
Sugar Transporter HmSWEET8 Cooperates with HmSTP1 to Enhance Powdery Mildew Susceptibility in Heracleum moellendorffii Hance
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
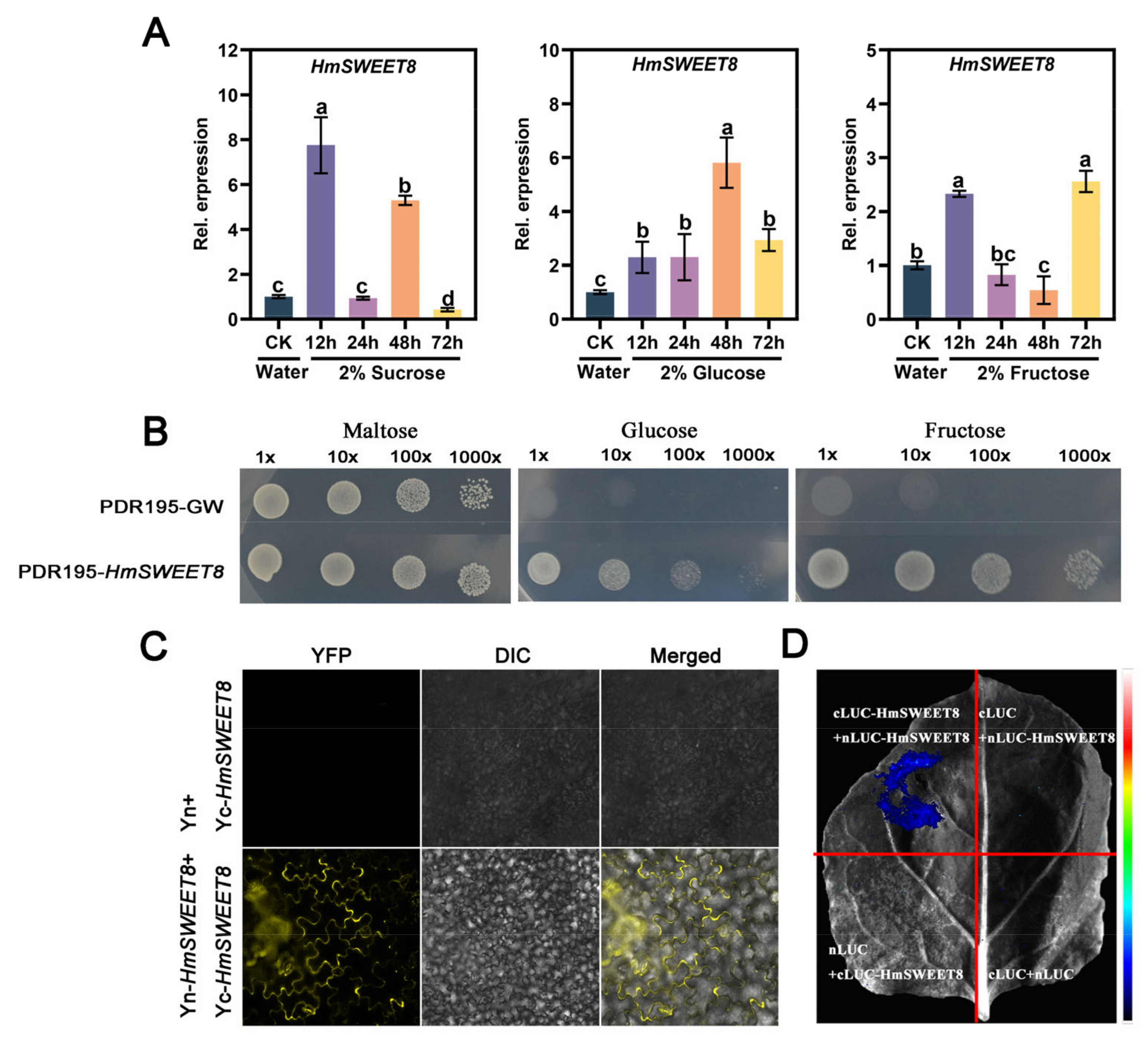
2.1. Cloning of HmSWEET8 Gene in H. moellendorffii
2.2. HmSWEET8 Could Transport Hexose
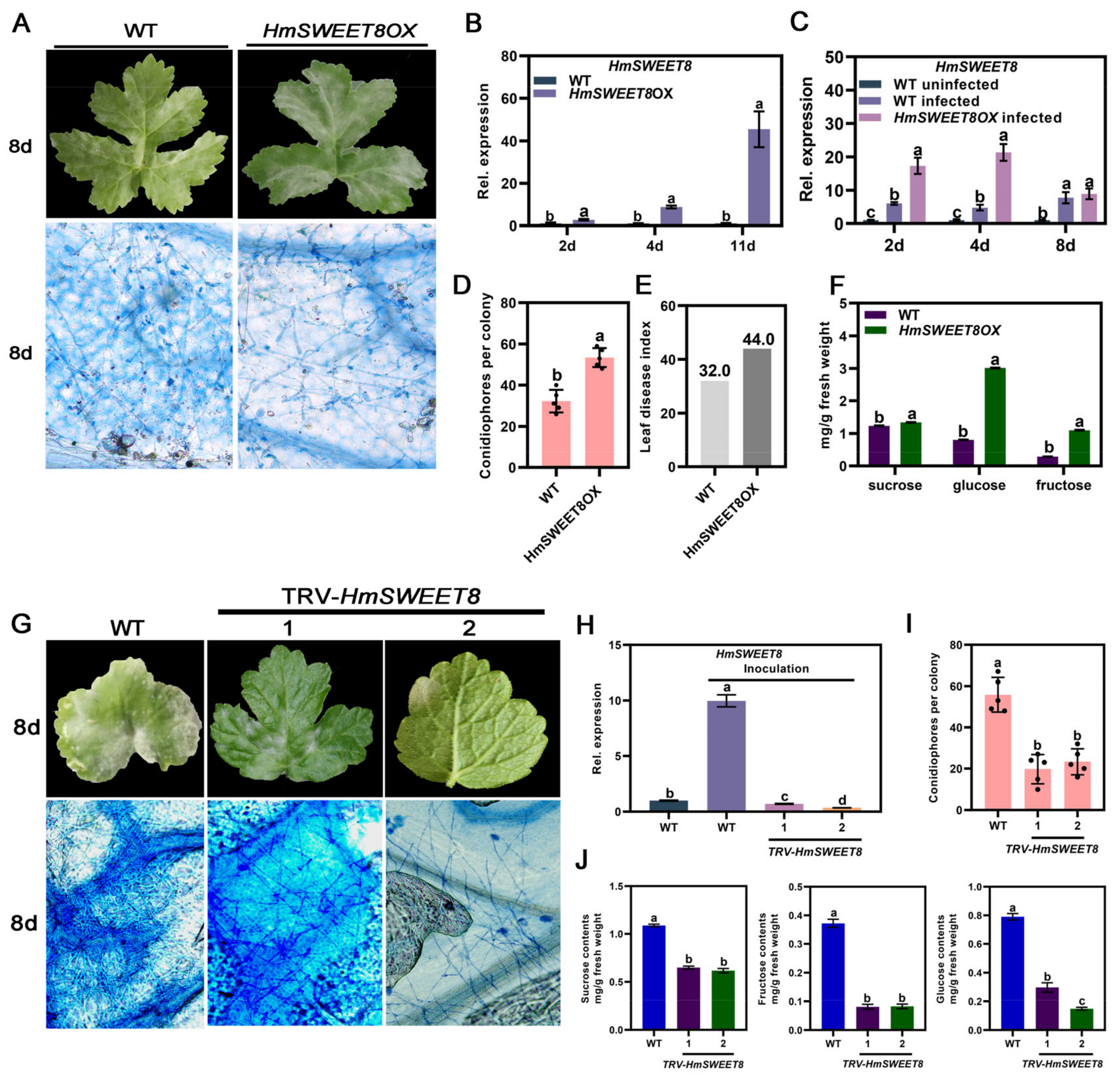
2.3. HmSWEET8 Promoted E. heraclei Infection in H. moellendorffii
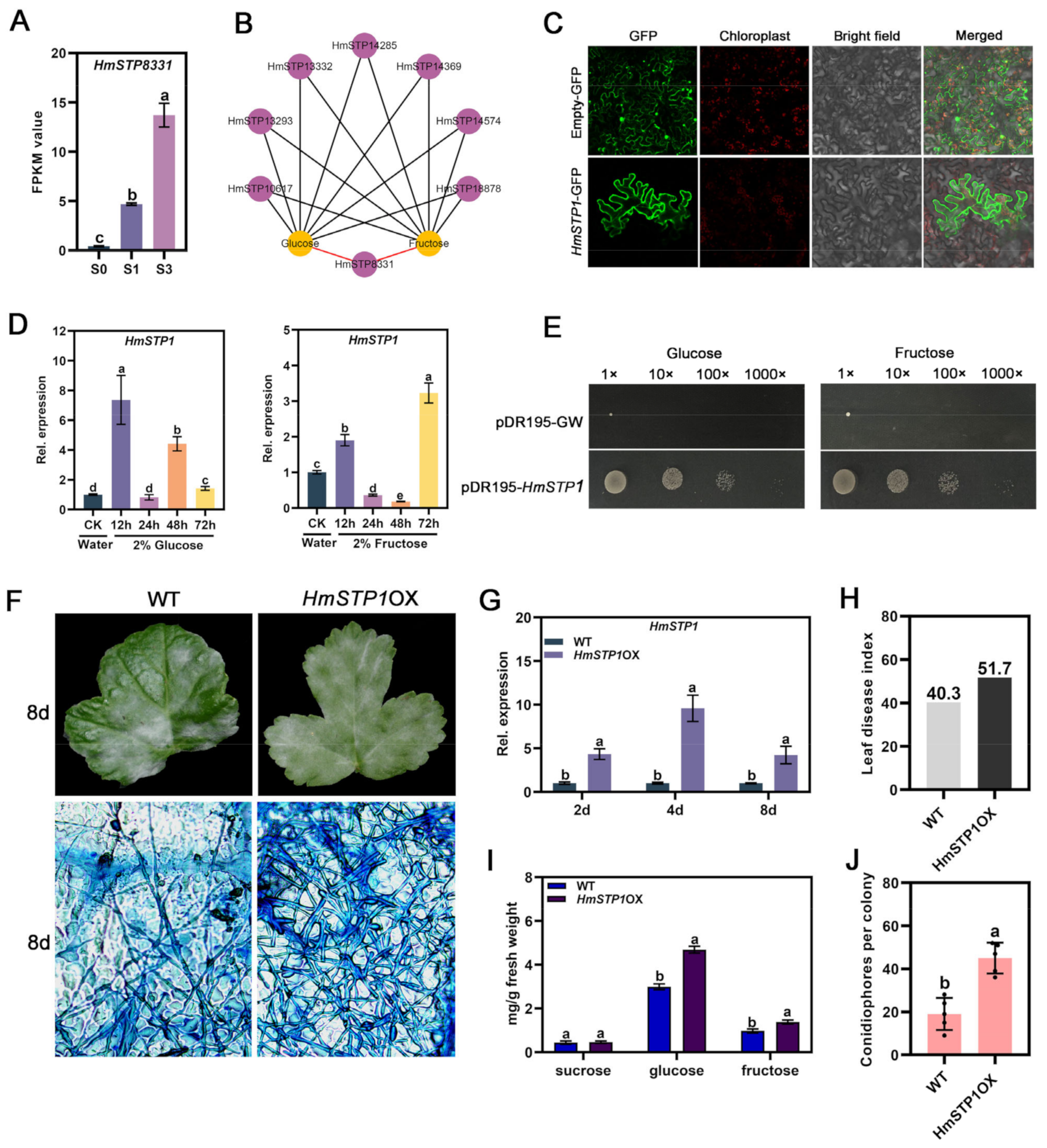
2.4. HmSTP1 Promoted E. heraclei Infection
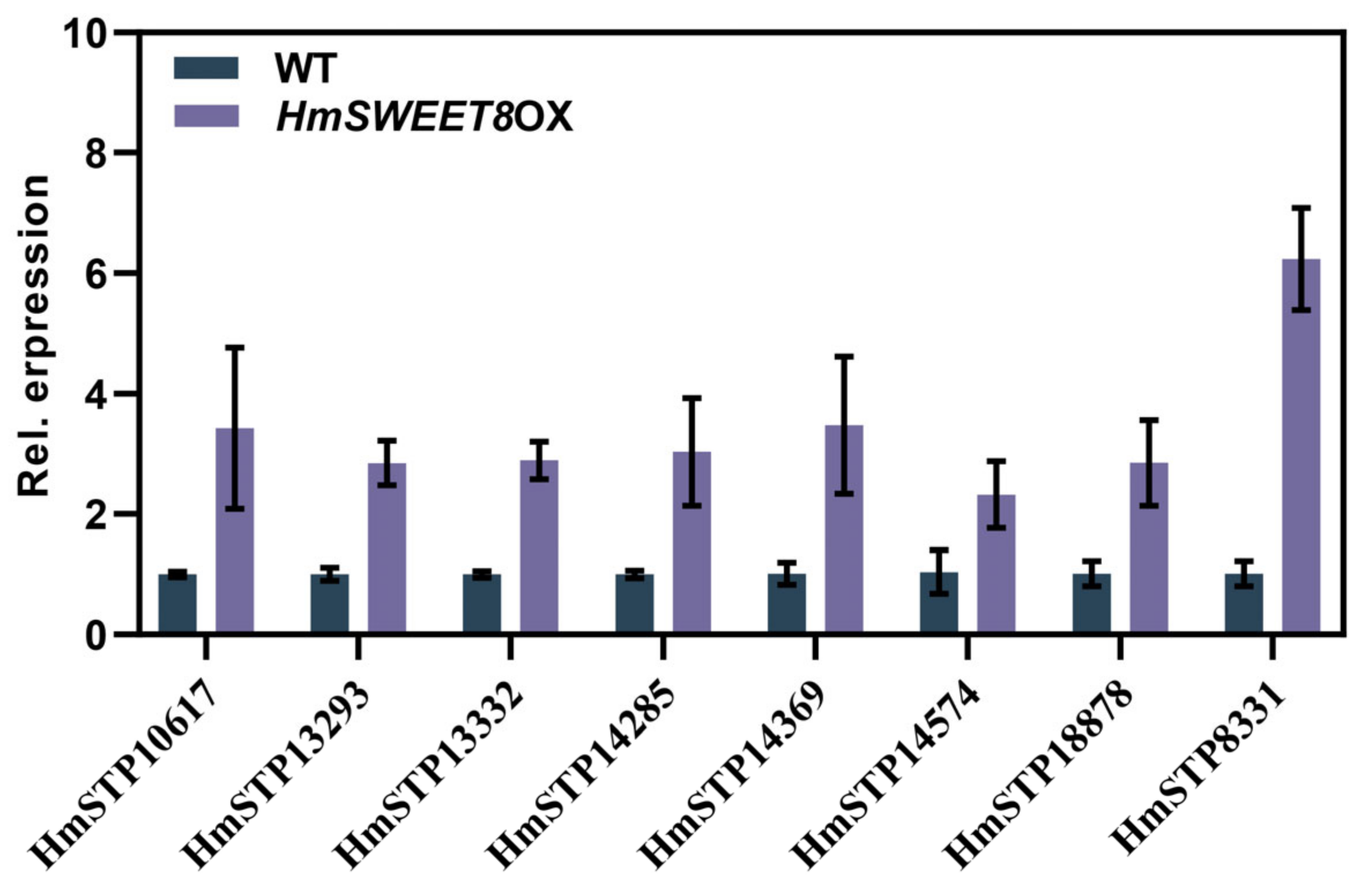
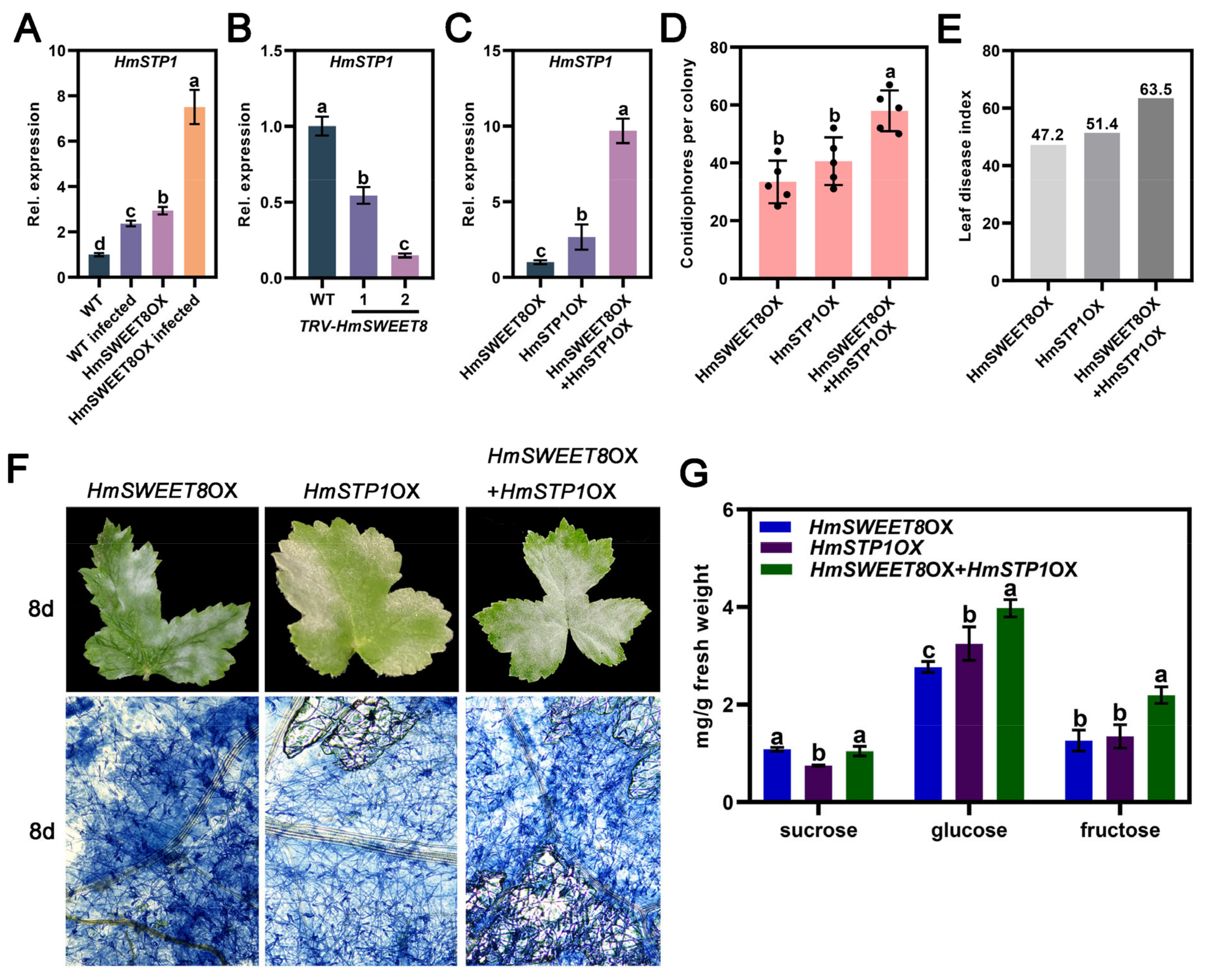
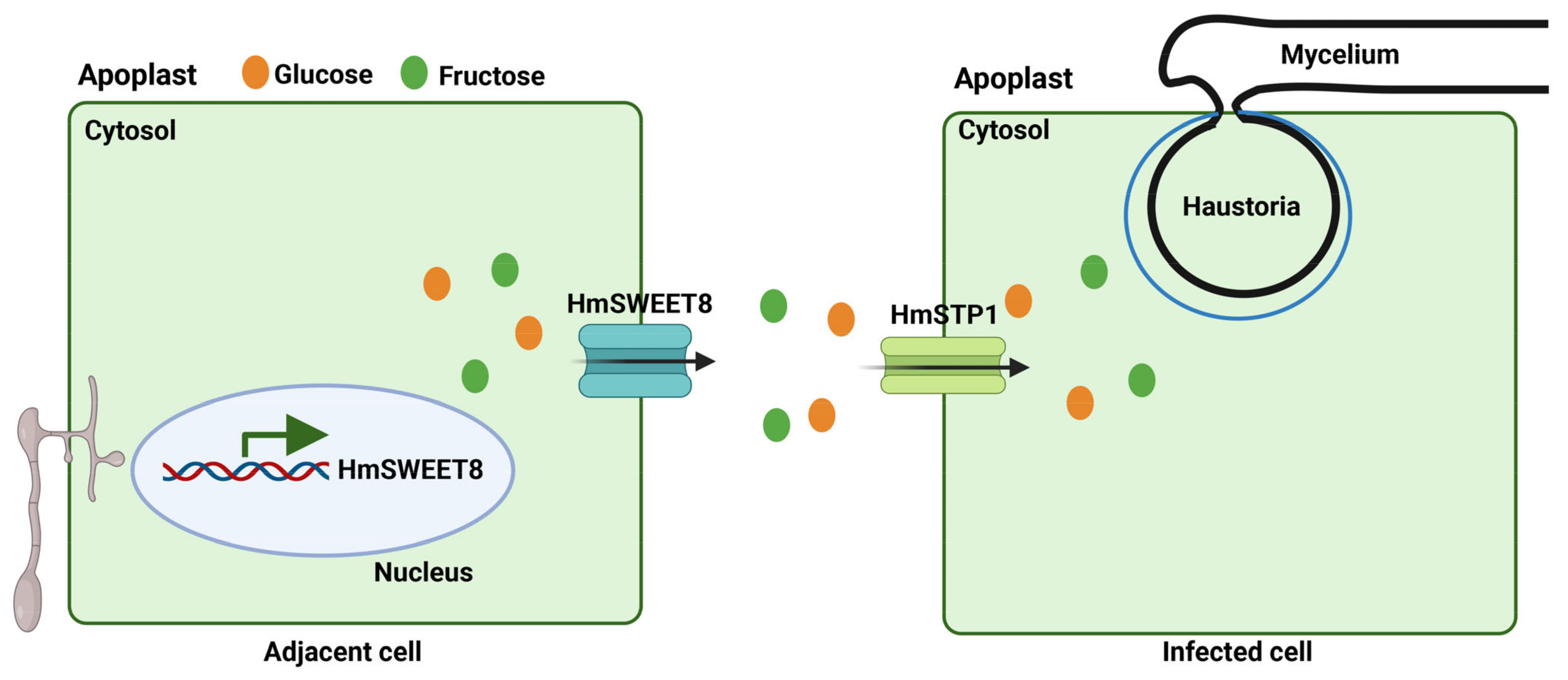
2.5. HmSTP1 May Assist with HmSWEET8 to Promote E. heraclei Infection
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Isolation of HmSWEET8 and HmSTP1
4.3. Subcellular Localization
4.4. Complementation Assays in Yeast
4.5. Bimolecular Fluorescence Complementation (BiFC) Assay
4.6. Luciferase Complementation Assay (LCA)
4.7. Transient Transformation of H. moellendorffii
4.8. Virus-Induced Gene Silencing (VIGS) in H. moellendorffii
4.9. E. heraclei Infection in H. moellendorffii
4.10. Soluble Sugar Content Measurement in H. moellendorffii
4.11. Quantitative Real-Time PCR Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, H.; Wang, Y.; Chang, Q.; Li, Q.; Fang, J.; Cao, N.; Tong, X.; Jiang, X.; Yu, X.; Cheng, Y. Combined Metabolome and Transcriptome Reveal HmF6’H1 Regulating Simple Coumarin Accumulation against Powdery Mildew Infection in Heracleum Moellendorffii Hance. BMC Plant Biol. 2024, 24, 507. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Jiang, X.; Cheng, Y.; Yu, X. First Report of Powdery Mildew Caused by Erysiphe Heraclei on Heracleum Moellendorffii Hance in China. Plant Dis. 2019, 103, 218–232. [Google Scholar] [CrossRef]
