
The Identification and Expression Analysis of the Liriodendron chinense F-Box Gene Family

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
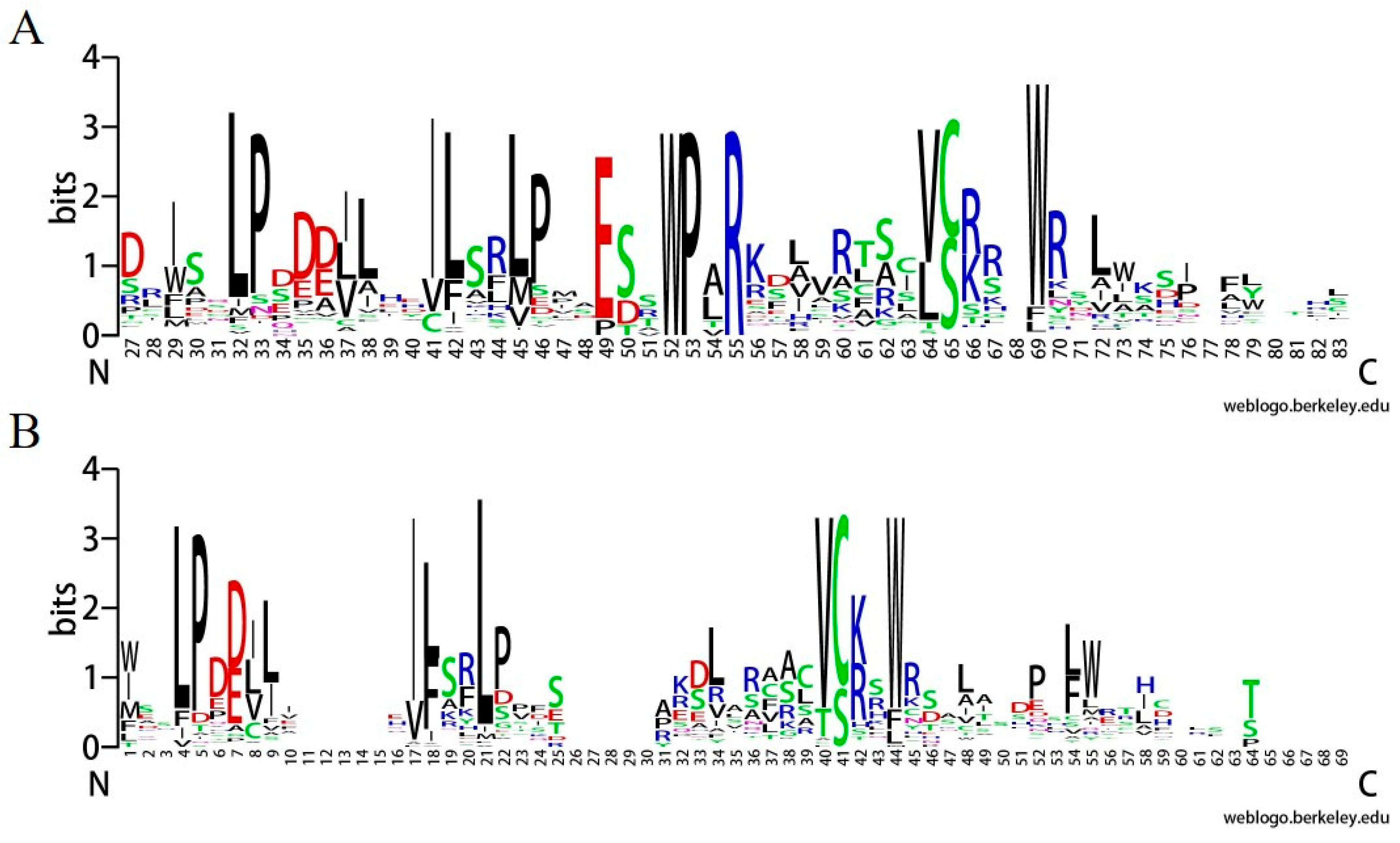
2.1. Genome-Wide Identification, Characteristics, and Classification of F-Box Genes in L. chinense
2.2. Chromosomal Locations, Gene Duplication Events, and Synteny Analysis in the L. chinense F-Box Gene Family
2.3. Subcellular Localization, GO, KEGG Enrichment Analysis, and Protein Interaction Prediction of the F-Box Gene in L. chinense
2.4. F-Box Gene Expression Levels across Different Tissue Types in Liriodendron Hybrids
2.5. F-Box Genes Are Universally Expressed at Different Stages of Somatic Embryogenesis in Liriodendron Hybrids
2.6. F-Box Responds Strongly to Cold, Heat, and Drought Stress Treatments in Liriodendron Hybrids
3. Discussion
4. Materials and Methods
4.1. Database Search for F-Box Proteins in L. chinense
4.2. Sequence Analysis
4.3. Phylogenetic Analysis and Gene Structure Analysis
4.4. Chromosomal Locations, Gene Duplication Analysis, and Synteny Analysis
4.5. Subcellular Localization, GO Enrichment Analysis, and KEGG Enrichment Analysis
4.6. RNA-Seq Analysis of F-Box Gene Expression Levels in Different Organs and Multiple Stresses
4.7. Plant Materials, Treatments, and Collections; RNA Extraction and Quantitative Real-Time PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amm, I.; Sommer, T.; Wolf, D.H. Protein quality control and elimination of protein waste: The role of the ubiquitin-proteasome system. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 15. [Google Scholar] [CrossRef] [PubMed]
- Sadanandom, A.; Bailey, M.; Ewan, R.; Lee, J.; Nelis, S. The ubiquitin-proteasome system: Central modifier of plant signalling. New Phytol. 2012, 196, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.S.; Ou, C.; Chan, Y.R.; Chien, C.T.; Pi, H. The utility F-box for protein destruction. Cell. Mol. Life Sci. 2008, 65, 1977–2000. [Google Scholar] [CrossRef] [PubMed]
