

Fine-Root Distribution and Soil Physicochemical Property Variations in Four Contrasting Urban Land-Use Types in South Korea

 , ,
, ,  ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Variation in Soil Properties by Land-Use Type
2.2. Edaphic Drivers of Fine-Root Mass
2.3. Variations in Standing Fine-Root Biomass and Necromass across Four Land-Use Types
2.4. Vertical Distribution of Fine-Root Biomass and Necromass in Four Land-Use Types
3. Discussion
3.1. Variation in Soil Physical and Chemical Properties across Land-Use Types
3.2. Variation in Standing Root Mass and Fine-Root Vertical Distribution across Land-Use Types
3.3. Implications for Sustainable Urban Planning
4. Materials and Methods
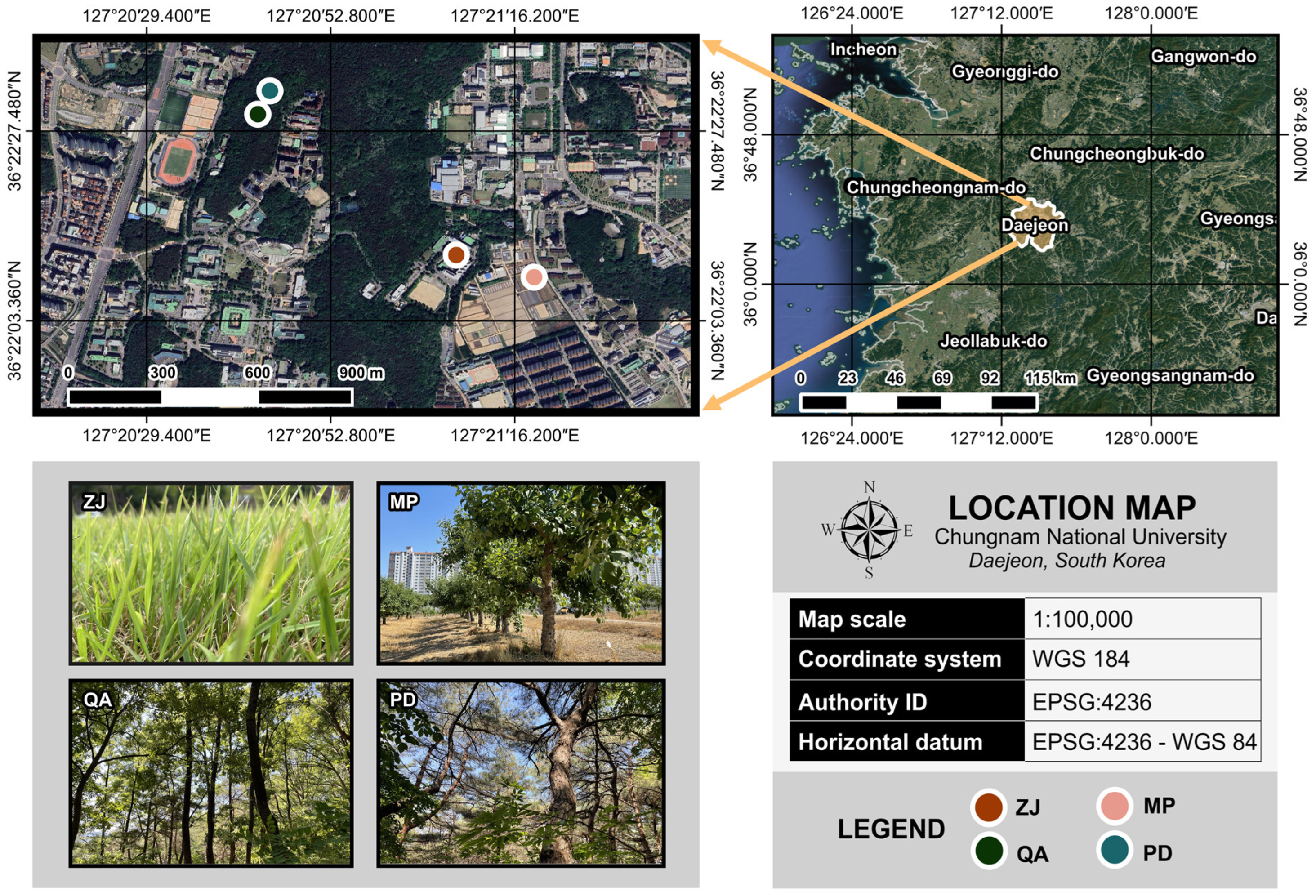
4.1. Study Site
4.2. Soil Sampling
4.3. Fine-Root Mass Inventory
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Creutzig, F.; d’Amour, C.B.; Weddige, U.; Fuss, S.; Beringer, T.; Gläser, A.; Kalkuhl, M.; Steckel, J.C.; Radebach, A.; Edenhofer, O. Assessing Human and Environmental Pressures of Global Land-Use Change 2000–2010. Glob. Sustain. 2019, 2, e1. [Google Scholar] [CrossRef]
- United Nations. World Urbanization Prospects, the 2011 Revision. In Population Division, Department of Economic and Social Affairs, United Nations Secretariat; United Nations: New York, NY, USA, 2014. [Google Scholar]
- Wei, H.; Guo, P.; Zheng, H.; He, X.; Wang, P.; Ren, Z.; Zhai, C. Micro-Scale Heterogeneity in Urban Forest Soils Affects Fine Root Foraging by Ornamental Seedlings of Buddhist Pine and Northeast Yew. Urban For. Urban Green. 2017, 28, 63–72. [Google Scholar] [CrossRef]
- Upadhyay, S.; Raghubanshi, A.S. Chapter 16—Determinants of Soil Carbon Dynamics in Urban Ecosystems; Verma, P., Singh, P., Singh, R., Raghubanshi, A.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 299–314. ISBN 978-0-12-820730-7. [Google Scholar]
- IPCC. Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2019. [Google Scholar]
- DeFries, R.S.; Foley, J.A.; Asner, G.P. Land-Use Choices: Balancing Human Needs and Ecosystem Function. Front. Ecol. Environ. 2004, 2, 249–257. [Google Scholar] [CrossRef]
- Leul, Y.; Assen, M.; Damene, S.; Legass, A. Effects of Land Use Types on Soil Quality Dynamics in a Tropical Sub-Humid Ecosystem, Western Ethiopia. Ecol. Indic. 2023, 147, 110024. [Google Scholar] [CrossRef]
- Ramesh, T.; Bolan, N.S.; Kirkham, M.B.; Wijesekara, H.; Kanchikerimath, M.; Rao, C.S.; Sandeep, S.; Rinklebe, J.; Ok, Y.S.; Choudhury, B.U. Soil Organic Carbon Dynamics: Impact of Land Use Changes and Management Practices: A Review. Adv. Agron. 2019, 156, 1–107. [Google Scholar]