- Zhang, T.; Cui, H.; Luan, F.; Liu, H.; Ding, Z.; Amanullah, S.; Zhang, M.; Ma, T.; Gao, P. A Recessive Gene Cmpmr2F Confers Powdery Mildew Resistance in Melon (Cucumis melo L.). Theor. Appl. Genet. 2023, 136, 4. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Saijo, Y.; Nakagami, H.; Takano, Y. Regulation of Sugar Transporter Activity for Antibacterial Defense in Arabidopsis. Science 2016, 354, 1427–1430. [Google Scholar] [CrossRef] [PubMed]
- Bezrutczyk, M.; Yang, J.; Eom, J.-S.; Prior, M.; Sosso, D.; Hartwig, T.; Szurek, B.; Oliva, R.; Vera-Cruz, C.; White, F.F.; et al. Sugar Flux and Signaling in Plant-Microbe Interactions. Plant J. 2018, 93, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-Q.; Qu, X.-Q.; Hou, B.-H.; Sosso, D.; Osorio, S.; Fernie, A.R.; Frommer, W.B. Sucrose Efflux Mediated by SWEET Proteins as a Key Step for Phloem Transport. Science 2012, 335, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Miller, A.J.; Qiu, B.; Huang, Y.; Zhang, K.; Fan, G.; Liu, X. The Role of Sugar Transporters in the Battle for Carbon between Plants and Pathogens. Plant Biotechnol. J. 2024. [Google Scholar] [CrossRef] [PubMed]
- Breia, R.; Conde, A.; Badim, H.; Fortes, A.M.; Gerós, H.; Granell, A. Plant SWEETs: From Sugar Transport to Plant-Pathogen Interaction and More Unexpected Physiological Roles. Plant Physiol. 2021, 186, 836–852. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, J.; Jia, H.; Sosso, D.; Li, T.; Frommer, W.B.; Yang, B.; White, F.F.; Wang, N.; Jones, J.B. Lateral Organ Boundaries 1 Is a Disease Susceptibility Gene for Citrus Bacterial Canker Disease. Proc. Natl. Acad. Sci. USA 2014, 111, E521–E529. [Google Scholar] [CrossRef]
- Blanvillain-Baufumé, S.; Reschke, M.; Solé, M.; Auguy, F.; Doucoure, H.; Szurek, B.; Meynard, D.; Portefaix, M.; Cunnac, S.; Guiderdoni, E.; et al. Targeted Promoter Editing for Rice Resistance to Xanthomonas oryzae Pv. oryzae reveals Differential Activities for SWEET14-Inducing TAL Effectors. Plant Biotechnol. J. 2017, 15, 306–317. [Google Scholar] [CrossRef]
- Cox, K.L.; Meng, F.; Wilkins, K.E.; Li, F.; Wang, P.; Booher, N.J.; Carpenter, S.C.D.; Chen, L.-Q.; Zheng, H.; Gao, X.; et al. TAL Effector Driven Induction of a SWEET Gene Confers Susceptibility to Bacterial Blight of Cotton. Nat. Commun. 2017, 8, 15588. [Google Scholar] [CrossRef]
- Hutin, M.; Sabot, F.; Ghesquière, A.; Koebnik, R.; Szurek, B. A Knowledge-Based Molecular Screen Uncovers a Broad-Spectrum OsSWEET14 Resistance Allele to Bacterial Blight from Wild Rice. Plant J. 2015, 84, 694–703. [Google Scholar] [CrossRef]
- Zhou, J.; Peng, Z.; Long, J.; Sosso, D.; Liu, B.; Eom, J.-S.; Huang, S.; Liu, S.; Vera Cruz, C.; Frommer, W.B.; et al. Gene Targeting by the TAL Effector PthXo2 Reveals Cryptic Resistance Gene for Bacterial Blight of Rice. Plant J. 2015, 82, 632–643. [Google Scholar] [CrossRef]
- Yang, S.; Fu, Y.; Zhang, Y.; Peng Yuan, D.; Li, S.; Kumar, V.; Mei, Q.; Hu Xuan, Y. Rhizoctonia Solani Transcriptional Activator Interacts with Rice WRKY53 and Grassy Tiller 1 to Activate SWEET Transporters for Nutrition. J. Adv. Res. 2023, 50, 1–12. [Google Scholar] [CrossRef]
