
Plasmodesmata Function and Callose Deposition in Plant Disease Defense

 , , and
, , and
Abstract
1. Introduction
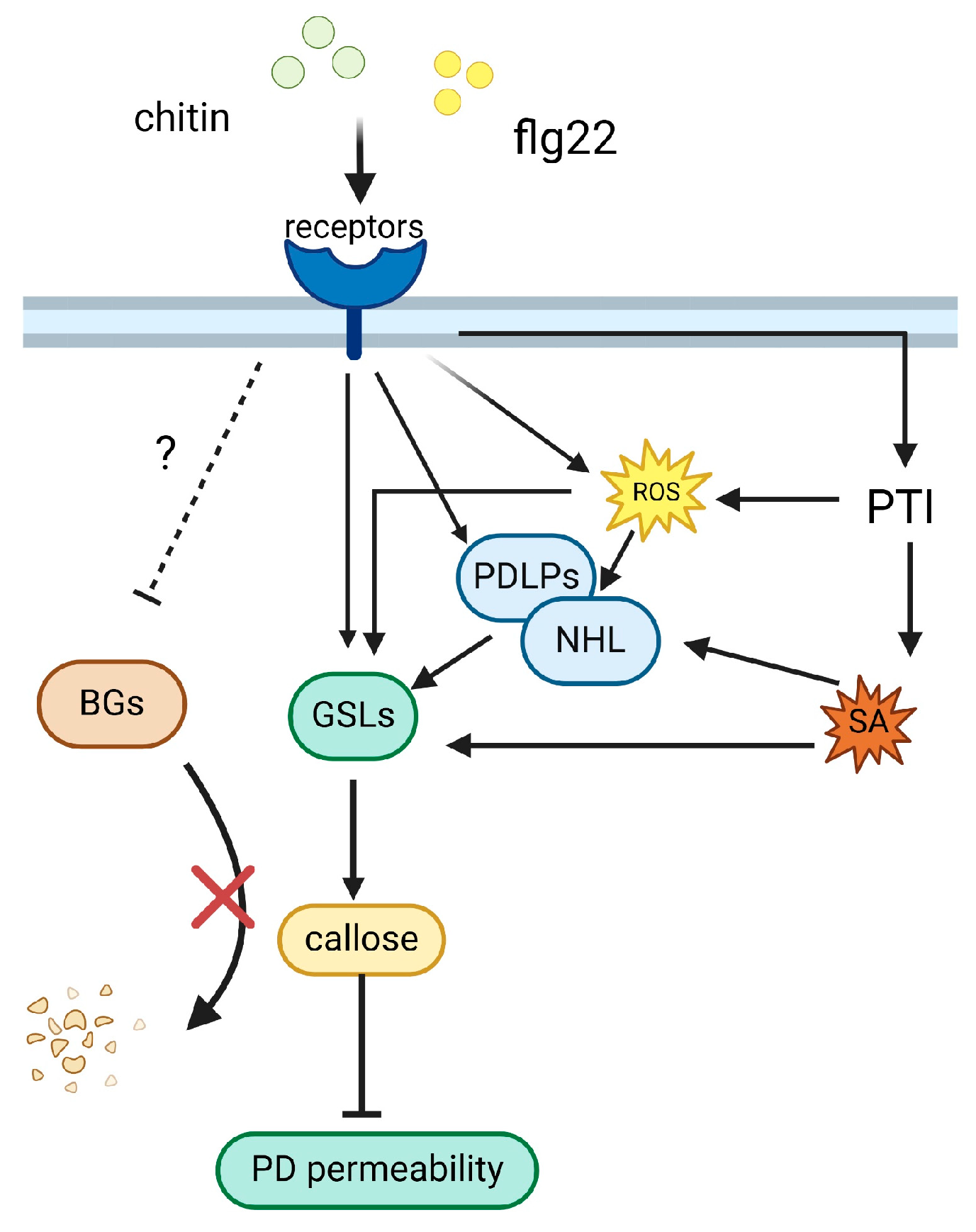
2. Plant-Immunity-Related Callose Accumulation
2.1. Callose Accumulation as a Hallmark of Plant Immune Responses
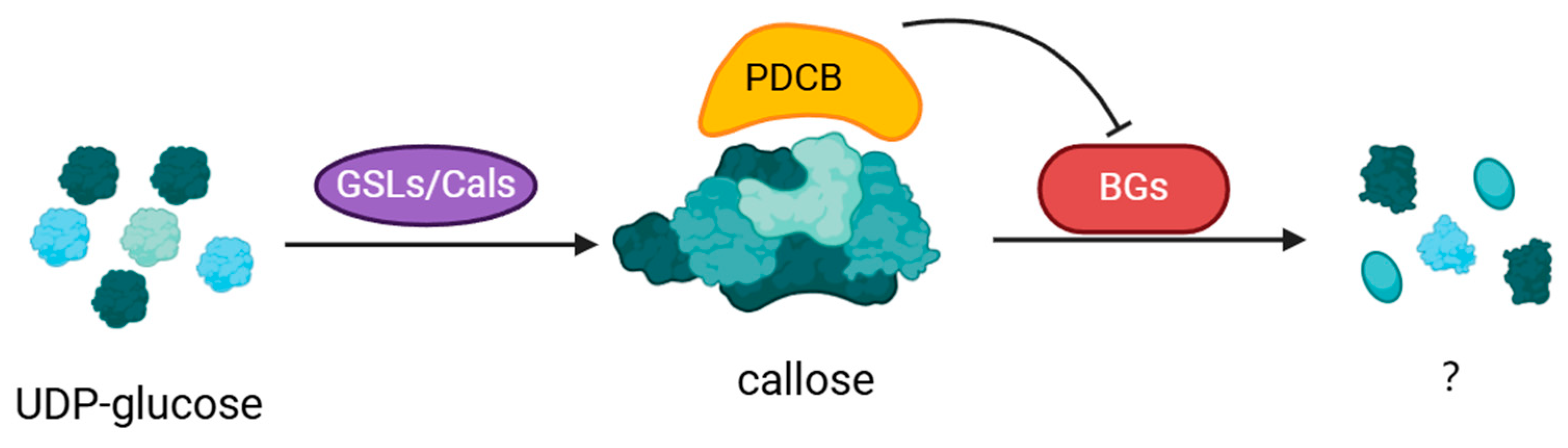
2.2. Enzymatic Regulation of Callose Deposition
2.2.1. Callose Synthase (CalS/GSL)
2.2.2. Callose Hydrolase
2.2.3. Plasmodesmata Callose Binding Protein (PDCB)
3. PD Is a Crucial Plant Component
3.1. Formation and Structure of PD
3.2. Callose Changes Permeability of PD
3.3. PD’s Contribution to Plant Immunity
3.3.1. PD’s Response to Viral Infestation
3.3.2. PD’s Reaction to Fungal Intrusion
3.3.3. PD Response to Bacteria Infestation
3.3.4. Pathogen Manipulation of Callose and Plasmodesmata in Plant Defense Mechanisms
4. Callose Controls PD Permeability for Plant Disease Resistance
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| ANK | Ankyrin repeat-containing protein |
| BG | β-glucanase |
| CalS | Callose synthase |
| CCs | Companion cells |
| CFDA | Carboxyfluorescein diacetate |
| DANS | Drop-and-see |
| EGFP | Enhanced green fluorescent protein |
| ETI | Effector-triggered immunity |
| GA | Gibberellin |
| GFP | Green fluorescent protein |
| GSL | Glucan synthase-like |
| IH | Invasive hyphae |
| MPs | Movement proteins |
| NCAPs | Non-cell-autonomous proteins |
| PAMPs | Pathogen-associated molecular patterns |
| PD | Plasmodesmata |
| PDCB | Plasmodesmata callose-binding protein |
| PDGB | PD-associated b-1,3 glucanase |
| PDLP | PD-located protein |
| PR | Pathogenesis-related protein |
| PTI | PAMP-triggered immunity |
| ROS | Reactive oxygen species |
| SA | Salicylic acid |
| SAR | Systemic acquired resistance |
| SEs | Sieve elements |
| SELs | Size exclusion limits |
References
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Dangl, J.L.; Horvath, D.M.; Staskawicz, B.J. Pivoting the Plant Immune System from Dissection to Deployment. Science 2013, 341, 746–751. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, F.; Tsuda, K. Understanding the Plant Immune System. Mol. Plant-Microbe Interact. 2010, 23, 1531–1536. [Google Scholar] [CrossRef]
- Bigeard, J.; Colcombet, J.; Hirt, H. Signaling Mechanisms in Pattern-Triggered Immunity (PTI). Mol. Plant 2015, 8, 521–539. [Google Scholar] [CrossRef]
- Zhou, J.M.; Zhang, Y. Plant Immunity: Danger Perception and Signaling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef]
- Thomma, B.P.H.J.; Nürnberger, T.; Joosten, M.H.A.J. Of PAMPs and Effectors: The Blurred PTI-ETI Dichotomy. Plant Cell 2011, 23, 4–15. [Google Scholar] [CrossRef]
- Ngou, B.P.M.; Ding, P.; Jones, J.D.G. Thirty years of resistance: Zig-zag through the plant immune system. Plant Cell 2022, 34, 1447–1478. [Google Scholar] [CrossRef]