- Cornejo, N.S.; Becker, J.N.; Hemp, A.; Hertel, D. Effects of Land-Use Change and Disturbance on the Fine Root Biomass, Dynamics, Morphology, and Related C and N Fluxes to the Soil of Forest Ecosystems at Different Elevations at Mt. Kilimanjaro (Tanzania). Oecologia 2023, 201, 1089–1107. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-G.; Kirschbaum, M.U.F.; Eichler-Löbermann, B.; Gifford, R.M.; Liáng, L.L. The Effect of Land-Use Change on Soil C, N, P, and Their Stoichiometries: A Global Synthesis. Agric. Ecosyst. Environ. 2023, 348, 108402. [Google Scholar] [CrossRef]
- Jackson, R.B.; Mooney, H.A.; Schulze, E.-D. A Global Budget for Fine Root Biomass, Surface Area, and Nutrient Contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef]
- Chapin, F.S.; Eviner, V.T. 8.06—Biogeochemistry of Terrestrial Net Primary Production. In Treatise on Geochemistry, 2nd ed.; Holland, H.D., Turekian, K.K., Eds.; Elsevier: Oxford, UK, 2007; pp. 1–35; 189–216. ISBN 978-0-08-043751-4. [Google Scholar]
- Van Do, T.; Sato, T.; Saito, S.; Kozan, O. Fine-Root Production and Litterfall: Main Contributions to Net Primary Production in an Old-Growth Evergreen Broad-Leaved Forest in Southwestern Japan. Ecol. Res. 2015, 30, 921–930. [Google Scholar] [CrossRef]
- Norby, R.J.; Jackson, R.B. Root Dynamics and Global Change: Seeking an Ecosystem Perspective. New Phytol. 2000, 147, 3–12. [Google Scholar] [CrossRef]
- Sims, P.L.; Singh, J.S. The Structure and Function of Ten Western North American Grasslands: IV. Compartmental Transfers and Energy Flow Within the Ecosystem. J. Ecol. 1978, 66, 983–1009. [Google Scholar] [CrossRef]
- Dong, L.; Berg, B.; Sun, T.; Wang, Z.; Han, X. Response of Fine Root Decomposition to Different Forms of N Deposition in a Temperate Grassland. Soil Biol. Biochem. 2020, 147, 107845. [Google Scholar] [CrossRef]
- Zanotelli, D.; Montagnani, L.; Manca, G.; Scandellari, F.; Tagliavini, M. Net Ecosystem Carbon Balance of an Apple Orchard. Eur. J. Agron. 2015, 63, 97–104. [Google Scholar] [CrossRef]
- Gan, Z.; Zhou, Z.; Liu, W. Vertical Distribution and Seasonal Dynamics of Fine Root Parameters for Apple Trees of Different Ages on the Loess Plateau of China. Agric. Sci. China 2010, 9, 46–55. [Google Scholar] [CrossRef]
- Coll, L.; Camarero, J.J.; Martinez De Aragon, J. Fine Root Seasonal Dynamics, Plasticity, and Mycorrhization in 2 Coexisting Mediterranean Oaks with Contrasting Aboveground Phenology. Ecoscience 2012, 19, 238–245. [Google Scholar] [CrossRef]
- Hao, H.; Wei, Y.; Cao, D.; Guo, Z.; Shi, Z. Vegetation Restoration and Fine Roots Promote Soil Infiltrability in Heavy-Textured Soils. Soil Tillage Res. 2020, 198, 104542. [Google Scholar] [CrossRef]
- Huang, G.; Zhao, X.; Su, Y.; Zhao, H.; Zhang, T. Vertical Distribution, Biomass, Production and Turnover of Fine Roots along a Topographical Gradient in a Sandy Shrubland. Plant Soil 2008, 308, 201–212. [Google Scholar] [CrossRef]
