellaporta, S.L.; Tonelli, C. A guard-cell-specific MYB transcription factor regulates stomatal movements and plant drought tolerance. Curr. Biol. 2005, 15, 1196–1200. [Google Scholar] [CrossRef]
- Bauer, H.; Ache, P.; Lautner, S.; Fromm, J.; Hartung, W.; Al-Rasheid, K.A.S.; Sonnewald, S.; Sonnewald, U.; Kneitz, S.; Lachmann, N.; et al. The stomatal response to reduced relative humidity requires guard cell-autonomous ABA synthesis. Curr. Biol. 2013, 23, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Scuffi, D.; Alvarez, C.; Laspina, N.; Gotor, C.; Lamattina, L.; García-Mata, C. Hydrogen sulfide generated by L-cysteine desulfhydrase acts upstream of nitric oxide to modulate ABA-dependent stomatal closure. Plant Physiol. 2014, 166, 2065–2076. [Google Scholar] [CrossRef]
- Mircea, D.M.; Calone, R.; Shakya, R.; Saavedra, M.F.; Sestras, R.E.; Boscaiu, M.; Sestras, A.F.; Vicente, O. Use of multivariate analysis in screening for drought tolerance in ornamental Asteraceae species. Agronomy 2023, 13, 687. [Google Scholar] [CrossRef]
- Reddy, P.S.; Jogeswar, G.; Rasineni, G.K.; Maheswari, M.; Reddy, A.R.; Varshney, R.K.; Kishor, P.B.K. Proline over-accumulation alleviates salt stress and protects photosynthetic and antioxidant enzyme activities in transgenic sorghum [Sorghum bicolor (L.) Moench]. Plant Physiol. Biochem. 2015, 94, 104–113. [Google Scholar] [CrossRef]
- Zhang, Q.L.; Liu, X.J.; Xu, D.P.; Hong, Z.; Zhang, N.N.; Cui, Z.Y. Effects of drought and host on the growth of seedlings in pot culture. Int. J. Mol. Sci. 2022, 23, 11241. [Google Scholar] [CrossRef]
- Cai, M.; Lin, X.; Peng, J.; Zhang, J.; Chen, M.; Huang, J.; Chen, L.; Sun, F.; Ding, W.; Peng, C. Why is the invasive plant Sphagneticola trilobata more resistant to high temperature than its native congener? Int. J. Mol. Sci. 2021, 22, 748. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.D.; Ren, D.T.; Pike, S.; Pallardy, S.; Gassmann, W.; Zhang, S.Q. Chloroplast-generated reactive oxygen species are involved in hypersensitive response-like cell death mediated by a mitogen-activated protein kinase cascade. Plant J. 2007, 51, 941–954. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.L.; Zhai, J.J.; Shao, L.; Lin, W.; Peng, C.L. Accumulation of anthocyanins: An adaptation strategy of to low temperature in winter. Front. Plant Sci. 2019, 10, 1049. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Wang, Y.; Weng, Z.; Chen, G.; Peng, C. Adaptation of the Invasive Plant Sphagneticola trilobata (L.) Pruski to Drought Stress. Plants 2024, 13, 2207. https://doi.org/10.3390/plants13162207
Zhang Q, Wang Y, Weng Z, Chen G, Peng C. Adaptation of the Invasive Plant Sphagneticola trilobata (L.) Pruski to Drought Stress. Plants. 2024; 13(16):2207. https://doi.org/10.3390/plants13162207
Chicago/Turabian StyleZhang, Qilei, Ye Wang, Zhilong Weng, Guangxin Chen, and Changlian Peng. 2024. "Adaptation of the Invasive Plant Sphagneticola trilobata (L.) Pruski to Drought Stress" Plants 13, no. 16: 2207. https://doi.org/10.3390/plants13162207
APA StyleZhang, Q., Wang, Y., Weng, Z., Chen, G., & Peng, C. (2024). Adaptation of the Invasive Plant Sphagneticola trilobata (L.) Pruski to Drought Stress. Plants, 13(16), 2207. https://doi.org/10.3390/plants13162207