
Effect of Picloram and Desiccation on the Somatic Embryogenesis of Lycium barbarum L.


Abstract
:1. Introduction
2. Results and Discussion
2.1. Effects of Explants and Plant Growth Regulators on Callus Induction and Somatic Embryogenesis
2.2. Somatic Embryo Development, Germination, and Plantlet Regeneration
2.3. Abnormal Embryos
2.4. Acclimatization
2.5. Cytochemical Staining
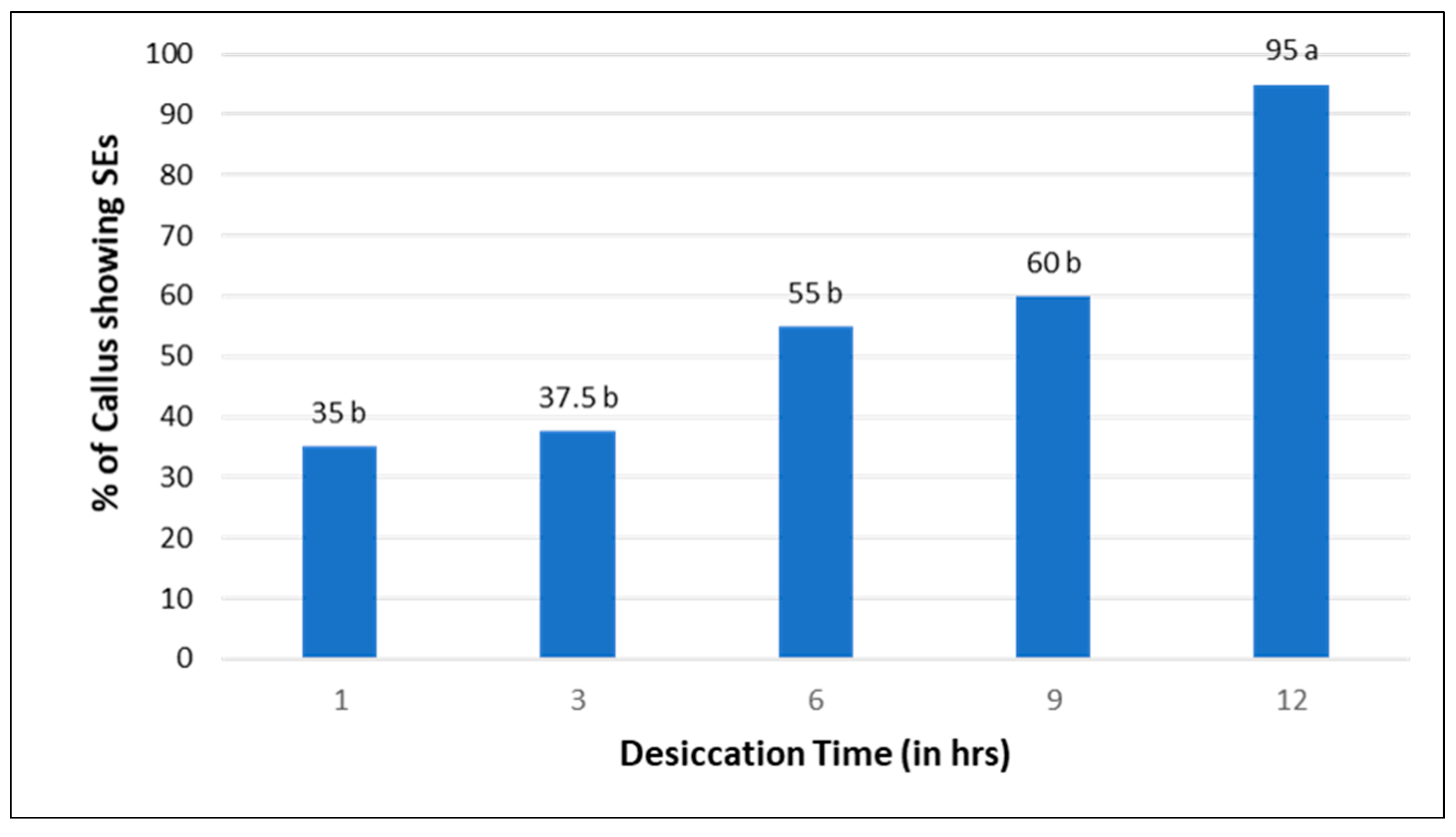
2.6. Effect of Desiccation Treatment on Somatic Embryo Induction
3. Materials and Methods
3.1. Plant Materials
3.2. Media Preparation and Culture Conditions
3.3. Induction of Embryogenic Callus and Somatic Embryo Formation
3.4. Cytochemical Analysis of Callus
3.5. Effect of Desiccation Treatment on Somatic Embryo Induction
3.6. Acclimatization, Hardening, and Greenhouse Transfer of Plants
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Felzenszwalb, I.; Da Costa Marques, M.R.; Mazzei, J.L.; Aiub, C.A.F. Toxicological evaluation of Euterpe edulis: A potential superfruit to be considered. Food Chem. Toxicol. 2013, 58, 536. [Google Scholar] [CrossRef]
- Zhang, D.; Xia, T.; Dang, S.; Fan, G.; Wang, Z. Investigation of Chinese wolfberry (Lycium spp.) germplasm by restriction site-associated DNA sequencing (RAD-seq). Biochem. Genet. 2018, 56, 575–585. [Google Scholar] [CrossRef]
- Vidović, B.B.; Milinčić, D.D.; Marčetić, M.D.; Djuriš, J.D.; Ilić, T.D.; Kostić, A.Ž.; Pešić, M.B. Health benefits and applications of goji berries in functional food products development: A review. Antioxidants 2022, 11, 248. [Google Scholar] [CrossRef]
- Potterat, O. Goji (Lycium barbarum and L. chinense): Phytochemistry, pharmacology and safety in the perspective of traditional uses and recent popularity. Planta Med. 2010, 7, 7–19. [Google Scholar] [CrossRef]
- Huang, D.F.; Tang, Y.F.; Nie, S.P.; Wan, Y.; Xie, M.Y.; Xie, X.M. Effect of phenylethanoid glycosides and polysaccharides from the seed of Plantago asiatica L. on the maturation of murine bone marrow-derived dendritic cells. Eur. J. Pharmacol. 2009, 620, 105–111. [Google Scholar] [CrossRef]
- Xu, X.; Shan, B.; Liao, C.H.; Xie, J.H.; Wen, P.W.; Shi, J.Y. Anti-diabetic properties of Momordica charantia L. polysaccharide in alloxan-induced diabetic mice. Int. J. Biol. Macromol. 2015, 81, 538–543. [Google Scholar] [CrossRef]
- Ma, R.H.; Zhang, X.X.; Ni, Z.J.; Thakur, K.; Wang, W.; Yan, Y.M.; Cao, Y.L.; Zhang, J.G.; Rengasamy, K.R.R.; Wei, Z.J. Lycium barbarum (goji) as functional food: A review of its nutrition, phytochemical structure, biological features, and food industry prospects. Crit. Rev. Food Sci. Nutr. 2022, 145, 10621–10635. [Google Scholar] [CrossRef]