- Xie, C.; Cai, S.; Yu, B.; Yan, L.; Liang, A.; Che, S. The Effects of Tree Root Density on Water Infiltration in Urban Soil Based on a Ground Penetrating Radar in Shanghai, China. Urban For. Urban Green. 2020, 50, 126648. [Google Scholar] [CrossRef]
- Kotowska, M.M.; Leuschner, C.; Triadiati, T.; Hertel, D. Conversion of Tropical Lowland Forest Reduces Nutrient Return through Litterfall, and Alters Nutrient Use Efficiency and Seasonality of Net Primary Production. Oecologia 2016, 180, 601–618. [Google Scholar] [CrossRef]
- Pransiska, Y.; Triadiati, T.; Tjitrosoedirjo, S.; Hertel, D.; Kotowska, M.M. Forest Conversion Impacts on the Fine and Coarse Root System, and Soil Organic Matter in Tropical Lowlands of Sumatera (Indonesia). For. Ecol. Manag. 2016, 379, 288–298. [Google Scholar] [CrossRef]
- Ishaq, M.; Ibrahim, M.; Lal, R. Tillage Effects on Soil Properties at Different Levels of Fertilizer Application in Punjab, Pakistan. Soil Tillage Res. 2002, 68, 93–99. [Google Scholar] [CrossRef]
- Ryadin, A.R.; Janz, D.; Schneider, D.; Tjoa, A.; Irawan, B.; Daniel, R.; Polle, A. Early Effects of Fertilizer and Herbicide Reduction on Root-Associated Biota in Oil Palm Plantations. Agronomy 2022, 12, 199. [Google Scholar] [CrossRef]
- Phillips, R.P.; Fahey, T.J. Fertilization Effects on Fineroot Biomass, Rhizosphere Microbes and Respiratory Fluxes in Hardwood Forest Soils. New Phytol. 2007, 176, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Fukuzawa, K.; Shibata, H.; Takagi, K.; Satoh, F.; Koike, T.; Sasa, K. Temporal Variation in Fine-Root Biomass, Production and Mortality in a Cool Temperate Forest Covered with Dense Understory Vegetation in Northern Japan. For. Ecol. Manag. 2013, 310, 700–710. [Google Scholar] [CrossRef]
- Solly, E.; Schöning, I.; Boch, S.; Müller, J.; Socher, S.A.; Trumbore, S.E.; Schrumpf, M. Mean Age of Carbon in Fine Roots from Temperate Forests and Grasslands with Different Management. Biogeosciences 2013, 10, 4833–4843. [Google Scholar] [CrossRef]
- Raizada, A.; Jayaprakash, J.; Rathore, A.C.; Tomar, J.M.S. Distribution of Fine Root Biomass of Fruit and Forest Tree Species Raised on Old River Bed Lands in the North West Himalaya. Trop. Ecol. 2013, 54, 251–261. [Google Scholar]
- Leuschner, C.; Harteveld, M.; Hertel, D. Consequences of Increasing Forest Use Intensity for Biomass, Morphology and Growth of Fine Roots in a Tropical Moist Forest on Sulawesi, Indonesia. Agric. Ecosyst. Environ. 2009, 129, 474–481. [Google Scholar] [CrossRef]
- Magalhães, T.M.; Mamugy, F.P.S. Fine Root Biomass and Soil Properties Following the Conversion of Miombo Woodlands to Shifting Cultivation Lands. Catena 2020, 194, 104693. [Google Scholar] [CrossRef]
- Pregitzer, K.S. Fine Roots of Trees: A New Perspective. New Phytol. 2002, 154, 267–270. [Google Scholar] [CrossRef]