- Yuan, M.; Ngou, B.P.M.; Ding, P.; Xin, X.-F. PTI-ETI crosstalk: An integrative view of plant immunity. Curr. Opin. Plant Biol. 2021, 62, 102030. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Hong, Z.; Sivaramakrishnan, M.; Mahfouz, M.; Verma, D.P. Callose synthase (CalS5) is required for exine formation during microgametogenesis and for pollen viability in Arabidopsis. Plant J. 2005, 42, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, S.; Zinkl, G.M.; Swanson, R.J.; Maruyama, D.; Preuss, D. Callose (beta-1,3 glucan) is essential for Arabidopsis pollen wall patterning, but not tube growth. BMC Plant Biol. 2005, 5, 22. [Google Scholar] [CrossRef]
- Enns, L.C.; Kanaoka, M.M.; Torii, K.U.; Comai, L.; Okada, K.; Cleland, R.E. Two callose synthases, GSL1 and GSL5, play an essential and redundant role in plant and pollen development and in fertility. Plant Mol. Biol. 2005, 58, 333–349. [Google Scholar] [CrossRef] [PubMed]
- Seale, M. Callose Deposition during Pollen Development. Plant Physiol. 2020, 184, 564–565. [Google Scholar] [CrossRef]
- Cai, G.; Faleri, C.; Del Casino, C.; Emons, A.M.C.; Cresti, M. Distribution of Callose Synthase, Cellulose Synthase, and Sucrose Synthase in Tobacco Pollen Tube Is Controlled in Dissimilar Ways by Actin Filaments and Microtubules. Plant Physiol. 2011, 155, 1169–1190. [Google Scholar] [CrossRef] [PubMed]
- Parre, E.; Geitmann, A. More Than a Leak Sealant. The Mechanical Properties of Callose in Pollen Tubes. Plant Physiol. 2005, 137, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Xie, B.; Wang, X.; Zhu, M.; Zhang, Z.; Hong, Z. CalS7 encodes a callose synthase responsible for callose deposition in the phloem. Plant J. 2011, 65, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, K.; Kollmann, R. Primary and secondary plasmodesmata Structure, origin, and functioning. Protoplasma 2001, 216, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z. A cell plate specific callose synthase and its interaction with phragmoplastin. Plant Cell 2001, 13, 755–768. [Google Scholar] [PubMed]
- Thiele, K.; Wanner, G.; Kindzierski, V.; Jurgens, G.; Mayer, U.; Pachl, F.; Assaad, F.F. The timely deposition of callose is essential for cytokinesis in Arabidopsis. Plant J. 2009, 58, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Radford, J.E.; Vesk, M.; Overall, R.L. Callose deposition at plasmodesmata. Protoplasma 1998, 201, 30–37. [Google Scholar] [CrossRef]
- Currier, H.B. Callose substance in plant cells. Am. J. Bot. 1957, 44, 478–488. [Google Scholar] [CrossRef]
- Wu, S.W.; Kumar, R.; Iswanto, A.B.B.; Kim, J.Y. Callose balancing at plasmodesmata. J. Exp. Bot. 2018, 69, 5325–5339. [Google Scholar] [CrossRef] [PubMed]
- Amsbury, S.; Kirk, P.; Benitez-Alfonso, Y. Emerging models on the regulation of intercellular transport by plasmodesmata-associated callose. J. Exp. Bot. 2017, 69, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Huang, L.J.; Feng, D.; Jiang, W.; Miu, W.; Li, N. Plasmodesmata-Related Structural and Functional Proteins: The Long Sought-After Secrets of a Cytoplasmic Channel in Plant Cell Walls. Int. J. Mol. Sci. 2019, 20, 2946. [Google Scholar] [CrossRef] [PubMed]