- Fatima, U.; Senthil-Kumar, M. Sweet Revenge: AtSWEET12 in Plant Defense against Bacterial Pathogens by Apoplastic Sucrose Limitation. Cold Spring Harb. Lab. 2021, 10, 1–49. [Google Scholar] [CrossRef]
- Herbers, K.; Sonnewald, U. Altered Gene Expression Brought about by Inter- and Intracellularly Formed Hexoses and Its Possible Implications for Plant-Pathogen Interactions. J. Plant Res. 1998, 111, 323–328. [Google Scholar] [CrossRef]
- Gebauer, P.; Korn, M.; Engelsdorf, T.; Sonnewald, U.; Koch, C.; Voll, L.M. Sugar Accumulation in Leaves of Arabidopsis Sweet11/Sweet12 Double Mutants Enhances Priming of the Salicylic Acid-Mediated Defense Response. Front. Plant Sci. 2017, 8, 1378. [Google Scholar] [CrossRef]
- Hall, J.L.; Williams, L.E. Assimilate Transport and Partitioning in Fungal Biotrophic Interactions. Funct. Plant Biol. 2000, 27, 549. [Google Scholar] [CrossRef]
- Staples, R.C. Research on the Rust Fungi During the Twentieth Century. Annu. Rev. Phytopathol. 2000, 38, 49–69. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, M.; Tan, L.; Huai, B.; Ma, X.; Pan, Q.; Zheng, P.; Wen, Y.; Zhang, Q.; Zhao, Q.; et al. AtSTP8, an Endoplasmic Reticulum-Localised Monosaccharide Transporter from Arabidopsis, Is Recruited to the Extrahaustorial Membrane during Powdery Mildew Infection. New Phytol. 2021, 230, 2404–2419. [Google Scholar] [CrossRef]
- Huai, B.; Yuan, P.; Ma, X.; Zhang, X.; Jiang, L.; Zheng, P.; Yao, M.; Chen, Z.; Chen, L.; Shen, Q.; et al. Sugar Transporter TaSTP3 Activation by TaWRKY19/61/82 Enhances Stripe Rust Susceptibility in Wheat. New Phytol. 2022, 236, 266–282. [Google Scholar] [CrossRef]
