
Transcriptome and Metabolome Analyses Reveal the Molecular Mechanisms of Albizia odoratissima’s Response to Drought Stress

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
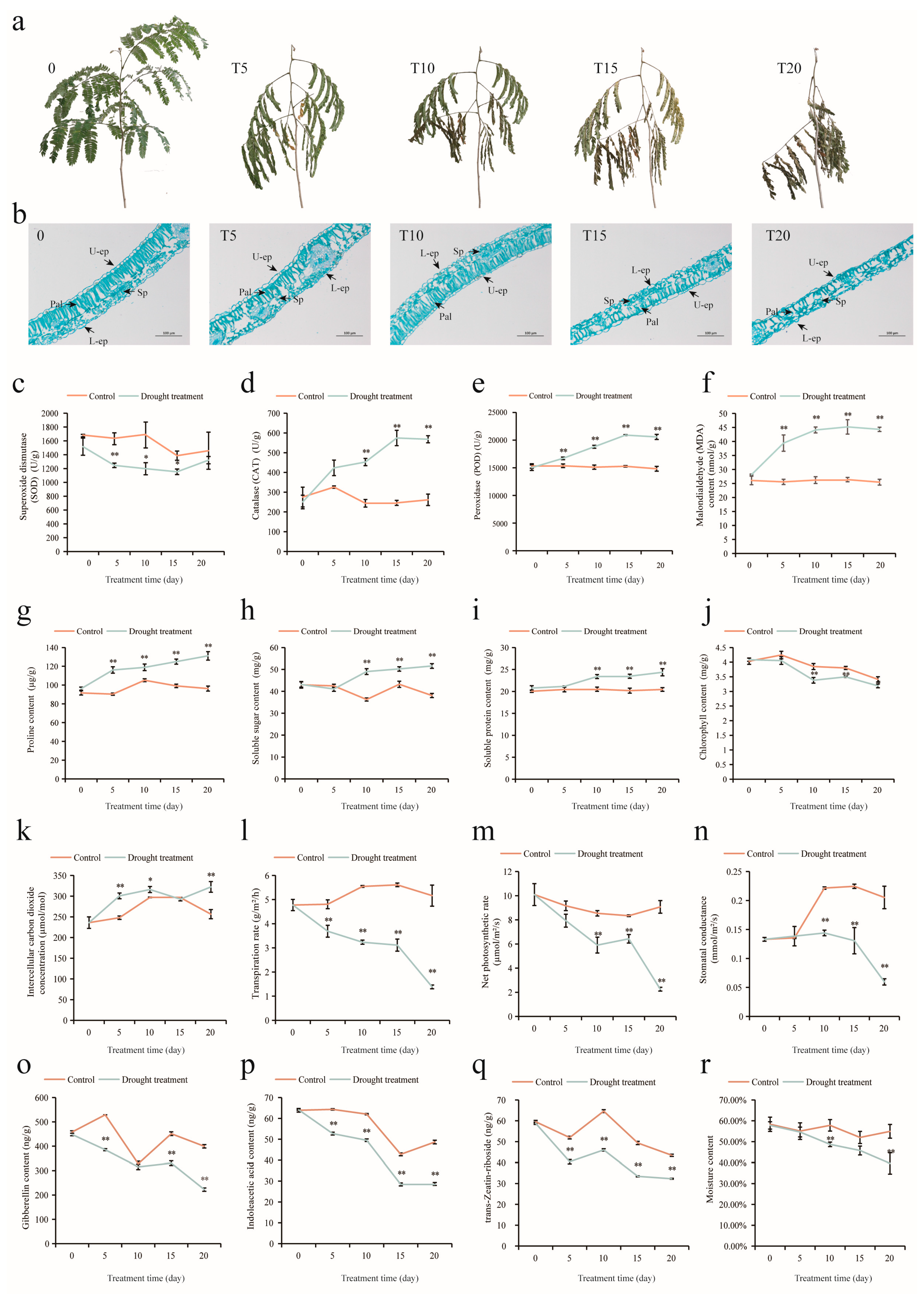
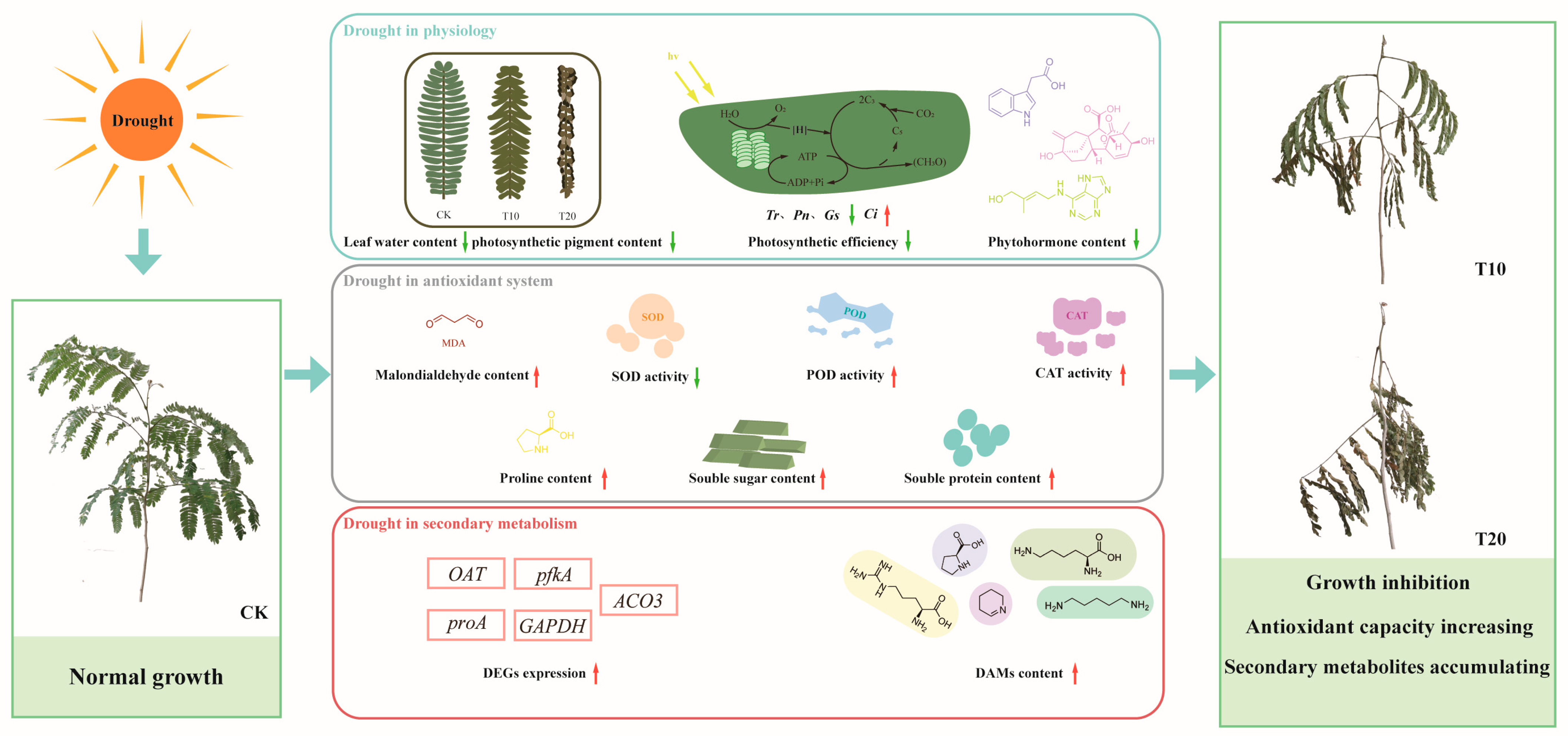
2.1. Structural and Physiological Changes in A. odoratissima Seedlings under Drought Stress
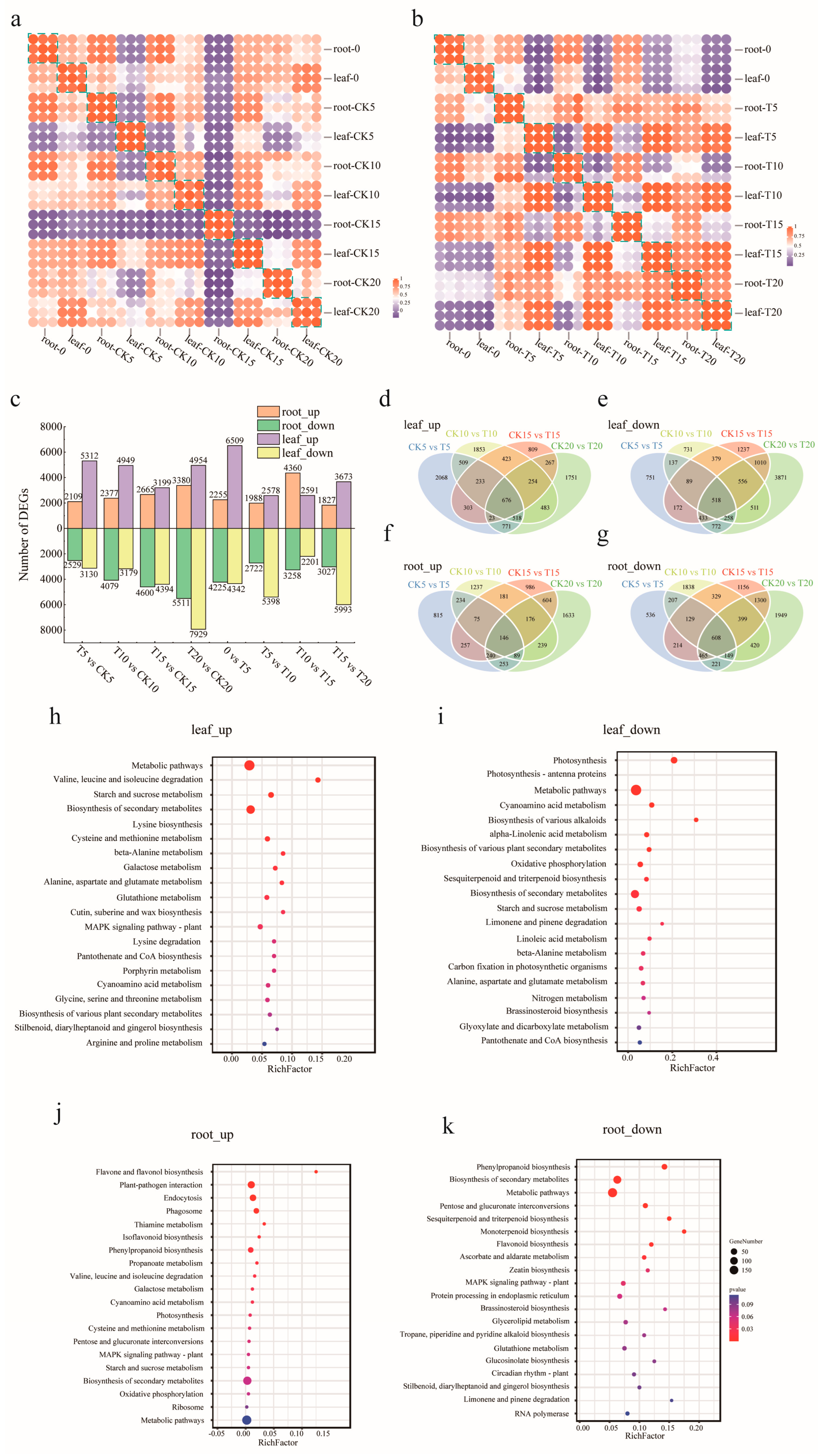
2.2. Transcriptome Analysis of A. odoratissima Seedlings under Drought Stress
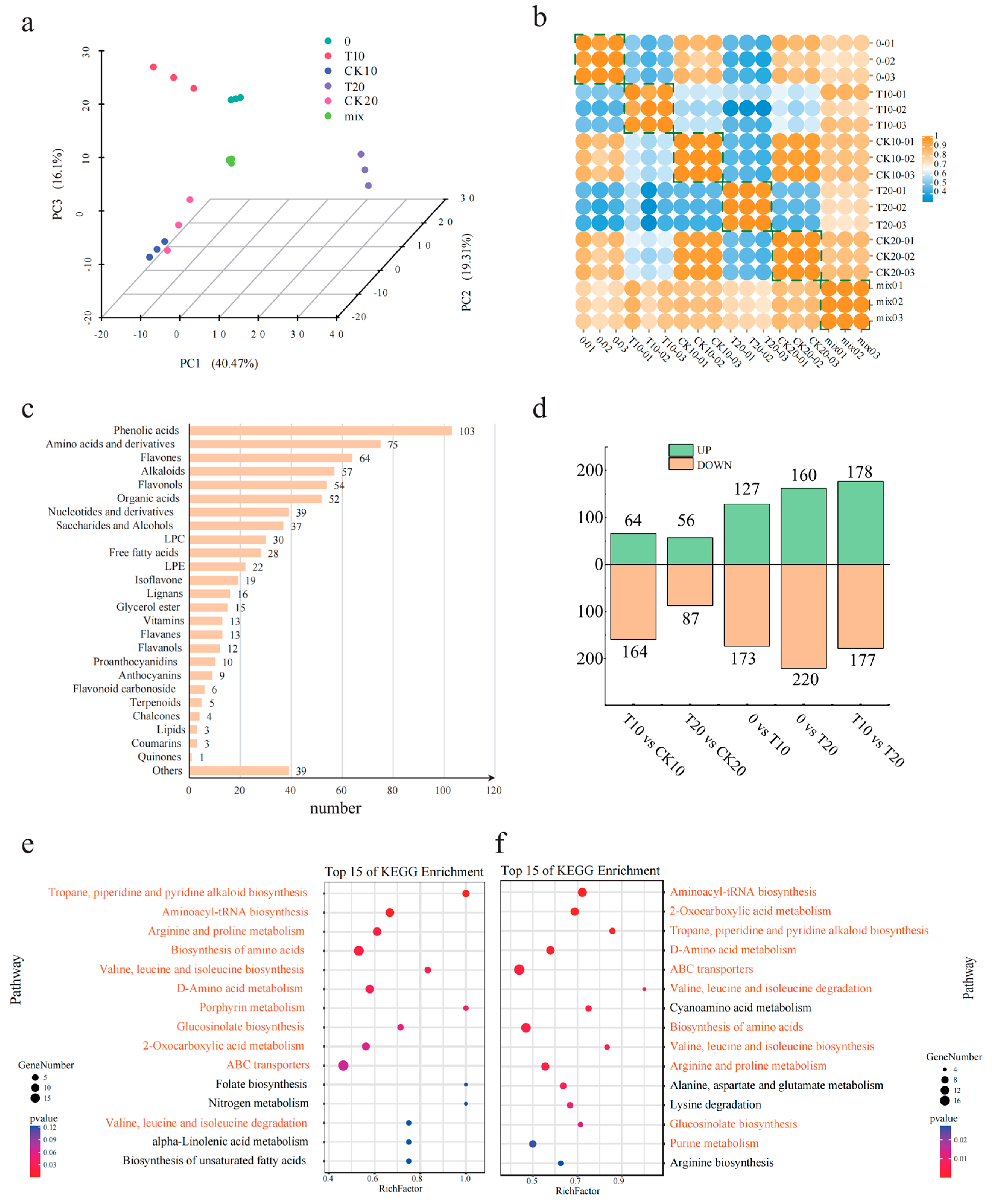
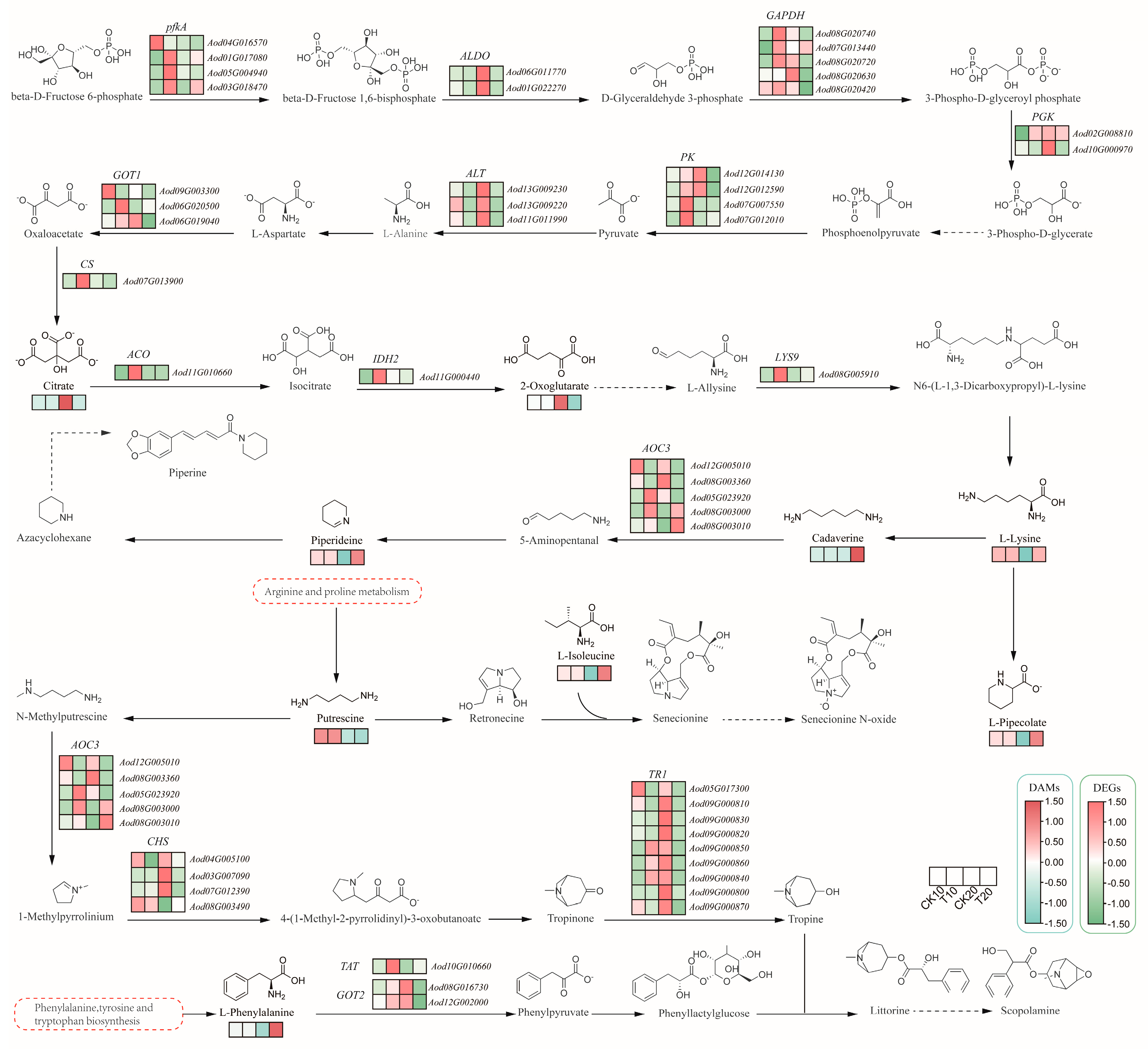
2.3. Metabolome Analysis of A. odoratissima Seedlings under Drought Stress
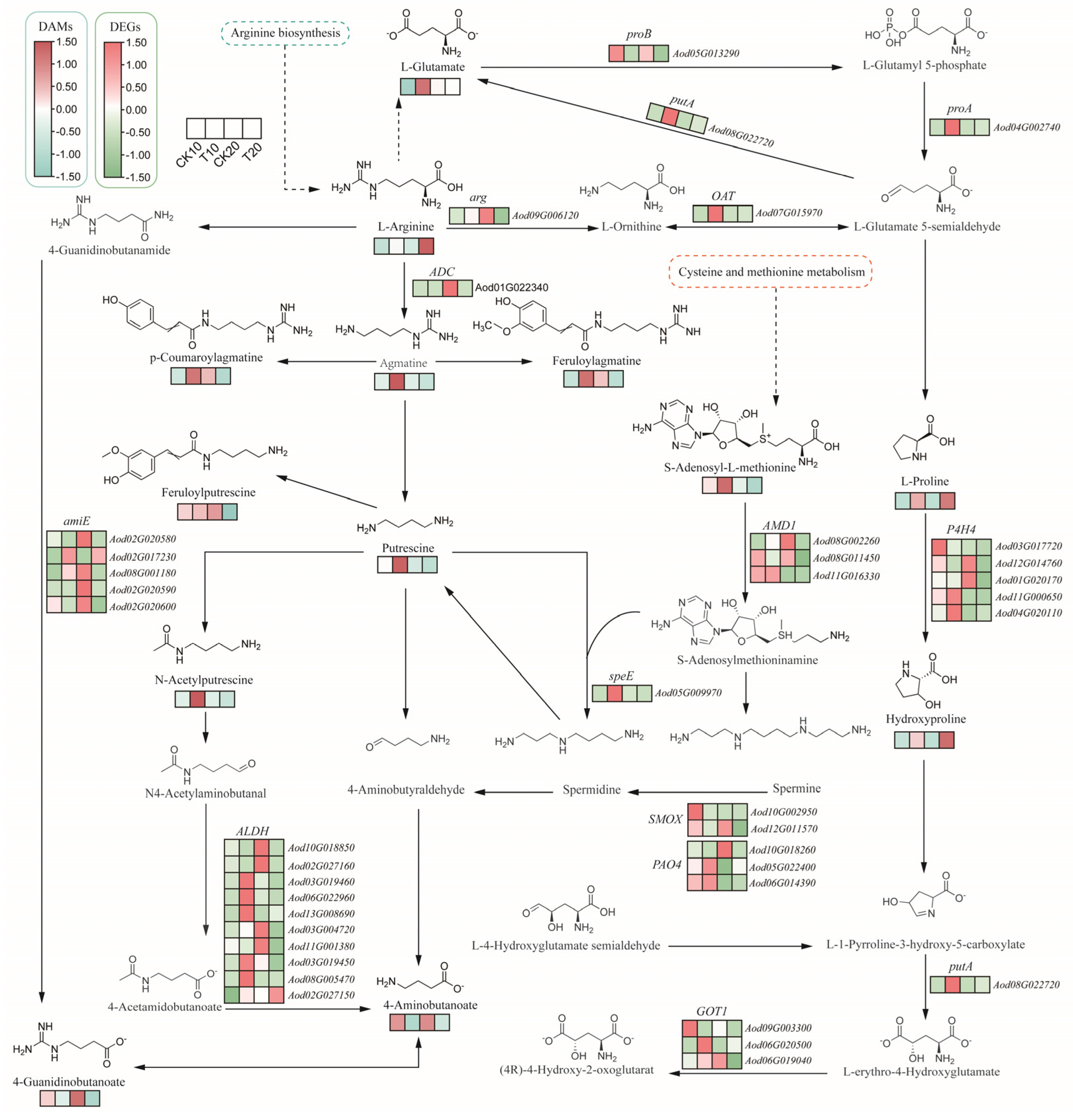
2.4. Integrated Transcriptome and Metabolome Analysis of A. odoratissima Seedlings under Drought Stress
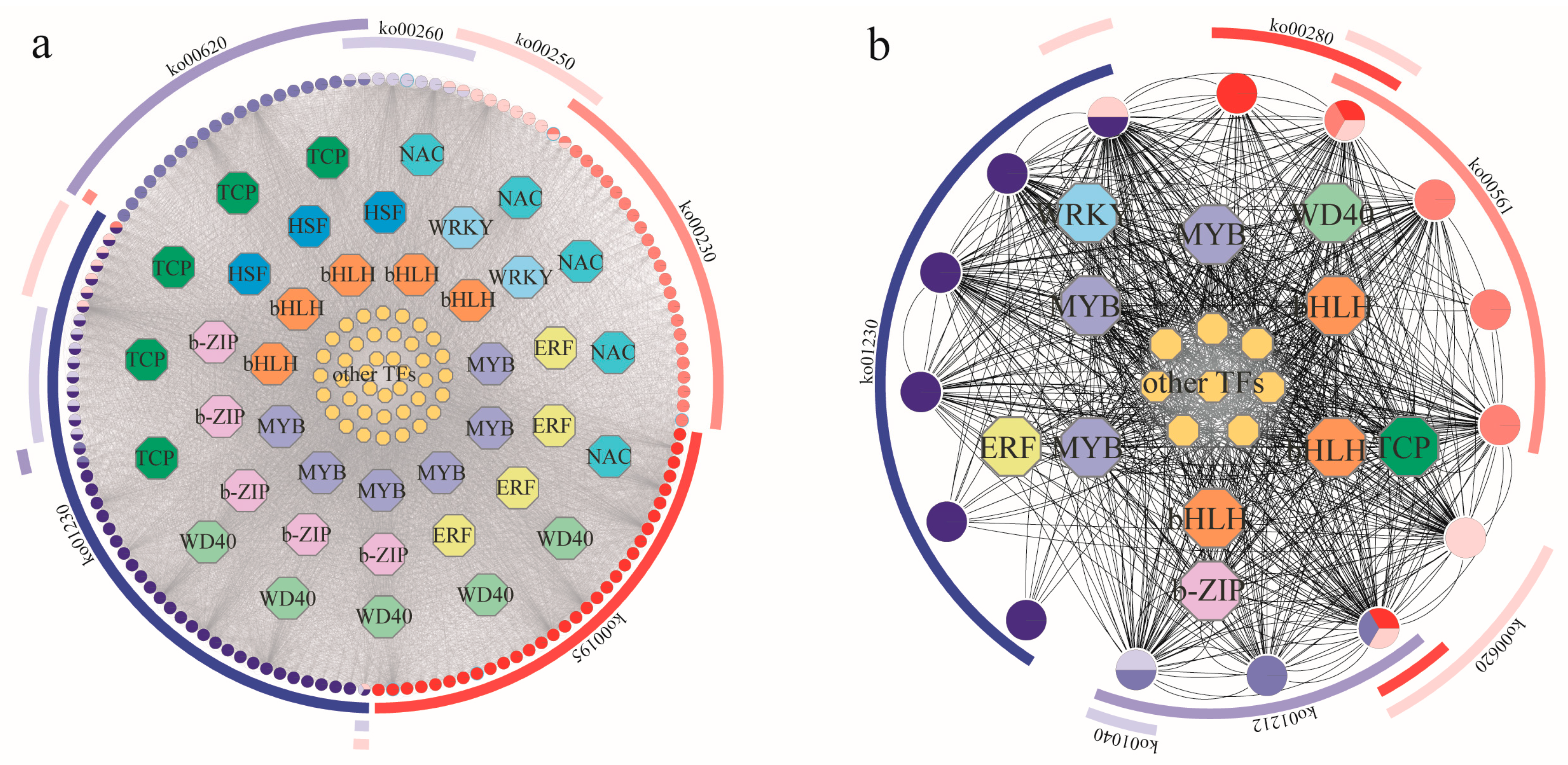
2.5. Weighted Gene Co-Expression Network Analysis of A. odoratissima under Drought Stress
3. Discussion
3.1. Drought Stress Inhibits the Growth of A. odoratissima Seedlings
3.2. Drought Stress Improves the Antioxidant Capacity of A. odoratissima Seedlings
3.3. Drought Stress Promotes the Accumulation of Amino Acids and Alkaloids
3.4. Identification of Key Transcription Factors in Response to Drought Stress
4. Materials and Methods
4.1. Design of Experiments
4.1.1. Drought-Stressed Environment
4.1.2. Pre-Experiment
4.1.3. Drought Stress Experiment
4.2. Measurement of Physiological Indicators
4.2.1. Total Chlorophyll Content in the Leaves
4.2.2. Photosynthetic Parameters
4.2.3. Osmosis-Regulating Substances
4.2.4. Enzyme Activity
4.2.5. Relative Water Content in the Leaves
4.2.6. Endogenous Hormones
4.3. Transcriptome Analysis
4.4. Metabolome Analysis
4.5. Integrated Transcriptome and Metabolome Analysis
4.6. WGCNA Co-Expression Network Construction
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, H.; Sun, X.; Dai, M. Improving crop drought resistance with plant growth regulators and rhizobacteria: Mechanisms, applications, and perspectives. Plant Commun. 2022, 3, 100228. [Google Scholar] [CrossRef]