- Wang, L.; Li, J.A.; Wei, L.J.; Bai, D.M.; Liu, B.C. Lycium barbarum polysaccharide with potential anti-gastric cancer effects mediated by regulation of miR-202-5p/PIK3CA. Arab. J. Chem. 2022, 15, 104162. [Google Scholar] [CrossRef]
- Silvestri, C.; Sabbatini, G.; Marangelli, F.; Rugini, E.; Cristofori, V. Micropropagation and ex vitro rooting of Wolfberry. HortScience 2018, 53, 494–1499. [Google Scholar] [CrossRef]
- Raemakers, C.; Jacobsen, E.; Visser, R.G.F. Secondary somatic embryogenesis and applications in plant breeding. Euphytica 1995, 81, 93–107. [Google Scholar] [CrossRef]
- Quiroz-Figueroa, F.R.; Rojas Herrera, R.; Galaz-Avalos, R.M.; Loyola-Vargas, V.M. Embryo production through somatic embryogenesis can be used to study cell differentiation in plants. Plant Cell Tissue Organ Cult. 2006, 86, 258–301. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Ratushnyak, Y.I.; Piven, N.M.; Rudas, V.A. Protoplast culture and plant regeneration in Lycium barbarum L. Plant Cell Tissue Organ Cult. 1989, 17, 183–190. [Google Scholar] [CrossRef]
- Gamborg, O.; Miller, R.; Ojimi, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Ratushnyak, Y.I.; Rudas, V.A.; Piven, N.M. Regeneration of Lycium barbarum L. plants from leaf tissue, callus culture and callus protoplast. Plant Cell Rep. 1990, 9, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Guo, G.Q.; Zhao, D.L.; Li, L.H.; Zheng, G.C. Shoot regeneration from leaf explant of Lycium barbarum and Agrobacterium-mediated genetic transformation. Russ. J. Plant Physiol. 2001, 48, 453–458. [Google Scholar] [CrossRef]
- Hu, Z.; Yang, J.; Guo, G.Q.; Zheng, G.C. High efficiency transfomation of Lycium barbarum and Agrobacterium tumefaciens and transgenic plant regeneration via somatic embryogenesis. Plant Cell Rep. 2002, 21, 233–237. [Google Scholar]
- Hu, Z.; Wang, Y.X.; Wu, Y.R.; Li, W. Effects of light and plant growth regulators on growth of normal and hairy root from Lycium barbarum in vitro. China J. Materia Med. 2006, 31, 106–110. [Google Scholar]
- Hu, Z.; Hu, Y.; Gao, H.H.; Guan, X.Q.; Zhuang, D.H. Callus production, somatic embryogenesis, and plant regeneration of Lycium barbarum root explants. Biol. Plant. 2008, 52, 93–96. [Google Scholar] [CrossRef]
- Dãnãilã-Guidea, S.M.; Dobrinoiu, R.V.; Vişan, L.; Toma, R.C. Protocol for efficient in vitro multiplication of Lycium barbarum L. (goji) by direct organogenesis. Sci. Bull. Ser. F Biotechnol. 2015, 19, 34–38. [Google Scholar]