- Huai, B.; Yang, Q.; Qian, Y.; Qian, W.; Kang, Z.; Liu, J. ABA-Induced Sugar Transporter TaSTP6 Promotes Wheat Susceptibility to Stripe Rust. Plant Physiol. 2019, 181, 1328–1343. [Google Scholar] [CrossRef] [PubMed]
- Voegele, R.T.; Mendgen, K. Rust Haustoria: Nutrient Uptake and Beyond. New Phytol. 2003, 159, 93–100. [Google Scholar] [CrossRef]
- Saja, D.; Janeczko, A.; Barna, B.; Skoczowski, A.; Dziurka, M.; Kornaś, A.; Gullner, G. Powdery Mildew-Induced Hormonal and Photosynthetic Changes in Barley Near Isogenic Lines Carrying Various Resistant Genes. Int. J. Mol. Sci. 2020, 21, 4536. [Google Scholar] [CrossRef]
- Doidy, J.; Grace, E.; Kühn, C.; Simon-Plas, F.; Casieri, L.; Wipf, D. Sugar Transporters in Plants and in Their Interactions with Fungi. Trends Plant Sci. 2012, 17, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Das, S.; Jagadis Gupta, K.; Ranjan, A.; Foyer, C.H.; Thakur, J.K. Physiological Implications of SWEETs in Plants and Their Potential Applications in Improving Source-Sink Relationships for Enhanced Yield. Plant Biotechnol. J. 2023, 21, 1528–1541. [Google Scholar] [CrossRef]
- Chen, L.-Q.; Hou, B.-H.; Lalonde, S.; Takanaga, H.; Hartung, M.L.; Qu, X.-Q.; Guo, W.-J.; Kim, J.-G.; Underwood, W.; Chaudhuri, B.; et al. Sugar Transporters for Intercellular Exchange and Nutrition of Pathogens. Nature 2010, 468, 527–532. [Google Scholar] [CrossRef]