- Ashok, P.; Jaykumar, R.; Kumar, A.P. Physiological and metabolic adjustments in the xero-halophyte Haloxylon salicornicum conferring drought tolerance. Physiol. Plant. 2021, 172, 1189–1211. [Google Scholar] [CrossRef]
- Rodrigues, J.; Inzé, D.; Nelissen, H.; Saibo, N.J. Source-sink regulation in crops under water deficit. Trends Plant Sci. 2019, 24, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Jin, H.-D.; Cao, X.-Y.; Deng, W.-H.; Du, L.-J. Physiological response to drought stress and drought resistance of six Helleborus orientlis cultivars. J. Appl. Ecol. 2023, 34, 2644–2654. [Google Scholar] [CrossRef]
- Lawson, T.; Oxborough, K.; Morison, J.; Baker, N.R. The responses of guard and mesophyll cell photosynthesis to CO2, O2, light and water stress in a range of species are similar. J. Exp. Bot. 2003, 54, 1743–1752. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M. Inducing drought tolerance in plants: Recent advances. Biotechnol. Adv. 2010, 28, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Yang, J.; Duan, W.; Ma, X.; Qu, L.; Xu, Z.; Yang, Y.; Xu, J. NtRAV4 negatively regulates drought tolerance in Nicotiana tabacum by enhancing antioxidant capacity and defence system. Plant Cell Rep. 2022, 41, 1775–1788. [Google Scholar] [CrossRef]
- Li, J.; Guo, W.; Zhao, J.; Meng, H.; Yang, Y.; Zheng, G.; Yuan, W. Transcriptional Regulation of the Acer truncatum B. Response to Drought and the Contribution of AtruNAC36 to Drought Tolerance. Antioxidants 2023, 12, 1339. [Google Scholar] [CrossRef]
- Zhao, M.; Zhang, Q.; Liu, H.; Tang, S.; Shang, C.; Zhang, W.; Sui, Y.; Zhang, Y.; Zheng, C.; Zhang, H.; et al. The osmotic stress-activated receptor-like kinase DPY1 mediates SnRK2 kinase activation and drought tolerance in Setaria. Plant Cell 2023, 35, 3782–3808. [Google Scholar] [CrossRef]