- Lucas, W.J.; Ham, B.K.; Kim, J.Y. Plasmodesmata–bridging the gap between neighboring plant cells. Trends Cell Biol. 2009, 19, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Miras, M.; Pottier, M.; Schladt, T.M.; Ejike, J.O.; Redzich, L.; Frommer, W.B.; Kim, J.Y. Plasmodesmata and their role in assimilate translocation. J. Plant Physiol. 2022, 270, 153633. [Google Scholar] [CrossRef] [PubMed]
- Sager, R.E.; Lee, J.Y. Plasmodesmata at a glance. J. Cell Sci. 2018, 131, jcs209346. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, X.; Fan, B.; Zhu, C.; Chen, Z. Regulation and Function of Defense-Related Callose Deposition in Plants. Int. J. Mol. Sci. 2021, 22, 2393. [Google Scholar] [CrossRef] [PubMed]
- German, L.; Yeshvekar, R.; Benitez-Alfonso, Y. Callose metabolism and the regulation of cell walls and plasmodesmata during plant mutualistic and pathogenic interactions. Plant Cell Environ. 2022, 46, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Brown, I.; Mansfield, J.; Bonas, U. The hrp genes in Xanthomonas campestris pv. vesicatoria determine ability to suppress papilla deposition in pepper mesophyll cells. Mol. Plant-Microbe Interact. MPMI 1995, 8, 825–836. [Google Scholar] [CrossRef]
- O’Connell, R.J.; Brown, I.R.; Mansfield, J.W.; Bailey, J.A.; Mazau, D.; Rumeau, D.; Esquerre-Tugaye, M.T. Immunocytochemical localisation of hydroxyproline-rich glycoproteins accumulating in melon and bean at sites of resistance to bacteria and fungi. Mol. Plant-Microbe Interact. 1990, 3, 33–40. [Google Scholar] [CrossRef]
- Brown, I.; Trethowan, J.; Kerry, M.; Mansfield, J.; Bolwell, G.P. Localization of components of the oxidative cross-linking of glycoproteins and of callose synthesis in papillae formed during the interaction between non-pathogenic strains of Xanthomonas campestris and French bean mesophyll cells. Plant J. 2002, 15, 333–343. [Google Scholar] [CrossRef]
- Nishimura, M.T.; Stein, M.; Hou, B.-H.; Vogel, J.P.; Edwards, H.; Somerville, S.C. Loss of a callose synthase results in salicylic acid-dependent disease resistance. Science 2003, 301, 969–972. [Google Scholar] [CrossRef]
- Sankoh, A.F.; Burch-Smith, T.M. Approaches for investigating plasmodesmata and effective communication. Curr. Opin. Plant Biol. 2021, 64, 102143. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Gomez, L.; Felix, G.; Boller, T. A single locus determines sensitivity to bacterial flagellin in Arabidopsis thaliana. Plant J. 1999, 18, 277–284. [Google Scholar] [CrossRef]
- Kohler, A.; Schwindling, S.; Conrath, U. Benzothiadiazole-induced priming for potentiated responses to pathogen infection, wounding, and infiltration of water into leaves requires the NPR1/NIM1 gene in Arabidopsis. Plant Physiol. 2002, 128, 1046–1056. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Hong, Z.; Chatterjee, J.; Kim, S.; Verma, D.P. Expression of callose synthase genes and its connection with Npr1 signaling pathway during pathogen infection. Planta 2008, 229, 87–98. [Google Scholar] [CrossRef]
- DebRoy, S.; Thilmony, R.; Kwack, Y.-B.; Nomura, K.; He, S.Y. A family of conserved bacterial effectors inhibits salicylic acid-mediated basal immunity and promotes disease necrosis in plants. Proc. Natl. Acad. Sci. USA 2004, 101, 9927–9932. [Google Scholar] [CrossRef]