- An, J.; Zeng, T.; Ji, C.; de Graaf, S.; Zheng, Z.; Xiao, T.T.; Deng, X.; Xiao, S.; Bisseling, T.; Limpens, E.; et al. A Medicago Truncatula SWEET Transporter Implicated in Arbuscule Maintenance during Arbuscular Mycorrhizal Symbiosis. New Phytol. 2019, 224, 396–408. [Google Scholar] [CrossRef] [PubMed]
- Bezrutczyk, M.; Hartwig, T.; Horschman, M.; Char, S.N.; Yang, J.; Yang, B.; Frommer, W.B.; Sosso, D. Impaired Phloem Loading in Zmsweet13a,b,c Sucrose Transporter Triple Knock-out Mutants in Zea mays. New Phytol. 2018, 218, 594–603. [Google Scholar] [CrossRef]
- Chang, Q.; Liu, J.; Wang, Q.; Han, L.; Liu, J.; Li, M.; Huang, L.; Yang, J.; Kang, Z. The Effect of Puccinia striiformis f. Sp Tritici on the Levels of Water-Soluble Carbohydrates and the Photosynthetic Rate in Wheat Leaves. Physiol. Mol. Plant Pathol. 2013, 84, 131–137. [Google Scholar] [CrossRef]
- Manck-Götzenberger, J.; Requena, N. Arbuscular Mycorrhiza Symbiosis Induces a Major Transcriptional Reprogramming of the Potato SWEET Sugar Transporter Family. Front. Plant Sci. 2016, 7, 487. [Google Scholar] [CrossRef] [PubMed]
- Fotopoulos, V.; Gilbert, M.J.; Pittman, J.K.; Marvier, A.C.; Buchanan, A.J.; Sauer, N.; Hall, J.L.; Williams, L.E. The Monosaccharide Transporter Gene, AtSTP4, and the Cell-Wall Invertase, Atbetafruct1, Are Induced in Arabidopsis during Infection with the Fungal Biotroph Erysiphe cichoracearum. Plant Physiol. 2003, 132, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Sutton, P.N.; Gilbert, M.J.; Williams, L.E.; Hall, J.L. Powdery Mildew Infection of Wheat Leaves Changes Host Solute Transport and Invertase Activity. Physiol. Plant. 2007, 129, 787–795. [Google Scholar] [CrossRef]