- Khan, R.; Ma, X.; Zhang, J.; Wu, X.; Iqbal, A.; Wu, Y.; Zhou, L.; Wang, S. Circular drought-hardening confers drought tolerance via modulation of the antioxidant defense system, osmoregulation, and gene expression in tobacco. Physiol. Plant. 2021, 172, 1073–1088. [Google Scholar] [CrossRef] [PubMed]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Meyer, E.; Aspinwall, M.J.; Lowry, D.B.; Palacio-Mejía, J.D.; Logan, T.L.; Fay, P.A.; Juenger, T.E. Integrating transcriptional, metabolomic, and physiological responses to drought stress and recovery in switchgrass (Panicum virgatum L.). BMC Genom. 2014, 15, 527. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sarma, S.J.; Sumner, L.W.; Jones, A.D.; Last, R.L. Switchgrass metabolomics reveals striking genotypic and developmental differences in specialized metabolic phenotypes. J. Agric. Food Chem. 2022, 70, 8010–8023. [Google Scholar] [CrossRef] [PubMed]
- Tiedge, K.; Muchlinski, A.; Zerbe, P. Genomics-enabled analysis of specialized metabolism in bioenergy crops: Current progress and challenges. Synth. Biol. 2020, 5, ysaa005. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Lv, G. Combined analysis of transcriptome and metabolome reveals the molecular mechanism and candidate genes of Haloxylon drought tolerance. Front. Plant Sci. 2022, 13, 1020367. [Google Scholar] [CrossRef]
- Jiao, P.; Liu, T.; Zhao, C.; Fei, J.; Guan, S.; Ma, Y. ZmTCP14, a TCP transcription factor, modulates drought stress response in Zea mays L. Environ. Exp. Bot. 2023, 208, 105232–105244. [Google Scholar] [CrossRef]
- Liang, Q.; Han, Y.; Qiao, Y.; Xie, K.; Li, S.; Dong, Y.; Li, S.; Zhang, S. Effects of drought stress on the growth and physiological characteristics of Sect. Aigeiros clones. J. Beijing For. Univ. 2023, 45, 81–89. [Google Scholar]
- Xu, Y.; Hu, W.; Song, S.; Ye, X.; Ding, Z.; Liu, J.; Wang, Z.; Li, J.; Hou, X.; Xu, B.; et al. MaDREB1F confers cold and drought stress resistance through common regulation of hormone synthesis and protectant metabolite contents in banana. Hortic. Res. 2023, 10, uhac275. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, C.; Li, Z.; Xin, F. Physiological characteristics and anatomical structure of typical shrubs in semi-arid valley areas of Tibet in response to drought stress. J. Northeast For. Univ. 2023, 51, 81–88. [Google Scholar] [CrossRef]