- Ton, J.; Mauch-Mani, B. Beta-amino-butyric acid-induced resistance against necrotrophic pathogens is based on ABA-dependent priming for callose. Plant J. 2004, 38, 119–130. [Google Scholar] [CrossRef]
- Luna, E.; Pastor, V.; Robert, J.; Flors, V.; Mauch-Mani, B.; Ton, J. Callose Deposition: A Multifaceted Plant Defense Response. Mol. Plant-Microbe Interact. 2010, 24, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Richmond, T.A.; Somerville, C.R. The cellulose synthase superfamily. Plant Physiol. 2000, 124, 495–498. [Google Scholar] [CrossRef]
- Verma, D.P.S.; Hong, Z. Plant callose synthase complexes. Plant Mol. Biol. 2001, 47, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Schneider, R.; Hanak, T.; Persson, S.; Voigt, C.A. Cellulose and callose synthesis and organization in focus, what’s new? Curr. Opin. Plant Biol. 2016, 34, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.K.; Lipka, V.; Burton, R.A.; Panstruga, R.; Strizhov, N.; Schulze-Lefert, P.; Fincher, G.B. An Arabidopsis Callose Synthase, GSL5, Is Required for Wound and Papillary Callose Formation. Plant Cell 2003, 15, 2503–2513. [Google Scholar] [CrossRef]
- Blümke, A.; Somerville, S.C.; Voigt, C.A. Transient expression of the Arabidopsis thaliana callose synthase PMR4 increases penetration resistance to powdery mildew in barley. Adv. Biosci. Biotechnol. 2013, 4, 810–813. [Google Scholar] [CrossRef]
- Ellinger, D.; Naumann, M.; Falter, C.; Zwikowics, C.; Jamrow, T.; Manisseri, C.; Somerville, S.C.; Voigt, C.A. Elevated early callose deposition results in complete penetration resistance to powdery mildew in Arabidopsis. Plant Physiol. 2013, 4, 810–813. [Google Scholar] [CrossRef] [PubMed]
- Barratt, D.H.; Kolling, K.; Graf, A.; Pike, M.; Calder, G.; Findlay, K.; Zeeman, S.C.; Smith, A.M. Callose synthase GSL7 is necessary for normal phloem transport and inflorescence growth in Arabidopsis. Plant Physiol. 2011, 155, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Guseman, J.M.; Lee, J.S.; Bogenschutz, N.L.; Peterson, K.M.; Virata, R.E.; Xie, B.; Kanaoka, M.M.; Hong, Z.; Torii, K.U. Dysregulation of cell-to-cell connectivity and stomatal patterning by loss-of-function mutation in Arabidopsis CHORUS (GLUCAN SYNTHASE-LIKE 8). Development 2010, 137, 1731–1741. [Google Scholar] [CrossRef]
- Srivastava, V.; Malm, E.; Sundqvist, G.; Bulone, V. Quantitative proteomics reveals that plasma membrane microdomains from poplar cell suspension cultures are enriched in markers of signal transduction, molecular transport, and callose biosynthesis. Mol. Cell. Proteom. 2013, 12, 3874–3885. [Google Scholar] [CrossRef]
- Beffa, R.; Frederick Meins, J. Pathogenesis-related functions of plant beta-1,3-glucanases investigated by antisense transformation—A review. Gene 1996, 179, 97–103. [Google Scholar] [CrossRef]
- Levy, A.; Erlanger, M.; Rosenthal, M.; Epel, B.L. A plasmodesmata-associated beta-1,3-glucanase in Arabidopsis. Plant J. 2007, 49, 669–682. [Google Scholar] [CrossRef]