- Helber, N.; Wippel, K.; Sauer, N.; Schaarschmidt, S.; Hause, B.; Requena, N. A Versatile Monosaccharide Transporter That Operates in the Arbuscular Mycorrhizal Fungus Glomus Sp Is Crucial for the Symbiotic Relationship with Plants. Plant Cell 2011, 23, 3812–3823. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.W.; Herrera-Foessel, S.; Lan, C.; Schnippenkoetter, W.; Ayliffe, M.; Huerta-Espino, J.; Lillemo, M.; Viccars, L.; Milne, R.; Periyannan, S.; et al. A Recently Evolved Hexose Transporter Variant Confers Resistance to Multiple Pathogens in Wheat. Nat. Genet. 2015, 47, 1494–1498. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-H.; Song, Y.-H.; Ruan, Y.-L. Sugar Conundrum in Plant-Pathogen Interactions: Roles of Invertase and Sugar Transporters Depend on Pathosystems. J. Exp. Bot. 2022, 73, 1910–1925. [Google Scholar] [CrossRef] [PubMed]
- Asai, Y.; Kobayashi, Y. Increased Expression of the Tomato SISWEET15 Gene during Grey Mold Infection and the Possible Involvement of the Sugar Efflux to Apoplasm in the Disease Susceptibility. J. Plant Pathol. Microbiol. 2016, 7, 1. [Google Scholar] [CrossRef]
- Lemonnier, P.; Gaillard, C.; Veillet, F.; Verbeke, J.; Lemoine, R.; Coutos-Thévenot, P.; La Camera, S. Expression of Arabidopsis Sugar Transport Protein STP13 Differentially Affects Glucose Transport Activity and Basal Resistance to Botrytis cinerea. Plant Mol. Biol. 2014, 85, 473–484. [Google Scholar] [CrossRef]
- Harrison, M.J. Biotrophic Interfaces and Nutrient Transport in Plant/Fungal Symbioses. J. Exp. Bot. 1999, 50, 1013–1022. [Google Scholar] [CrossRef]