- Fira, A.; Joshee, N.; Cristea, V.; Simu, M.; Hârța, M.; Pamfil, D.; Clapa, D. Optimization of micropropagation protocol for goji berry (Lycium barbarum L.). Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca Horticulture 2016, 73, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Osman, N.I.; Awal, A.; Sidik, N.J.; Abdullah, S. Callus induction and somatic embryogenesis from leaf and nodal explants of Lycium barbarum L. (Goji). Biotechnology 2013, 12, 36–45. [Google Scholar] [CrossRef]
- Ahmed, M.A. In vitro propagation and improving accumulation of coumarin in Lycium barbarum, a rare plant in the flora of Egypt. Bull. Natl. Res. Cent. 2022, 46, 220. [Google Scholar] [CrossRef]
- Verma, S.K.; Gantait, S.; Mukherjee, E.; Gurel, E. Enhanced somatic embryogenesis, plant regeneration and total phenolic content estimation in Lycium barbarum L.: A highly nutritive and medicinal plant. J. Crop Sci. Biotechnol. 2022, 25, 547–555. [Google Scholar] [CrossRef]
- Ruta, C.; De Mastro, G.; Ancona, S.; Tagarelli, A.; De Cillis, F.; Benelli, C.; Lambardi, M. Large-scale plant production of Lycium barbarum L. by liquid culture in temporary immersion system and possible application to the synthesis of bioactive substance. Plants 2020, 9, 844. [Google Scholar] [CrossRef] [PubMed]
- Anwar, F.; Sharmila, P.; Saradhi, P.P. No more recalcitrant: Chickpea regeneration and genetic transformation. Afr. J. Biotechnol. 2010, 9, 782–797. [Google Scholar]
- Tang, Y.; Liu, B.; Li, X.M.; Li, J.; Li, H.X. Endogenous hormone concentrations in explants and calluses of bitter melon (Momordica charantia L.). Interciencia 2010, 35, 680–683. [Google Scholar]
- Joshee, N.; Biswas, B.K.; Yadav, A.K. Somatic embryogenesis and plant development in Centella asiatica L., a Highly prized medicinal plant of the tropics. HortScience 2007, 42, 633–637. [Google Scholar] [CrossRef]
- Kordestani, G.K.; Karami, O. Picloram- induced somatic embryogenesis in leaves of strawberry (Fragaria ananassa L.). Acta Biol. Crac. Ser. Bot. 2008, 50, 69–72. [Google Scholar]
- Barro, F.; Martin, A.; Lazzeri, P.A.; Barcelo, P. Medium optimization for efficient somatic embryogenesis and plant regeneration from immature inflorescences and immature scutella of elite cultivars of wheat, barley and tridordeum. Euphytica 1999, 108, 161–167. [Google Scholar] [CrossRef]
- Fehér, A.; Pasternak, T.P.; Dudits, D. Transition of somatic plant cells to an embryogenic state. Plant Cell Tissue Organ Cult. 2003, 74, 201–228. [Google Scholar] [CrossRef]