- Wei, S.; Liang, R.; Lin, J.; He, Y.; Jiang, Y.; Ou, H.; Wei, H. Geographical distribution and community characteristics of Albizia odoratissima in China. Guangxi For. Sci. 2020, 49, 71–75. [Google Scholar] [CrossRef]
- Wei, H.; Wei, S.; Jiang, Y.; Liang, R.; Ou, H. Analysis of growth differences in seedlings of three different provenances of Albizia odoratissima. Chin. J. Trop. Agric. 2020, 40, 16–21. [Google Scholar]
- Jiang, Y.; Wei, S.; Lin, J.; Ou, H.; Liang, R. Analysis of seed phenotypic traits and growth differences of different provenances of Albizia odoratissima. Guangxi For. Sci. 2020, 49, 66–70. [Google Scholar] [CrossRef]
- Liu, G.; Ou, H.B.; Wei, Q.S.; Lu, Z.F.; Wei, Y.X.; Chen, D.Y.; Li, Z.G.; Li, S.H. Effects of Drought Stress on Growth and Photosynthetic Characteristics of Albizia odoratissima Seedlings. Guangxi For. Sci. 2023, 52, 181–185. [Google Scholar] [CrossRef]
- Li, S. Mechanism Study on the Effects of Drought on Leaf Photosynthesis and CO2 Diffusion. Doctoral Dissertation, Chinese Academy of Agricultural Sciences, Beijing, China, 2021. [Google Scholar]
- Rizhsky, L.; Liang, H.; Mittler, R. The combined effect of drought stress and heat shock on gene expression in tobacco. Plant Physiol. 2002, 130, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Yuan, Y.; Zhang, J.; Shi, Z. Effects of different water stress methods on chlorophyll fluorescence dynamics of walnut seedlings. J. Hebei Agric. Univ. 2003, 26, 20–24. [Google Scholar] [CrossRef]
- Wu, Z.; Yu, L.; Yan, L.; Zhou, C.; Cai, G.; Zhang, J. Response of leaf anatomical structure and photosynthetic characteristics of Clematis terniflora to drought stress. J. Southern Agric. 2018, 49, 1156–1163. [Google Scholar] [CrossRef]
- Wei, Y.; Ji, Y.; Jiang, S.; Zhang, Y.; Zhao, L. Effects of drought stress and sandstorm on photosynthesis of Caragana korshinskii. J. Arid Land Resour. Environ. 2022, 36, 136–140. [Google Scholar] [CrossRef]