- Rinne, P.L.; Paul, L.K.; Vahala, J.; Kangasjarvi, J.; van der Schoot, C. Axillary buds are dwarfed shoots that tightly regulate GA pathway and GA-inducible 1,3-beta-glucanase genes during branching in hybrid aspen. J. Exp. Bot. 2016, 67, 5975–5991. [Google Scholar] [CrossRef]
- Simpson, C.; Thomas, C.; Findlay, K.; Bayer, E.; Maule, A.J. An Arabidopsis GPI-anchor plasmodesmal neck protein with callose binding activity and potential to regulate cell-to-cell trafficking. Plant Cell 2009, 21, 581–594. [Google Scholar] [CrossRef]
- Maule, A.J.; Benitez-Alfonso, Y.; Faulkner, C. Plasmodesmata–membrane tunnels with attitude. Curr. Opin. Plant Biol. 2011, 14, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, C.; Zeng, J.; Yu, H.; Li, Y.; Yuan, C. Identification of two additional plasmodesmata localization domains in the tobacco mosaic virus cell-to-cell-movement protein. Biochem. Biophys. Res. Commun. 2020, 521, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Hepler, P.K. Endoplasmic reticulum in the formation of the cell plate and plasmodesmata. Protoplasma 1982, 111, 121–133. [Google Scholar] [CrossRef]
- Van Bel, A.J.; Knoblauch, M. Sieve element and companion cell: The story of the comatose patient and the hyperactive nurse. Funct. Plant Biol. 2000, 27, 477–487. [Google Scholar] [CrossRef]
- Sager, R.; Lee, J.Y. Plasmodesmata in integrated cell signalling: Insights from development and environmental signals and stresses. J. Exp. Bot. 2014, 65, 6337–6358. [Google Scholar] [CrossRef] [PubMed]
- Robards, A.; Lucas, W. Plasmodesmata. Annu. Rev. Plant Biol. 1990, 41, 369–419. [Google Scholar] [CrossRef]
- Lucas, W.J.; Lee, J.Y. Plasmodesmata as a supracellular control network in plants. Nat. Rev. Mol. Cell Biol. 2004, 5, 712–726. [Google Scholar] [CrossRef]
- Peters, W.S.; Jensen, K.H.; Stone, H.A.; Knoblauch, M. Plasmodesmata and the problems with size: Interpreting the confusion. J. Plant Physiol. 2021, 257, 153341. [Google Scholar] [CrossRef]
- Carella, P.; Isaacs, M.; Cameron, R.K. Plasmodesmata-located protein overexpression negatively impacts the manifestation of systemic acquired resistance and the long-distance movement of Defective in Induced Resistance1 in Arabidopsis. Plant Biol. 2015, 17, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Wang, X.; Cui, W.; Sager, R.; Modla, S.; Czymmek, K.; Zybaliov, B.; van Wijk, K.; Zhang, C.; Lu, H.; et al. A plasmodesmata-localized protein mediates crosstalk between cell-to-cell communication and innate immunity in Arabidopsis. Plant Cell 2011, 23, 3353–3373. [Google Scholar] [CrossRef] [PubMed]
- Schoelz, J.E.; Harries, P.A.; Nelson, R.S. Intracellular transport of plant viruses: Finding the door out of the cell. Mol. Plant 2011, 4, 813–831. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Wang, X.; Lee, J.Y. Drop-ANd-See: A simple, real-time, and noninvasive technique for assaying plasmodesmal permeability. Methods Mol. Biol. 2015, 1217, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sager, R.; Lee, J.-Y. Evaluating molecular movement through plasmodesmata. In Methods in Cell Biology; Elsevier: Amsterdam, The Netherlands, 2020; Volume 160, pp. 99–117. [Google Scholar]
