
Nutrient Allocation and Growth Responses of Senegalia polyphylla under Varied Fertilizer Regimes for Effective Forest Recovery

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Plant Cultivation
2.2. Measured Variables
2.3. Statistical Analysis
3. Results
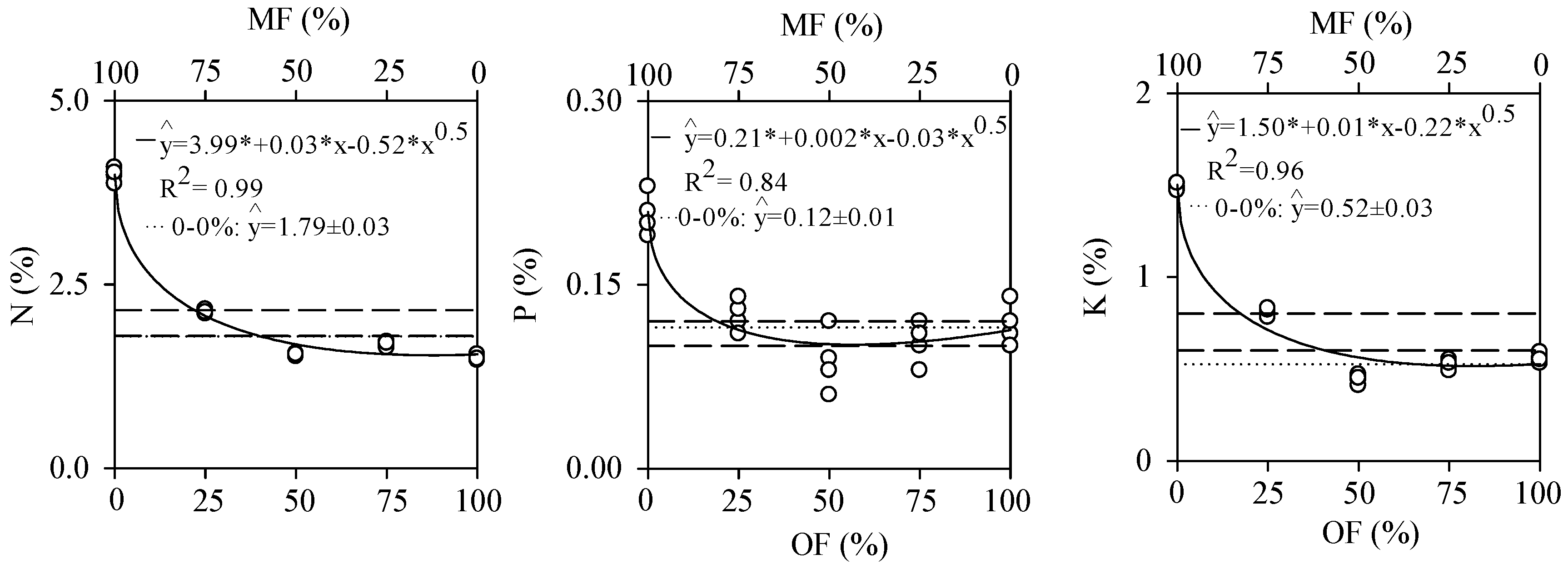
3.1. Nutrient Use Efficiency Results
3.2. Growth Characteristics Results
4. Discussion
4.1. Nutrient Use Efficiency
4.2. Growth Characteristics
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adelino, J.R.P.; Heringer, G.; Diagne, C.; Courchamp, F.; Faria, L.D.B.; Zenni, R.D. The economic costs of biological invasions in Brazil: A first assessment. NeoBiota 2021, 67, 349–374. [Google Scholar] [CrossRef]
- Zenni, R.D.; Ziller, S.R.; da Rosa, C.A.; Sühs, R.B.; Puechagut, P.B.; Marterer, B.T.; Chapla, T.E. Invasive non-native species in Brazil: An updated overview. Biol. Invasions 2024, 26, 2397–2405. [Google Scholar] [CrossRef]
- Canavan, S.; Meyerson, L.A.; Packer, J.G.; Pyšek, P.; Maurel, N.; Lozano, V.; Richardson, D.M.; Brundu, G.; Canavan, K.; Cicatelli, A.; et al. Tall-statured grasses: A useful functional group for invasion science. Biol. Invasions 2019, 21, 37–58. [Google Scholar] [CrossRef]
- Gornish, E.S.; Franklin, K.; Rowe, J.; Barberán, A. Buffelgrass invasion and glyphosate effects on desert soil microbiome communities. Biol. Invasions 2020, 22, 2587–2597. [Google Scholar] [CrossRef]
- Assis, G.B.; Pilon, N.A.; Siqueira, M.F.; Durigan, G. Effectiveness and costs of invasive species control using different techniques to restore cerrado grasslands. Restor. Ecol. 2021, 29, e13219. [Google Scholar] [CrossRef]
- Dezotti, G.; Fidelis, A.; Damasceno, G.; Siqueira, T. Interactive roles of fire seasons and biological invasions in the short-term dynamics of tropical savannas. J. Veg. Sci. 2024, 35, e13250. [Google Scholar] [CrossRef]
- Damasceno, G.; Souza, L.; Pivello, V.R.; Gorgone-Barbosa, E.; Giroldo, P.Z.; Fidelis, A. Impact of invasive grasses on Cerrado under natural regeneration. Biol. Invasions 2018, 20, 3621–3629. [Google Scholar] [CrossRef]
- Mazía, N.; Moyano, J.; Perez, L.; Aguiar, S.; Garibaldi, L.A.; Schlichter, T. The sign and magnitude of tree–grass interaction along a global environmental gradient. Glob. Ecol. Biogeogr. 2016, 25, 1510–1519. [Google Scholar] [CrossRef]
- Almeida, C.; Viani, R.A. Selection of shade trees in forest restoration plantings should not be based on crown tree architecture alone. Restor. Ecol. 2019, 27, 832–839. [Google Scholar] [CrossRef]
- Meli, P.; Isernhagen, I.; Brancalion, P.H.; Isernhagen, E.C.; Behling, M.; Rodrigues, R.R. Optimizing seeding density of fast-growing native trees for restoring the Brazilian Atlantic Forest. Restor. Ecol. 2018, 26, 212–219. [Google Scholar] [CrossRef]