- Jin, E.J.; Yoon, J.-H.; Lee, H.; Bae, E.J.; Yong, S.H.; Choi, M.S. Evaluation of drought stress level in Sargent’s cherry (Prunus sargentii Rehder) using photosynthesis and chlorophyll fluorescence parameters and proline content analysis. PeerJ 2023, 11, e15954. [Google Scholar] [CrossRef]
- Anwar, T.; Shehzadi, A.; Qureshi, H.; Shah, M.N.; Danish, S.; Salmen, S.H.; Ansari, M.J. Alleviation of cadmium and drought stress in wheat by improving growth and chlorophyll contents amended with GA3 enriched deashed biochar. Sci. Rep. 2023, 28, 18503. [Google Scholar] [CrossRef]
- Liu, H.; Song, S.; Liu, M.; Mu, Y.; Li, Y.; Xuan, Y.; Niu, L.; Zhang, H.; Wang, W. Transcription Factor ZmNAC20 Improves Drought Resistance by Promoting Stomatal Closure and Activating Expression of Stress-Responsive Genes in Maize. Int. J. Mol. Sci. 2023, 24, 4712. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, W.; Lv, Y.; Li, T.; Tang, J.; Yang, X.; Bai, J.; Jin, X.; Zhou, H. Effects of drought stress during critical periods on the photosynthetic characteristics and production performance of Naked oat (Avena nuda L.). Sci. Rep. 2022, 12, 11199. [Google Scholar] [CrossRef] [PubMed]
- Al-Yasi, H.; Attia, H.; Alamer, K.; Hassan, F.; Ali, E.; Elshazly, S.; Siddique, K.H.; Hessini, K. Impact of drought on growth, photosynthesis, osmotic adjustment, and cell wall elasticity in Damask rose. Plant Physiol. Biochem. 2020, 150, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Zhao, Y.; Xie, H.; Shi, Y.; Xie, X.; Chai, A.; Li, L.; Li, B. Selection and evaluation of suitable reference genes for quantitative gene expression analysis during infection of Cucumis sativus with Pectobacterium brasiliense. J. Appl. Microbiol. 2022, 132, 3717–3734. [Google Scholar] [CrossRef] [PubMed]
- Abraham, B. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Du, L.; Huang, X.; Ding, L.; Wang, Z.; Tang, D.; Chen, B.; Ao, L.; Liu, Y.; Kang, Z.; Mao, H. TaERF87 and TaAKS1 synergistically regulate TaP5CS1/TaP5CR1-mediated proline biosynthesis to enhance drought tolerance in wheat. New Phytol. 2023, 237, 232–250. [Google Scholar] [CrossRef] [PubMed]