- Cui, W.; Lee, J.Y. Arabidopsis callose synthases CalS1/8 regulate plasmodesmal permeability during stress. Nat. Plants 2016, 2, 16034. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z. Modulating the function of a β-1,3-glucanosyltransferase.pdf. Appl. Microbiol. Biotechnol. 2015, 100, 1765–1776. [Google Scholar] [CrossRef]
- Sperisen, C. Comparison of cloned genes provides evidence for intergenomic exchange of DNA in the evolution of a tobacco glucan endo-1,3-j3-glucosidase gene family. Botany 1991, 88, 1820–1824. [Google Scholar] [CrossRef]
- den Hollander, P.W.; Kieper, S.N.; Borst, J.W.; van Lent, J.W.M. The role of plasmodesma-located proteins in tubule-guided virus transport is limited to the plasmodesmata. Arch. Virol. 2016, 161, 2431–2440. [Google Scholar] [CrossRef]
- Su, S.; Liu, Z.; Chen, C.; Zhang, Y.; Wang, X.; Zhu, L.; Miao, L.; Wang, X.C.; Yuan, M. Cucumber mosaic virus movement protein severs actin filaments to increase the plasmodesmal size exclusion limit in tobacco. Plant Cell 2010, 22, 1373–1387. [Google Scholar] [CrossRef]
- Vu, M.H.; Hyun, T.K.; Bahk, S.; Jo, Y.; Kumar, R.; Thiruppathi, D.; Iswanto, A.B.B.; Chung, W.S.; Shelake, R.M.; Kim, J.-Y. ROS-mediated plasmodesmal regulation requires a network of an Arabidopsis receptor-like kinase, calmodulin-like proteins, and callose synthases. Front. Plant Sci. 2023, 13, 1107224. [Google Scholar] [CrossRef]
- Ueki, S.; Spektor, R.; Natale, D.M.; Citovsky, V. ANK, a host cytoplasmic receptor for the Tobacco mosaic virus cell-to-cell movement protein, facilitates intercellular transport through plasmodesmata. PLoS Pathog. 2010, 6, e1001201. [Google Scholar] [CrossRef] [PubMed]
- Pouwels, J.; Carette, J.E.; Lent, J.V.; Wellink, J. Cowpea mosaic virus effects on host cell processes. Mol. Plant Pathol. 2002, 3, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Pouwels, J.; Van Der Velden, T.; Willemse, J.; Borst, J.; Van Lent, J.; Bisseling, T.; Wellink, J. Studies on the origin and structure of tubules made by the movement protein of Cowpea mosaic virus. J. Gen. Virol. 2004, 85, 3787–3796. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.L.; Bayer, E.M.; Ritzenthaler, C.; Fernandez-Calvino, L.; Maule, A.J. Specific targeting of a plasmodesmal protein affecting cell-to-cell communication. PLoS Biol. 2008, 6, e7. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-H.; Tian, G.-W.; Gafni, Y.; Citovsky, V. Effects of calreticulin on viral cell-to-cell movement. Plant Physiol. 2005, 138, 1866–1876. [Google Scholar] [CrossRef]
- Kankanala, P.; Czymmek, K.; Valent, B. Roles for rice membrane dynamics and plasmodesmata during biotrophic invasion by the blast fungus. Plant Cell 2007, 19, 706–724. [Google Scholar] [CrossRef]
- Cheval, C.; Samwald, S.; Johnston, M.G.; de Keijzer, J.; Breakspear, A.; Liu, X.; Bellandi, A.; Kadota, Y.; Zipfel, C.; Faulkner, C. Chitin perception in plasmodesmata characterizes submembrane immune-signaling specificity in plants. Proc. Natl. Acad. Sci. USA 2020, 117, 9621–9629. [Google Scholar] [CrossRef]