- Souza, M.T.P.; Azevedo, G.B.; Azevedo, G.T.D.O.S.; Teodoro, L.P.R.; Plaster, O.B.; Assunção, P.C.G.; Teodoro, P.E. Growth of native forest species in a mixed stand in the Brazilian Savanna. For. Ecol. Manag. 2020, 462, 118011. [Google Scholar] [CrossRef]
- Barbosa, R.S.; Vale, R.S.; Schwartz, G.; Martins, W.B.R.; Ribeiro, S.S.; Rodrigues, J.I.M.; Ferreira, G.C.; Barbosa, V.M. Restoration of degraded areas after bauxite mining in the eastern Amazon: Which method to apply? Ecol. Eng. 2022, 180, 106639. [Google Scholar] [CrossRef]
- Miller-Adamany, A.; Baumann, D.; Thomsen, M. Facilitating natural succession in a heavily invaded ecosystem. For. Ecol. Manag. 2019, 444, 235–243. [Google Scholar] [CrossRef]
- Thomas, P.A.; Schüler, J.; Boavista, L.D.R.; Torchelsen, F.P.; Overbeck, G.E.; Müller, S.C. Controlling the invader Urochloa decumbens: Subsidies for ecological restoration in subtropical Campos grassland. Appl. Veg. Sci. 2018, 22, 96–104. [Google Scholar] [CrossRef]
- Massa, D.; Benvenuti, S.; Cacini, S.; Lazzereschi, S.; Burchi, G. Effect of hydro-compacting organic mulch on weed control and crop performance in the cultivation of three container-grown ornamental shrubs: Old solutions meet new insights. Sci. Hortic. 2019, 252, 260–267. [Google Scholar] [CrossRef]
- Pokharel, P.; Kwak, J.H.; Chang, S.X. Growth and nitrogen uptake of jack pine seedlings in response to exponential fertilization and weed control in reclaimed soil. Biol. Fertil. Soils 2017, 53, 701–713. [Google Scholar] [CrossRef]
- Krapfl, K.J.; Hatten, J.A.; Roberts, S.D.; Baldwin, B.S.; Rousseau, R.J.; Shankle, M.W. Capacity of biochar application and nitrogen fertilization to mitigate grass competition upon tree seedlings during stand regeneration. For. Ecol. Manag. 2016, 376, 298–309. [Google Scholar] [CrossRef]
- Brancalion, P.H.; Campoe, O.; Mendes, J.C.T.; Noel, C.; Moreira, G.G.; Melis, J.; Noel, C.; Stape, J.L.; Guillemot, J. Intensive silviculture enhances biomass accumulation and tree diversity recovery in tropical forest restoration. Ecol. Appl. 2019, 29, e01847. [Google Scholar] [CrossRef] [PubMed]
- Ornelas, A.C.S.; Providello, A.; Soares, M.R.; Viani, R.A.G. Silvicultural intensification has a limited impact on tree growth in forest restoration plantations in croplands. For. Ecol. Manag. 2022, 503, 119795. [Google Scholar] [CrossRef]
- Tremblay, P.Y.; Thiffault, E.; Pinno, B.D. Effects of land reclamation practices on the productivity of young trembling aspen and white spruce on a reclaimed oil sands mining site in northern Alberta. New For. 2019, 50, 911–942. [Google Scholar] [CrossRef]
- Schott, K.M.; Snively, A.E.; Landhäusser, S.M.; Pinno, B.D. Nutrient loaded seedlings reduce the need for field fertilization and vegetation management on boreal forest reclamation sites. New Forests 2016, 47, 393–410. [Google Scholar] [CrossRef]