- Javid, M.G.; Sorooshzadeh, A.; Moradi, F.; Modarres Sanavy, S.A.M.; Allahdadi, I. The role of phytohormones in alleviating salt stress in crop plants. Austral. J. Crop Sci. 2011, 5, 726–734. [Google Scholar]
- Zheng, Q.Z.; Tan, H.Y.; Gao, X.; Ji, X.F.; Tian, X. Effects of drought, salt and their combined stress on physiology and biochemistry characteristics of hulless barley seedlings. Jiangsu Ari Sci. 2020, 48, 97–103. [Google Scholar] [CrossRef]
- Lv, L.; Chen, X.; Li, H.; Huang, J.; Liu, Y.; Zhao, A. Different adaptive patterns of wheat with different drought tolerance under drought stresses and rehydration revealed by integrated metabolomic and transcriptomic analysis. Front. Plant Sci. 2022, 13, 1008624. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.; Jiang, C.; Chen, L.; Paul, A.; Chatterjee, A.; Shen, G. Achieving abiotic stress tolerance in plants through antioxidative defense mechanisms. Front. Plant Sci. 2023, 14, 1110622. [Google Scholar] [CrossRef]
- Lu, Q.J.; Liang, C.; Huang, S.W.; Zeng, X.B.; Huang, K.T.; Zhang, S.J. Effect of combined aluminium and manganese stress on chlorophyll and membrane lipid peroxidation in Helianthus annuus seedlings. Anhui Agric. Sci. Bull. 2021, 27, 27–28+37. [Google Scholar] [CrossRef]
- Li, F.; Chen, G.; Xie, Q.; Zhou, S.; Hu, Z. Down-regulation of SlGT-26 gene confers dwarf plants and enhances drought and salt stress resistance in tomato. Plant Physiol. Biochem. 2023, 203, 108053. [Google Scholar] [CrossRef]
- Hildebrandt, T.M.; Nunes, N.A.; Araújo, W.L.; Braun, H.-P. Amino Acid Catabolism in Plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef]
- Pratelli, R.; Pilot, G. Regulation of amino acid metabolic enzymes and transporters in plants. J. Exp. Bot. 2014, 65, 5535–5556. [Google Scholar] [CrossRef] [PubMed]
- Ashrafi, M.; Azimi-Moqadam, M.-R.; Moradi, P.; MohseniFard, E.; Shekari, F.; Kompany-Zareh, M. Effect of drought stress on metabolite adjustments in drought tolerant and sensitive thyme. Plant Physiol. Biochem. 2018, 132, 391–399. [Google Scholar] [CrossRef]