- Hertel, D.; Harteveld, M.A.; Leuschner, C. Conversion of a Tropical Forest into Agroforest Alters the Fine Root-Related Carbon Flux to the Soil. Soil Biol. Biochem. 2009, 41, 481–490. [Google Scholar] [CrossRef]
- Helmisaari, H.-S.; Lehto, T.; Makkonen, K. Fine Roots and Soil Properties BT—Forest Condition in a Changing Environment: The Finnish Case; Mälkönen, E., Ed.; Springer: Dordrecht, The Netherlands, 2000; pp. 203–217. ISBN 978-94-015-9373-1. [Google Scholar]
- Bengough, A.G.; Mullins, C.E. Mechanical Impedance to Root Growth: A Review of Experimental Techniques and Root Growth Responses. J. Soil Sci. 1990, 41, 341–358. [Google Scholar] [CrossRef]
- Silver, W.L.; Neff, J.; McGroddy, M.; Veldkamp, E.; Keller, M.; Cosme, R. Effects of Soil Texture on Belowground Carbon and Nutrient Storage in a Lowland Amazonian Forest Ecosystem. Ecosystems 2000, 3, 193–209. [Google Scholar] [CrossRef]
- Barrow, N.J.; Hartemink, A.E. The Effects of PH on Nutrient Availability Depend on Both Soils and Plants. Plant Soil 2023, 487, 21–37. [Google Scholar] [CrossRef]
- Chen, L.; Mu, X.; Yuan, Z.; Deng, Q.; Chen, Y.; Yuan, L.Y.; Ryan, L.T.; Kallenbach, R.L. Soil Nutrients and Water Affect the Age-Related Fine Root Biomass but Not Production in Two Plantation Forests on the Loess Plateau, China. J. Arid Environ. 2016, 135, 173–180. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Fine Root Biomass, Production, Turnover Rates, and Nutrient Contents in Boreal Forest Ecosystems in Relation to Species, Climate, Fertility, and Stand Age: Literature Review and Meta-Analyses. CRC. Crit. Rev. Plant Sci. 2010, 29, 204–221. [Google Scholar] [CrossRef]
- Neina, D. The Role of Soil PH in Plant Nutrition and Soil Remediation. Appl. Environ. Soil Sci. 2019, 2019, 1–9. [Google Scholar] [CrossRef]
- Sun, Y.; Bi, H.; Xu, H.; Duan, H.; Peng, R.; Wang, J. Variation of Fine Roots Distribution in Apple (Malus Pumila M.)–Crop Intercropping Systems on the Loess Plateau of China. Agronomy 2018, 8, 280. [Google Scholar] [CrossRef]
- Ge, S.; Zhu, Z.; Jiang, Y. Long-Term Impact of Fertilization on Soil PH and Fertility in an Apple Production System. J. Soil Sci. Plant Nutr. 2018, 18, 282–293. [Google Scholar] [CrossRef]
- Han, S.H.; Kim, S.; Chang, H.; Kim, H.-J.; Khamzina, A.; Son, Y. Soil Depth- and Root Diameter-Related Variations Affect Root Decomposition in Temperate Pine and Oak Forests. J. Plant Ecol. 2019, 12, 871–881. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. In Methods of Soil Analysis; Agronomy Monographs; Wiley: Hoboken, NJ, USA, 1983; pp. 539–579. ISBN 9780891189770. [Google Scholar]
- Park, B.B.; Rahman, A.; Han, S.H.; Youn, W.B.; Hyun, H.J.; Hernandez, J.; An, J.Y. Carbon and Nutrient Inputs by Litterfall in Evergreen and Deciduous Forests in Korea. Forests 2020, 11, 143. [Google Scholar] [CrossRef]