- Khang, C.H.; Berruyer, R.; Giraldo, M.C.; Kankanala, P.; Park, S.Y.; Czymmek, K.; Kang, S.; Valent, B. Translocation of Magnaporthe oryzae effectors into rice cells and their subsequent cell-to-cell movement. Plant Cell 2010, 22, 1388–1403. [Google Scholar] [CrossRef]
- Vidhyasekaran, P.; Ponmalar, T.R.; Samiyappan, R.; Velazhahan, R.; Vimala, R.; Ramanathan, A.; Paranidharan, V.; Muthukrishn, S. Host-Specific Toxin Production by Rhizoctonia solani, the Rice Sheath Blight Pathogen. Biochem. Cell Biol. 1997, 87, 1258–1263. [Google Scholar] [CrossRef]
- Li, Z.; Variz, H.; Chen, Y.; Liu, S.-L.; Aung, K. Plasmodesmata-Dependent Intercellular Movement of Bacterial Effectors. Front. Plant Sci. 2021, 12, 640277. [Google Scholar] [CrossRef]
- Iswanto, A.B.B.; Vu, M.H.; Pike, S.; Lee, J.; Kang, H.; Son, G.H.; Kim, J.Y.; Kim, S.H. Pathogen effectors: What do they do at plasmodesmata? Mol. Plant Pathol. 2022, 23, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Tee, E.E.; Johnston, M.G.; Papp, D.; Faulkner, C. A PDLP-NHL3 complex integrates plasmodesmal immune signaling cascades. Proc. Natl. Acad. Sci. USA 2023, 120, e2216397120. [Google Scholar] [CrossRef] [PubMed]
- Karasov, T.L.; Horton, M.W.; Bergelson, J. Genomic variability as a driver of plant–pathogen coevolution? Curr. Opin. Plant Biol. 2014, 18, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.J. Molecular Responses of Host Tissue Carbon Metabolism to Infection by Powdery Mildew (Erysiphe sp.). Ph.D. Thesis, University of Southampton, Southampton, UK, 2000. [Google Scholar]
- Veneault-Fourrey, C.; Parisot, D.; Gourgues, M.; Laugé, R.; Lebrun, M.-H.; Langin, T. The tetraspanin gene ClPLS1 is essential for appressorium-mediated penetration of the fungal pathogen Colletotrichum lindemuthianum. Fungal Genet. Biol. 2005, 42, 306–318. [Google Scholar] [CrossRef]
- Asselbergh, B.; Höfte, M. Basal tomato defences to Botrytis cinerea include abscisic acid-dependent callose formation. Physiol. Mol. Plant Pathol. 2007, 71, 33–40. [Google Scholar] [CrossRef]
- López-Cruz, J.; Óscar, C.S.; Emma, F.C.; Pilar, G.A.; Carmen, G.B. Absence of Cu–Zn superoxide dismutase BCSOD1 reduces Botrytis cinerea virulence in Arabidopsis and tomato plants, revealing interplay among reactive oxygen species, callose and signalling pathways. Mol. Plant Pathol. 2017, 18, 16–31. [Google Scholar] [CrossRef]
- Scalschi, L.; Sanmartín, M.; Camañes, G.; Troncho, P.; Sánchez-Serrano, J.J.; García-Agustín, P.; Vicedo, B. Silencing of OPR3 in tomato reveals the role of OPDA in callose deposition during the activation of defense responses against Botrytis cinerea. Plant J. 2015, 81, 304–315. [Google Scholar] [CrossRef]
- Tomczynska, I.; Stumpe, M.; Doan, T.G.; Mauch, F. A Phytophthora effector protein promotes symplastic cell-to-cell trafficking by physical interaction with plasmodesmata-localised callose synthases. New Phytol. 2020, 227, 1467–1478. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, J.-M.; Flores, S.; Keefe, D.; Ahl-Goy, P.; Meins, F. The function of vacuolar β-1, 3-glucanase investigated by antisense transformation. Susceptibility of transgenic Nicotiana sylvestris plants to Cercospora nicotianae infection. Plant Mol. Biol. 1992, 19, 803–813. [Google Scholar] [CrossRef]