- Otero, M.; Salcedo, I.; Txarterina, K.; González-Murua, C.; Duñabeitia, M.K. Quality assessment of Pinus radiata production under sustainable nursery management based on compost tea. J. Plant Nutr. Soil Sci. 2019, 182, 356–366. [Google Scholar] [CrossRef]
- Arif, M.S.; Shahzad, S.M.; Riaz, M.; Yasmeen, T.; Shahzad, T.; Akhtar, M.J.; Bragazza, L.; Buttler, A. Nitrogen-enriched compost application combined with plant growth-promoting rhizobacteria (PGPR) improves seed quality and nutrient use efficiency of sunflower. J. Plant Nutr. Soil Sci. 2017, 180, 464–473. [Google Scholar] [CrossRef]
- Santos, H.G.; Jacomine, P.K.T.; Anjos, L.H.C.; Oliveira, V.A.; Lumbreras, J.F.; Coelho, M.R.; Almeida, J.A.; Cunha, T.J.F.; Oliveira, J.B. Sistema Brasileiro de Classificação de Solos; Embrapa: Brasília, Brazil, 2018. [Google Scholar]
- Teixeira, P.C.; Donagemma, G.K.; Fontana, A.; Teixeira, W.G. Manual de Métodos de Análise de Solo; Embrapa: Rio de Janeiro, Brazil, 2017. [Google Scholar]
- Gonçalves, E.O.; Paiva, H.N.; Neves, J.C.L.; Gomes, J.M. Crescimento de mudas de angico-vermelho (Anadenanthera macrocarpa (Benth.) Brenan) sob diferentes doses de macronutrientes. Rev. Árvore 2008, 32, 1029–1040. [Google Scholar] [CrossRef]
- CFSEMG (Comissão de Fertilidade do Solo do Estado de Minas Gerais). Adubação orgânica. In Recomendações para o Uso de Corretivos e Fertilizantes em Minas Gerais; Ribeiro, A.C., Guimarães, P.T.G., Alvarez, V.H., Eds.; CFSEMG: Viçosa, Brazil, 1999; pp. 87–92. [Google Scholar]
- Silva, F.C. Manual de Análises Químicas de Solos, Plantas e Fertilizantes, 2nd ed.; Embrapa Informações Tecnológicas: Brasília, Brazil, 2009. [Google Scholar]
- O’Brien, P.L.; Thomas, A.L.; Sauer, T.J.; Brauer, D.K. Foliar nutrient concentrations of three economically important tree species in an alley-cropping system. J. Plant Nutr. 2020, 43, 2557–2568. [Google Scholar] [CrossRef]
- Voigtlaender, M.; Brandani, C.B.; Caldeira, D.R.M.; Tardy, F.; Bouillet, J.P.; Gonçalves, J.L.M.; Moreira, M.Z.; Leite, F.P.; Brunet, D.; Paula, R.R.; et al. Nitrogen cycling in monospecific and mixed-species plantations of Acacia mangium and Eucalyptus at 4 sites in Brazil. For. Ecol. Manag. 2019, 436, 56–67. [Google Scholar] [CrossRef]
- Manghabati, H.; Kohlpaintner, M.; Ettl, R.; Mellert, K.; Blum, U.; Göttlein, A. Correlating phosphorus extracted by simple soil extraction methods with foliar phosphorus concentrations of Picea abies (L.) H. Karst. and Fagus sylvatica (L.). J. Plant Nutr. Soil Sci. 2018, 181, 547–556. [Google Scholar] [CrossRef]
- Lamontagne, M.; Adégbidi, H.G.; Assamoi, A.J. Organic fertilization of Christmas tree (Abies balsamea (L.) Mill.) plantations with poultry manure in northwestern New Brunswick, Canada. For. Chron. 2019, 95, 39–49. [Google Scholar] [CrossRef]