- Uselman, S.M.; Qualls, R.G.; Lilienfein, J. Fine Root Production across a Primary Successional Ecosystem Chronosequence at Mt. Shasta, California. Ecosystems 2007, 10, 703–717. [Google Scholar] [CrossRef]
- Zhou, G.; Meng, S.; Yu, J.; Zhou, H.; Liu, Q. Quantitative Relationships between Fine Roots and Stand Characteristics. Eur. J. For. Res. 2018, 137, 385–399. [Google Scholar] [CrossRef]
- Osono, T.; Azuma, J.; Hirose, D. Plant Species Effect on the Decomposition and Chemical Changes of Leaf Litter in Grassland and Pine and Oak Forest Soils. Plant Soil 2014, 376, 411–421. [Google Scholar] [CrossRef]
- Vang-Petersen, O. Calcium Nutrition of Apple Trees: A Review. Sci. Hortic. 1980, 12, 1–9. [Google Scholar] [CrossRef]
- Park, G.-S.; Kang, G.-N.; Lee, S.-J.; Lee, H.-G.; Lee, S.; Chae, H.-M.; Ohga, S. Net Fine Root Carbon Production in Pinus Densiflora, Pinus Koraiensis, Larix Leptolepis and Quercus Acutissima Stands, Gongju Area, Chungnam Province, Korea; Faculty of Agriculture, Kyushu University: Fukuoka, Japan, 2010. [Google Scholar]
- Han, S.H.; Kim, S.; Chang, H.; Kim, H.; An, J.; Son, Y. Fine Root Biomass and Production Regarding Root Diameter in Pinus Densiflora and Quercus Serrata Forests: Soil Depth Effects and the Relationship with Net Primary Production. Turk. J. Agric. For. 2021, 45, 46–54. [Google Scholar]
- Bueis, T.; Bravo, F.; Pando, V.; Kissi, Y.-A.; Turrión, M.-B. Phosphorus Availability in Relation to Soil Properties and Forest Productivity in Pinus sylvestris L. Plantations. Ann. For. Sci. 2019, 76, 97. [Google Scholar] [CrossRef]
- Havlin, J.L.; Beaton, J.D.; Tisdale, S.L.; Nelson, W.L. An Introduction to Nutrient Management. In Soil Fertility and Fertilizers, 6th ed.; Prentice Hall Up: Saddle River, NJ, USA, 1999; p. 499. [Google Scholar]
- Joslin, J.D.; Gaudinski, J.B.; Torn, M.S.; Riley, W.J.; Hanson, P.J. Fine-Root Turnover Patterns and Their Relationship to Root Diameter and Soil Depth in a 14C-Labeled Hardwood Forest. New Phytol. 2006, 172, 523–535. [Google Scholar] [CrossRef]
- Du, H.; Liu, L.; Su, L.; Zeng, F.; Wang, K.; Peng, W.; Zhang, H.; Song, T. Seasonal Changes and Vertical Distribution of Fine Root Biomass During Vegetation Restoration in a Karst Area, Southwest China. Front. Plant Sci. 2019, 9, 2001. [Google Scholar] [CrossRef]
- Wang, C.; Han, S.; Zhou, Y.; Yan, C.; Cheng, X.; Zheng, X.; Li, M.-H. Responses of Fine Roots and Soil N Availability to Short-Term Nitrogen Fertilization in a Broad-Leaved Korean Pine Mixed Forest in Northeastern China. PLoS ONE 2012, 7, e31042. [Google Scholar] [CrossRef]
- Von Haden, A.C.; Dornbush, M.E. Depth Distributions of Belowground Production, Biomass and Decomposition in Restored Tallgrass Prairie. Pedosphere 2019, 29, 457–467. [Google Scholar] [CrossRef]