- Bracha-Drori, K.; Shichrur, K.; Katz, A.; Oliva, M.; Angelovici, R.; Yalovsky, S.; Ohad, N. Detection of Protein-Protein Interactions in Plants Using Bimolecular Fluorescence Complementation. Plant J. 2004, 40, 419–427. [Google Scholar] [CrossRef]
- Spitzer, B.; Zvi, M.M.B.; Ovadis, M.; Marhevka, E.; Barkai, O.; Edelbaum, O.; Marton, I.; Masci, T.; Alon, M.; Morin, S.; et al. Reverse Genetics of Floral Scent: Application of Tobacco Rattle Virus-Based Gene Silencing in Petunia. Plant Physiol. 2007, 145, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Qu, J.; Bui, H.T.N.; Chua, N.-H. Rapid Analysis of Jatropha Curcas Gene Functions by Virus-Induced Gene Silencing. Plant Biotechnol. J. 2009, 7, 964–976. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Liu, J.; Si, X.; Zhang, S.; Zhang, L.; Tong, X.; Yu, X.; Jiang, X.; Cheng, Y. Sugar Transporter HmSWEET8 Cooperates with HmSTP1 to Enhance Powdery Mildew Susceptibility in Heracleum moellendorffii Hance. Plants 2024, 13, 2302. https://doi.org/10.3390/plants13162302
Liu H, Liu J, Si X, Zhang S, Zhang L, Tong X, Yu X, Jiang X, Cheng Y. Sugar Transporter HmSWEET8 Cooperates with HmSTP1 to Enhance Powdery Mildew Susceptibility in Heracleum moellendorffii Hance. Plants. 2024; 13(16):2302. https://doi.org/10.3390/plants13162302
Chicago/Turabian StyleLiu, Hanbing, Junxia Liu, Xiaohui Si, Shuhong Zhang, Lili Zhang, Xuejiao Tong, Xihong Yu, Xinmei Jiang, and Yao Cheng. 2024. "Sugar Transporter HmSWEET8 Cooperates with HmSTP1 to Enhance Powdery Mildew Susceptibility in Heracleum moellendorffii Hance" Plants 13, no. 16: 2302. https://doi.org/10.3390/plants13162302