- Ferreira, E.B.; Cavalcanti, P.P.; Nogueira, D.A. ExpDes: An R package for ANOVA and experimental designs. Appl. Math. 2014, 5, 2952–2958. [Google Scholar] [CrossRef]
- Achat, D.L.; Pousse, N.; Nicolas, M.; Augusto, L. Nutrient remobilization in tree foliage as affected by soil nutrients and leaf life span. Ecol. Monogr. 2018, 88, 408–428. [Google Scholar] [CrossRef]
- De Souza Kulmann, M.S.; Arruda, W.S.; Vitto, B.B.; de Souza, R.O.S.; Berghetti, Á.L.P.; Tarouco, C.P.; Araújo, M.M.; Nicoloso, F.T.; Schumacher, M.V.; Brunetto, G. Morphological and physiological parameters influence the use efficiency of nitrogen and phosphorus by Eucalyptus seedlings. New For. 2021, 53, 431–448. [Google Scholar] [CrossRef]
- Zhao, Q.; Zeng, D.H. Nitrogen addition effects on tree growth and soil properties mediated by soil phosphorus availability and tree species identity. For. Ecol. Manag. 2019, 449, 117478. [Google Scholar] [CrossRef]
- Lima Neto, A.J.; Neves, J.C.L.; Martinez, H.E.P.; Sousa, J.S.; Fernandes, L.V. Nutrient accumulation and nutritional efficiency in eucalyptus. J. Plant Nutr. 2021, 44, 2421–2434. [Google Scholar] [CrossRef]
- Morais, E.G.; Silva, C.A. Novel slow-release NPK biochar-based fertilizers with acidulated apatite: Evaluation of the fertilization value in a short-term experiment. J. Soil Sci. Plant Nutr. 2023, 23, 4937–4954. [Google Scholar] [CrossRef]
- De Souza Kulmann, M.S.; Stefanello, L.O.; Arruda, W.S.; Sans, G.A.; Parcianello, C.F.; Hindersmann, J.; Berghetti, A.L.P.; Araujo, M.M.; Gatiboni, L.C.; Brunetto, G. Nitrogen supply methods affect the root growth dynamics in Eucalyptus grandis. For. Ecol. Manag. 2020, 473, 118320. [Google Scholar] [CrossRef]
- Chatzistathis, T.; Kavvadias, V.; Sotiropoulos, T.; Papadakis, I.E. Organic fertilization and tree orchards. Agriculture 2021, 11, 692. [Google Scholar] [CrossRef]
- Jaquetti, R.K.; Nascimento, H.E.M.; Zotarelli, L.; Rathinasabapathi, B.; Carvalho, J.F.G. Coordinated adjustments of carbohydrates and growth of tree legumes under different fertilization regimes in degraded areas in Amazonia. New For. 2022, 53, 221–240. [Google Scholar] [CrossRef]
- Andivia, E.; Villar-Salvador, P.; Oliet, J.A.; Puertolas, J.; Dumroese, R.K.; Ivetić, V.; Molina-Venegas, R.; Arellano, E.C.; Ovalle, J.F. Climate and species stress resistance modulate the higher survival of large seedlings in forest restorations worldwide. Ecol. Appl. 2021, 31, e02394. [Google Scholar] [CrossRef]
- Pillay, T.; Ngcobo, S.; Ward, D. Nutrient addition, fire and grass competition affects biological nitrogen fixation in Vachellia sieberiana, and associated soil respiration. Pedobiologia 2023, 96, 150848. [Google Scholar] [CrossRef]
- Shi, Z.; Bai, Z.; Guo, D.; Li, S.; Chen, M. Species Diversity and Soil Interconstraints Exert Significant Influences on Plant Survival during Ecological Restoration in Semi-Arid Mining Areas. Diversity 2023, 15, 1100. [Google Scholar] [CrossRef]