- Boeri, P.A.; Unruh, J.B.; Kenworthy, K.E.; Trenholm, L.E.; Rios, E.F. Aboveground and Belowground Traits of Turf-type Bahiagrass (Paspalum notatum Flügge) Genotypes under Simulated Drought. Int. Turfgrass Soc. Res. J. 2022, 14, 276–287. [Google Scholar] [CrossRef]
- Fu, J.; Fry, J.; Huang, B. Growth and Carbon Metabolism of Tall Fescue and Zoysiagrass as Affected by Deficit Irrigation. HortScience 2007, 42, 378–381. [Google Scholar] [CrossRef]
- Song, X.; Gao, X.; Dyck, M.; Zhang, W.; Wu, P.; Yao, J.; Zhao, X. Soil Water and Root Distribution of Apple Tree (Malus Pumila Mill) Stands in Relation to Stand Age and Rainwater Collection and Infiltration System (RWCI) in a Hilly Region of the Loess Plateau, China. Catena 2018, 170, 324–334. [Google Scholar] [CrossRef]
- Li, X.; Minick, K.J.; Li, T.; Williamson, J.C.; Gavazzi, M.; McNulty, S.; King, J.S. An Improved Method for Quantifying Total Fine Root Decomposition in Plantation Forests Combining Measurements of Soil Coring and Minirhizotrons with a Mass Balance Model. Tree Physiol. 2020, 40, 1466–1473. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Xiang, W.; Lei, P.; Deng, X.; Tian, D.; Fang, X.; Peng, C. Standing Fine Root Mass and Production in Four Chinese Subtropical Forests along a Succession and Species Diversity Gradient. Plant Soil 2014, 376, 445–459. [Google Scholar] [CrossRef]
- Jiang, P.; Wang, H.; Fu, X.; Dai, X.; Kou, L.; Wang, J. Elaborate Differences between Trees and Understory Plants in the Deployment of Fine Roots. Plant Soil 2018, 431, 433–447. [Google Scholar] [CrossRef]
- Meinen, C.; Hertel, D.; Leuschner, C. Biomass and Morphology of Fine Roots in Temperate Broad-Leaved Forests Differing in Tree Species Diversity: Is There Evidence of below-Ground Overyielding? Oecologia 2009, 161, 99–111. [Google Scholar] [CrossRef]
- Bourne, K.S.; Conway, T.M. The Influence of Land Use Type and Municipal Context on Urban Tree Species Diversity. Urban Ecosyst. 2014, 17, 329–348. [Google Scholar] [CrossRef]
- Jim, C.Y. Sustainable Urban Greening Strategies for Compact Cities in Developing and Developed Economies. Urban Ecosyst. 2013, 16, 741–761. [Google Scholar] [CrossRef]
- Francos, M.; Bogunovic, I.; Úbeda, X.; Pereira, P. Soil Physico-Chemical Properties and Organic Carbon Stocks across Different Land Use in an Urban Park of Vilnius, Lithuania. J. Cent. Eur. Agric. 2023, 24, 519–530. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Physical Properties | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sites | Sand | Silt | Clay | Soil Texture | SBD | |||||||||
| ZJ | a | b | c | ab | ||||||||||
| 0–10 cm | 87 (0) | 10 (0) | 3 (0.01) | sand | 1.27 (0.07) | |||||||||
| 10–30 cm | 85 (0.02) | 11 (0.02) | 4 (0) | loamy sand | 1.41 (0.07) | |||||||||
| MP | c | a | b | a | ||||||||||