- Zhao, X.; Huang, L.J.; Sun, X.F.; Zhao, L.L.; Wang, P.C. Transcriptomic and Metabolomic Analyses Reveal Key Metabolites, Pathways and Candidate Genes in Sophora davidii (Franch.) Skeels Seedlings under Drought Stress. Front. Plant Sci. 2022, 13, 785702. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Li, B.; Guan, S.; Jia, J.; Xu, X.; Zhang, Z.; Lu, Z.; Li, X.; Pang, X. EuRBG10 involved in indole alkaloids biosynthesis in Eucommia ulmoides induced by drought and salt stresses. J. Plant Physiol. 2022, 278, 153813. [Google Scholar] [CrossRef] [PubMed]
- Malekzadeh, P. Influence of exogenous application of glycinebetaine on antioxidative system and growth of salt-stressed soybean seedlings (Glycine max L.). Physiol. Mol. Biol. Plants 2015, 21, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Shan, T.; Jin, P.; Zhang, Y.; Huang, Y.; Wang, X.; Zheng, Y. Exogenous glycine betaine treatment enhances chilling tolerance of peach fruit during cold storage. Postharv. Biol. Technol. 2016, 114, 104–110. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’aversana, E.; Carillo, P. Spatial and Temporal Profile of Glycine Betaine Accumulation in Plants Under Abiotic Stresses. Front. Plant Sci. 2019, 10, 230. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Morgil, H.; Tardu, M.; Cevahir, G.; Kavakli, İ.H. Comparative RNA-seq analysis of the drought-sensitive lentil (Lens culinaris) root and leaf under short- and long-term water deficits. Funct. Integr. Genom. 2019, 19, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Li, N.; Fan, Y.; Dong, B.; Song, Z.; Cao, H.; Du, T.; Liu, T.; Qi, M.; Niu, L.; et al. Transcriptome analysis reveals abscisic acid enhancing drought resistance by regulating genes related to flavonoid metabolism in Pigeon pea. Environ. Exp. Bot. 2021, 191, 104627. [Google Scholar] [CrossRef]
- Sun, X.; Xu, Y.; Zhang, Q.; Li, X.; Yan, Z. Combined effect of water inundation and heavy metals on the photosynthesis and physiology of Spartina alterniflora. Ecotoxicol. Environ. Saf. 2018, 153, 248–258. [Google Scholar] [CrossRef]