- Lelu-Walter, M.A.; Gautier, F.; Eliášová, K.; Sanchez, L.; Teyssier, C.; Lomenech, A.M.; Le Mette, C.; Hargreaves, C.; Trontin, J.F.; Reeves, C. High gellan gum concentration and secondary somatic embryogenesis: Two key factors to improve somatic embryo development in Pseudotsuga menziesii [Mirb.]. Plant Cell Tissue Organ Cult. 2018, 132, 137–155. [Google Scholar] [CrossRef]
- Atak, M.; Kaya, M.; Khawar, M.M.; Saglam, S.; Özcan, S.; Ciftci, C.Y. Effect of age on somatic embryogenesis from immature zygotic embryos of 5 Turkish triticale genotypes. Afr. J. Biotechnol. 2008, 7, 1765–1768. [Google Scholar]
- Pinto, G.; Park, Y.S.; Silva, S.; Neves, L.; Araújo, C.; Santos, C. Factors affecting maintenance, proliferation, and germination of secondary somatic embryos of Eucalyptus globulus Labill. Plant Cell Tissue Organ Cult. 2008, 95, 69–78. [Google Scholar] [CrossRef]
- Bahgat, S.; Shabban, O.A.; El-Shihy, O.; Lightfoot, D.; El-Shemy, H.A. Establishment of the regeneration system for Vicia faba L. Curr. Issues Mol. Biol. 2008, 11, 47–54. [Google Scholar]
- Garcia, C.; Furtado de Almeida, A.A.; Costa, M.; Britto, D.; Valle, R.; Royaert, S.; Marell, J.P. Abnormalities in somatic embryogenesis caused by 2,4-D: An overview. Plant Cell Tissue Organ Cult. 2019, 137, 193–212. [Google Scholar] [CrossRef]
- Gupta, P.K.; Durzan, D.J. Biotechnology of somatic polyembryogenesis and plantlet regeneration in loblolly pine. Nat. Biotechnol. 1987, 5, 147–151. [Google Scholar] [CrossRef]
- Pádua, M.S.; Paiva, L.V.; Silva, L.C.D.; Livramento, K.D.; Alves, E.; Castro, A.H.F. Morphological characteristics, and cell viability of coffee plants calli. Rural Sci. 2014, 44, 660–665. [Google Scholar] [CrossRef]
- Siddique, A.B.; Ara, I.; Islam, S.M.S.; Tuteja, N. Effect of air desiccation and salt stress factors on in vitro regeneration of rice (Oryza sativa L.). Plant Signal. Behav. 2014, 9, e977209. [Google Scholar] [CrossRef]
- Rance, I.M.; Tian, W.; Mathews, H.; Kochko, A.D.; Beachy, R.N.; Fauquet, C. Partial desiccation of mature embryo-derived calli, a simple treatment that dramatically enhances the regeneration ability of indica rice. Plant Cell Rep. 1994, 13, 647–651. [Google Scholar] [CrossRef]
- Rahman, Z.A.; Suri Roowi, W.Z.W.S.; Subramaniam, S. Regeneration of Malaysian indica rice (Oryza sativa) variety MR232 via optimized somatic embryogenesis system. J. Phytol. 2010, 2, 30–38. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Explants | Media Compositions | Plant Regeneration | References |