- Sloan, J.L.; Uscola, M.; Jacobs, D.F. Nitrogen recovery in planted seedlings, competing vegetation, and soil in response to fertilization on a boreal mine reclamation site. For. Ecol. Manag. 2016, 360, 60–68. [Google Scholar] [CrossRef]
- Maciel, J.C.; Duque, T.S.; Ferreira, E.A.; Zanuncio, J.C.; Plata-Rueda, A.; Silva, V.P.; dos Santos, J.B. Growth, Nutrient Accumulation, and Nutritional Efficiency of a Clonal Eucalyptus Hybrid in Competition with Grasses. Forests 2022, 13, 1157. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH Water | P | K | Ca | Mg | Al | T | m | V | CO | Sand | Silt | Clay |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1:2.5 | -mg dm−3- | ----cmolc dm−3---- | --%-- | ------------g kg−1----------- | ||||||||
| 6.0 | 0.4 | 15.9 | 2.5 | 0.4 | 0.02 | 7.1 | 1 | 41 | 1.0 | 460 | 230 | 310 |
| MF–OF | N | P | K | Ca | Mg | S | CO |
|---|---|---|---|---|---|---|---|
| --%-- | -------------------------mg dm−3------------------------- | -----g dm−3----- | |||||
| 0–0 | 0–0 | 0–0 | 0–0 | 0–0 | 0–0 | 0–0 | 0–0 |
| 100–0 | 50–0 | 150–0 | 50–0 | 383–0 | 50–0 | 0.3–0 | 0–0 |
| 75–25 | 38–39 | 113–10 | 38–15 | 330–110 | 45–15 | 0.5–0.2 | 0.0–0.7 |
| 50–50 | 25–78 | 75–20 | 25–35 | 206–206 | 35–35 | 0.4–0.4 | 0–1.4 |
| 25–75 | 13–117 | 38–30 | 13–58 | 105–317 | 19–58 | 0.3–0.8 | 0–2.1 |
| 0–100 | 0–155 | 0–40 | 0–87 | 0–437 | 0–87 | 0–1.3 | 0–2.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Araújo, F.V.d.; Duque, T.S.; Ferreira, E.A.; Pereira, I.M.; Souza, I.M.; Oliveira, F.S.; dos Santos, J.B. Nutrient Allocation and Growth Responses of Senegalia polyphylla under Varied Fertilizer Regimes for Effective Forest Recovery. Plants 2024, 13, 2420. https://doi.org/10.3390/plants13172420
Araújo FVd, Duque TS, Ferreira EA, Pereira IM, Souza IM, Oliveira FS, dos Santos JB. Nutrient Allocation and Growth Responses of Senegalia polyphylla under Varied Fertilizer Regimes for Effective Forest Recovery. Plants. 2024; 13(17):2420. https://doi.org/10.3390/plants13172420
Chicago/Turabian StyleAraújo, Fillipe Vieira de, Tayna Sousa Duque, Evander Alves Ferreira, Israel Marinho Pereira, Iasmim Marcella Souza, Fernanda Santos Oliveira, and José Barbosa dos Santos. 2024. "Nutrient Allocation and Growth Responses of Senegalia polyphylla under Varied Fertilizer Regimes for Effective Forest Recovery" Plants 13, no. 17: 2420. https://doi.org/10.3390/plants13172420
APA StyleAraújo, F. V. d., Duque, T. S., Ferreira, E. A., Pereira, I. M., Souza, I. M., Oliveira, F. S., & dos Santos, J. B. (2024). Nutrient Allocation and Growth Responses of Senegalia polyphylla under Varied Fertilizer Regimes for Effective Forest Recovery. Plants, 13(17), 2420. https://doi.org/10.3390/plants13172420