- Cilia, M.L.; Jackson, D. Plasmodesmata form and function. Curr. Opin. Cell Biol. 2004, 16, 500–506. [Google Scholar] [CrossRef]
- Shi, H.; Liu, Y.; Ding, A.; Wang, W.; Sun, Y. Induced defense strategies of plants against Ralstonia solanacearum. Front. Microbiol. 2023, 14, 1059799. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; He, Y.; Han, X.; Sun, Q.; Xuan, Y.H. beta-Glucanase Family Genes Promote Resistance to Sheath Blight in Rice by Inhibiting the Permeability of Plasmodesmata. J. Agric. Food Chem. 2023, 71, 9667–9676. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Protein Function | Regulation of PD Permeability | References |
|---|---|---|---|
| GSLs/CalS | Callose synthesis | Reduce | [9,32,36,43,46,66] |
| BGs | Hydrolyze callose | Increase | [50,51,67,68] |
| PDCBs | Bind callose and enhance stability | Reduce | [52] |
| PDLPs | PD localization and regulate PD SEL | Reduce | [59,62,69] |
| PDMP | Enlarges plasmodesmata size | Increase | [63,69,70] |
| Microbe (or Effector) | Interaction with Callose/Plasmodesmata | References |
|---|---|---|
| Pseudomonas syringae | Induces callose deposition | [81] |
| Colletotrichum lindemuthianum | Induces callose accumulation | [86] |
| Xanthomonas campestris | Flg22 triggers callose synthesis | [83] |
| Botrytis cinerea | Secretes enzymes that degrade callose | [87,88,89] |
| Phytophthora infestans | Effectors target plasmodesmata | [90] |
| Erysiphe cichoracearum | Induces callose accumulation | [84,85] |
| Magnaporthe oryzae | Induces callose accumulation | [77] |
| Hyaloperonospora arabidopsidis | Effectors alter PD permeability | [82] |
| Cercospora nicotianae | Blocks the induction of glucanases | [91] |
| Tobacco mosaic virus (TMV) | TMV movement protein (MP) targets PD | [69,72,73,92] |
| Cucumber mosaic virus (CMV) | CMV movement protein increases the PD size | [70,74] |
| Ralstonia solanacearum | Induces callose deposition and reduces PD permeability | [93,94] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Xu, X.; Liu, W.; Feng, Z.; Chen, Q.; Zhou, Y.; Sun, M.; Gan, L.; Zhou, T.; Xuan, Y. Plasmodesmata Function and Callose Deposition in Plant Disease Defense. Plants 2024, 13, 2242. https://doi.org/10.3390/plants13162242
Chen J, Xu X, Liu W, Feng Z, Chen Q, Zhou Y, Sun M, Gan L, Zhou T, Xuan Y. Plasmodesmata Function and Callose Deposition in Plant Disease Defense. Plants. 2024; 13(16):2242. https://doi.org/10.3390/plants13162242
Chicago/Turabian StyleChen, Jingsheng, Xiaofeng Xu, Wei Liu, Ziyang Feng, Quan Chen, You Zhou, Miao Sun, Liping Gan, Tiange Zhou, and Yuanhu Xuan. 2024. "Plasmodesmata Function and Callose Deposition in Plant Disease Defense" Plants 13, no. 16: 2242. https://doi.org/10.3390/plants13162242
APA StyleChen, J., Xu, X., Liu, W., Feng, Z., Chen, Q., Zhou, Y., Sun, M., Gan, L., Zhou, T., & Xuan, Y. (2024). Plasmodesmata Function and Callose Deposition in Plant Disease Defense. Plants, 13(16), 2242. https://doi.org/10.3390/plants13162242