- Wang, J.; Xu, W.; Su, Y.; Zhang, B. The effect of drought stress on drought resistance in different germplasms of alfalfa during the seedling stage. Guizhou Agric. Sci. 2023, 51, 14–24. [Google Scholar] [CrossRef]
- Senthilkumar, M.; Amaresan, N.; Sankaranarayanan, A. Estimation of malondialdehyde (MDA) by thiobarbituric acid (TBA) assay. In Plant-Microbe Interactions; Springer Protocols Handbooks; Humana Press: New York, NY, USA, 2021; pp. 103–105. [Google Scholar]
- Chance, B.; Maehly, A.C. Assay of catalase and peroxidases. Methods Enzymol. 1995, 2, 764–775. [Google Scholar] [CrossRef]
- Zhang, C.; Shi, S. Physiological and Proteomic Responses of Contrasting Alfalfa (Medicago sativa L.) Varieties to PEG-Induced Osmotic Stress. Front. Plant Sci. 2018, 9, 242. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Yang, C.; Tan, S.; Gu, Z.; Tang, S.; Yu, F. Effects of drought stress on proline and endogenous hormone content in Pinus massoniana seedlings of different provenances. South China For. Sci. 2020, 48, 24–28,53. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, S.; Gao, F.; Wang, Z.; Yin, G.; Wen, S.; Ou, H.; Liu, Z. Transcriptome and Metabolome Analyses Reveal the Molecular Mechanisms of Albizia odoratissima’s Response to Drought Stress. Plants 2024, 13, 2732. https://doi.org/10.3390/plants13192732
Wei S, Gao F, Wang Z, Yin G, Wen S, Ou H, Liu Z. Transcriptome and Metabolome Analyses Reveal the Molecular Mechanisms of Albizia odoratissima’s Response to Drought Stress. Plants. 2024; 13(19):2732. https://doi.org/10.3390/plants13192732
Chicago/Turabian StyleWei, Shuoxing, Feng Gao, Zhihui Wang, Guoping Yin, Shizhi Wen, Hanbiao Ou, and Zhiming Liu. 2024. "Transcriptome and Metabolome Analyses Reveal the Molecular Mechanisms of Albizia odoratissima’s Response to Drought Stress" Plants 13, no. 19: 2732. https://doi.org/10.3390/plants13192732
APA StyleWei, S., Gao, F., Wang, Z., Yin, G., Wen, S., Ou, H., & Liu, Z. (2024). Transcriptome and Metabolome Analyses Reveal the Molecular Mechanisms of Albizia odoratissima’s Response to Drought Stress. Plants, 13(19), 2732. https://doi.org/10.3390/plants13192732