|---|---|---|---|
| Leaf and young internodes | MS with 0.2 mg/L NAA and 1 mg/L 2,4-D MS: Murashige and Skoog [12] | Shoot Organogenesis | [13] |
| Leaf, callus, and callus protoplast | B5 medium [14] containing 1.5 mg/L (BAP) and 0.5 mg/L NAA | Shoot Organogenesis | [15] |
| Leaf | MS medium supplemented with 2 mg/L BA and 0.5 mg/L NAA | Shoot Organogenesis | [16] |
| Leaf | MS medium 1 mg/L IBA (Indole 3 Butyric Acid) and 1 mg/L IBA IAA | Somatic Embryogenesis | [17,18] |
| Root | MS medium containing 0.2 mg/L 2,4-D | Indirect Somatic Embryogenesis | [19] |
| Nodal Segment | DKW (Driver and Kuniyuki Walnut) medium supplemented with 6-benzylaminopurine (BAP; 0.5 mg/L) and sucrose 3% w/v | Micropropagation and Ex vitro Rooting | [11] |
| Leaves, Apexes and Nodal segments | Liquid MS Medium with 1 mg/L IBA | Shoot Organogenesis | [20] |
| Leaf and nodal segments | MS Medium with 1.33 and 2.22 µM Benzyl adenine, gelled with wheat starch as an agar alternative | Shoot Organogenesis | [21] |
| Leaf and Nodal segments | MS Medium with 0.3 mg/L 2,4-D and 0.1–0.3 mg/L BAP | Somatic Embryogenesis | [22] |
| Shoot tips | MS medium supplemented with 225.24 µM BAP | Shoot Organogenesis | [23] |
| Hypocotyl | MS Medium with 0.25 mg/L 2,4-D + 1 mg/L TDZ | Somatic Embryogenesis | [24] |
| Axillary bud | 0.1 mg/L BAP + Liquid basal medium in temporary immersion system (TIS) using bioreactor PlantformTM | In vitro micropropagation (Large scale shoot production) | [25] |
| Treatments | Picloram (µM) | 2,4-D (µM) | Responsive Explant | Somatic Embryo Induction |
|---|---|---|---|---|
| 1 | Control | - | - | |
| 2 | 0.1 | Leaf | Somatic embryos | |
| 3 | 1.0 | Leaf | Somatic embryos | |
| 4 | 10.0 | Leaf and Leaf with petiole | Callus | |
| 5 | 0.1 | Leaf | Somatic embryos | |
| 6 | 1.0 | Leaf and Leaf with petiole | Callus | |
| 7 | 10.0 | leaf | Callus | |
| 8 | 0.05 | 0.05 | Leaf | Somatic embryos |
| 9 | 0.5 | 0.5 | leaf | Callus |
| 10 | 5.0 | 5.0 | leaf | Callus |
| Treatments | Picloram (µM) | 2,4-D (µM) | BAP (µM) | Mean Number of Somatic Embryos per Explant |
|---|---|---|---|---|
| 1 (Control) | 0.00 | 0.00 | 0.00 | 0.00 ± 0.00 a |
| 2 | 0.1 | 1.0 | 1.35 ± 0.13 ef | |
| 5.0 | 5.45 ± 0.40 c | |||
| 10.0 | 5.65 ± 0.63 c | |||
| 3 | 1.0 | 1.0 | 7.80 ± 0.50 b | |
| 5.0 | 4.00 ± 0.47 d | |||
| 10.0 | 10.40 ± 0.47 a | |||
| 4 | 0.1 | 1.0 | 4.05 ± 0.51 d | |
| 5.0 | 1.65 ± 0.34 ef | |||
| 10.0 | 2.55 ± 0.41 e | |||