| 0–10 cm | 65 (0.01) | 27 (0.01) | 8 (0.01) | sandy loam | 1.52 (0.09) | |||||||||
| 10–30 cm | 67 (0.01) | 25 (0.01) | 8 (0) | sandy loam | 1.58 (0.07) | |||||||||
| QA | b | b | b | ab | ||||||||||
| 0–10 cm | 78 (0.01) | 13 (0.01) | 9 (0.01) | sandy loam | 1.43 (0.07) | |||||||||
| 10–30 cm | 73 (0.03) | 16 (0.01) | 11 (0.02) | sandy loam | 1.45 (0.09) | |||||||||
| PD | ab | b | a | b | ||||||||||
| 0–10 cm | 73 (0.05) | 13 (0.02) | 13 (0.02) | sandy loam | 1.23 (0.06) | |||||||||
| 10–30 cm | 69 (0.04) | 15 (0.03) | 16 (0.01) | sandy loam | 1.37 (0.09) | |||||||||
| Chemical Properties | ||||||||||||||
| Exchangeable Cations | ||||||||||||||
| pH | EC | OM | AP | TN | C/N | NH4+-N | NO3−-N | CEC | K+ | Na+ | Ca2+ | Mg2+ | Al3+ | |
| ZJ | a | c | c | b | a | b | a | a | b | a | ||||
| 0–10 cm | 6.23 | 0.07 | 2.66 | 121.2 | 0.1 | 12.24 | 2.06 | 16.86 | 3.74 | 0.36 | 0.12 | 2.28 | 0.57 | 0.1 |
| (0.07) | (0.01) | (0.43) | (51.2) | (0.01) | (0.88) | (0.13) | (3.04) | (0.18) | (0.01) | (0.02) | (0.22) | (0.04) | (0.00) | |
| 10–30 cm | 6.23 | 0.05 | 0.91 | 39.4 | 0.02 | 14.16 | 1.73 | 10.23 | 4.41 | 0.23 | 0.09 | 3.19 | 0.56 | 0.1 |
| (0.06) | (0.00) | (0.14) | (25.3) | (0.00) | (1.49) | (0.34) | (2.25) | (0.46) | (0.07) | (0.03) | (0.09) | (0.12) | (0.00) | |
| MP | a | bc | c | b | b | b | a | a | a | a | ||||
| 0–10 cm | 6.9 | 0.23 | 0.23 | 296.5 | 0.06 | 10.63 | 2.25 | 7.6 | 4.92 | 0.31 | 0.08 | 3.31 | 1.25 | 0.1 |
| (0.48) | (0.07) | (0.07) | (240.6) | (0.02) | (0.92) | (0.20) | (2.38) | (0.99) | (0.10) | (0.01) | (0.68) | (0.25) | (0.00) | |
| 10–30 cm | 6.85 | 0.13 | 0.13 | 62.6 | 0.04 | 10.58 | 2.29 | 4.3 | 4.4 | 0.27 | 0.12 | 2.86 | 1.1 | 0.1 |
| (0.46) | (0.02) | (0.02) | (29.7) | (0.01) | (0.94) | (0.34) | (0.34) | (0.20) | (0.05) | (0.03) | (0.06) | (0.06) | (0.00) | |
| QA | b | ab | b | a | b | a | b | b | c | b | ||||
| 0–10 cm | 4.4 | 0.34 | 4.16 | 23.1 | 0.07 | 38.65 | 3.54 | 3.94 | 7.88 | 0.08 | 0.33 | 0.63 | 0.05 | 0.04 |
| (0.16) | (0.08) | (0.71) | (0.8) | (0.02) | (5.96) | (1.10) | (1.22) | (0.53) | (0.00) | (0.04) | (0.05) | (0.00) | (0.01) | |
| 10–30 cm | 4.55 | 0.28 | 4.22 | 12.4 | 0.06 | 49.22 | 3.1 | 3.45 | 7.2 | 0.1 | 0.41 | 0.62 | 0.05 | 0.03 |
| (0.07) | (0.06) | (0.89) | (1.0) | (0.03) | (10.41) | (1.34) | (1.49) | (1.13) | (0.02) | (0.17) | (0.07) | (0.00) | (0.01) | |
| PD | b | a | a | a | b | ab | b | c | c | a | ||||
| 0–10 cm | 4.18 | 0.46 | 6.86 | 27.2 | 0.11 | 39.45 | 5.29 | 5.92 | 5.82 | 0.09 | 0.21 | 0.44 | 0.05 | 0.12 |
| (0.06) | (0.04) | (0.73) | (1.0) | (0.02) | (3.72) | (0.89) | (0.99) | (1.56) | (0.01) | (0.12) | (0.09) | (0.00) | (0.02) | |
| 10–30 cm | 4.36 | 0.33 | 5.32 | 24.0 | 0.05 | 70.05 | 2.29 | 2.39 | 4.56 | 0.09 | 0.14 | 0.32 | 0.06 | 0.05 |
| (0.05) | (0.04) | (0.79) | (8.5) | (0.01) | (2.55) | (0.42) | (0.48) | (1.68) | (0.02) | (0.07) | (0.12) | (0.01) | (0.01) | |
| Site | Biomass (g m−2) | Necromass (g m−2) | |||||
|---|---|---|---|---|---|---|---|
| <2 mm | 2–5 mm | <5 mm | <2 mm | 2–5 mm | <5 mm | ||
| Fine root mass | |||||||
| ZJ | 246.24 (17.23) a | 193.85 (50.69) a | 440.09 (65.28) ab | 7.18 (1.90) bc | 81.31 (25.17) a | 88.49 (25.29) a | |
| MP | 48.25 (21.28) b | 50.05 (32.2) a | 98.30 (47.48) b | 1.06 (0.57) c | 0 b | 1.06 (0.57) b | |
| QA | 404.87 (56.82) a | 295.43 (117.81) a | 700.30 (153.69) a | 45.46 (9.51) a | 19.04 (14.22) b | 64.49 (8.59) a | |
| PD | 366.77 (55.10) a | 120.44 (30.91) a | 487.21 (57.35) a | 25.62 (7.17) ab | 8.71 (6.01) b | 34.33 (12.62) ab | |
| ANOVA Summary | |||||||
| Variation source | df | <2 mm | 2–5 mm | <5 mm | <2 mm | 2–5 mm | <5 mm |
| Site | 3 | <0.0001 | 0.054 | <0.001 | <0.0001 | 0.001 | 0.001 |
| Soil depth | 5 | <0.0001 | 0.002 | <0.001 | <0.0001 | <0.001 | <0.001 |
| Site x soil depth | 15 | <0.0001 | 0.003 | <0.001 | <0.0001 | <0.001 | <0.001 |
| Site | ZJ | MP | QA | PD |
|---|---|---|---|---|
| Tree density (tree ha−1) | NA | 700 | 900 | 800 |
| Mean height (m) | 3.7 | 13.8 | 12.5 | |
| Mean DBH (cm) | 15.5 | 30.3 | 24.2 | |
| Basal area (m2 ha−1) | 13.4 | 43.7 | 39.4 | |
| Dominant species | Zoysia japonica | Malus pumila | Quercus acutissima | Pinus densiflora |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, L.T.N.; An, J.Y.; Carayugan, M.B.; Hernandez, J.O.; Rahman, S.A.; Youn, W.B.; Carvalho, J.I.; Jo, M.S.; Han, S.H.; Nguyen, H.-H.; et al. Fine-Root Distribution and Soil Physicochemical Property Variations in Four Contrasting Urban Land-Use Types in South Korea. Plants 2024, 13, 164. https://doi.org/10.3390/plants13020164
Tran LTN, An JY, Carayugan MB, Hernandez JO, Rahman SA, Youn WB, Carvalho JI, Jo MS, Han SH, Nguyen H-H, et al. Fine-Root Distribution and Soil Physicochemical Property Variations in Four Contrasting Urban Land-Use Types in South Korea. Plants. 2024; 13(2):164. https://doi.org/10.3390/plants13020164
Chicago/Turabian StyleTran, Lan Thi Ngoc, Ji Young An, Mark Bryan Carayugan, Jonathan O. Hernandez, SK Abidur Rahman, Woo Bin Youn, Julia Inacio Carvalho, Min Seon Jo, Si Ho Han, Hai-Hoa Nguyen, and et al. 2024. "Fine-Root Distribution and Soil Physicochemical Property Variations in Four Contrasting Urban Land-Use Types in South Korea" Plants 13, no. 2: 164. https://doi.org/10.3390/plants13020164
APA StyleTran, L. T. N., An, J. Y., Carayugan, M. B., Hernandez, J. O., Rahman, S. A., Youn, W. B., Carvalho, J. I., Jo, M. S., Han, S. H., Nguyen, H.-H., & Park, B. B. (2024). Fine-Root Distribution and Soil Physicochemical Property Variations in Four Contrasting Urban Land-Use Types in South Korea. Plants, 13(2), 164. https://doi.org/10.3390/plants13020164