| 5 | 0.05 | 0.05 | 1.0 | 1.15 ± 0.29 f |
| 5.0 | 1.60 ± 0.31 ef | |||
| 10.0 | 1.10 ± 0.29 f |
| No. | Stages | Procedure and Media Composition | Conditions and Durations | Results |
|---|---|---|---|---|
| 1 | Culture establishment | Leaves and leaves with petioles explants were immersed in 70% (v/v) ethanol for 1 min, followed by immersion in 15% sodium hypochlorite solution for 10 min and then rinsed three times. | All steps were performed under laminar airflow cabinet. | Plant materials were surface sterilized. |
| 2 | Callus induction | MS basal medium [12] Picloram, 2,4-D and Picloram + 2,4-D (0.1 µM, 1.0 µM and 10.0 µM). | 25 ± 2 °C Darkness 5 weeks (1 subculture). Developing cultures were examined at weekly intervals using a microscope (Nikon SMZ 1270). | Nodular and friable embryogenic calluses were initiated. |
| 3 | Stabilization of embryogenic callus | MS basal medium BAP (1.0 µM, 5.0 µM and 10.0 µM). | 25 ± 2 °C; 14 h light 60 μmol m−2 s −1; 6 weeks (2 subcultures). | Embryogenic calluses were selected for further proliferation. Somatic embryos were differentiated. |
| 4 | Development of somatic embryos | MS basal medium, 30 g L−1 sucrose. | 25 ± 2 °C; 14 h light 60 μmol m−2 s−1; 7–8 weeks (3 subcultures). | Calluses with plenty of embryos were obtained. |
| 5 | Germination of somatic embryos | MS basal medium, 30 g L−1 sucrose. | 25 ± 2 °C; 14 h light at 60 μmol m−2 s −1 until plantlet regeneration. | Further conversion of embryos and plantlet regeneration was obtained. Germinated bipolar embryos with well-formed shoots and roots were used. |
| 6 | Acclimatization of plantlets | Germinated plants were washed from agar and planted into humidity-controlled boxes. Promix BX, Premier Horticulture Inc., Pennsylvania was used. | 25 ± 2 °C; 16 h light 60 μmol m−2 s−1; 21 days (about 3 weeks) until new growth was visible. | Plants were acclimatized into humidity-controlled boxes. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khatri, P.; Joshee, N. Effect of Picloram and Desiccation on the Somatic Embryogenesis of Lycium barbarum L. Plants 2024, 13, 151. https://doi.org/10.3390/plants13020151
Khatri P, Joshee N. Effect of Picloram and Desiccation on the Somatic Embryogenesis of Lycium barbarum L. Plants. 2024; 13(2):151. https://doi.org/10.3390/plants13020151
Chicago/Turabian StyleKhatri, Poonam, and Nirmal Joshee. 2024. "Effect of Picloram and Desiccation on the Somatic Embryogenesis of Lycium barbarum L." Plants 13, no. 2: 151. https://doi.org/10.3390/plants13020151