- Song, J.B.; Wang, Y.X.; Li, H.B.; Li, B.W.; Zhou, Z.S.; Gao, S.; Yang, Z.M. The F-box family genes as key elements in response to salt, heavy mental, and drought stresses in Medicago truncatula. Funct. Integr. Genom. 2015, 15, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.; Sen, P.; Hofmann, K.; Ma, L.; Goebl, M.; Harper, J.W.; Elledge, S.J. SKP1 connects cell cycle regulators to the ubiquitin proteolysis machinery through a novel motif, the F-box. Cell 1996, 86, 263–274. [Google Scholar] [CrossRef]
- Kipreos, E.T.; Pagano, M. The F-box protein family. Genome Biol. 2000, 1, REVIEWS3002. [Google Scholar] [CrossRef]
- Wang, Z.W.; Liu, P.D.; Inuzuka, H.; Wei, W.Y. Roles of F-box proteins in cancer. Nat. Rev. Cancer 2014, 14, 233–247. [Google Scholar] [CrossRef]
- Xu, G.X.; Ma, H.; Nei, M.; Kong, H.Z. Evolution of F-box genes in plants: Different modes of sequence divergence and their relationships with functional diversification. Proc. Nat. Acad. Sci. USA 2009, 106, 835–840. [Google Scholar] [CrossRef]
- Zhang, S.L.; Tian, Z.L.; Li, H.P.; Guo, Y.T.; Zhang, Y.Q.; Roberts, J.A.; Zhang, X.B.; Miao, Y.C. Genome-wide analysis and characterization of F-box gene family in Gossypium hirsutum L. BMC Genom. 2019, 20, 993. [Google Scholar] [CrossRef]
- Jia, Q.; Xiao, Z.X.; Wong, F.L.; Sun, S.; Liang, K.J.; Lam, H.M. Genome-wide analyses of the soybean F-Box gene family in response to salt stress. Int. J. Mol. Sci. 2017, 18, 818. [Google Scholar] [CrossRef]
- Gupta, S.; Garg, V.; Kant, C.; Bhatia, S. Genome-wide survey and expression analysis of F-box genes in chickpea. BMC Genom. 2015, 16, 67. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.M.; Yin, H.; Qiao, X.; Tan, X.; Gu, C.; Wang, B.H.; Cheng, R.; Wang, Y.Z.; Zhang, S.L. F-box genes: Genome-wide expansion, evolution and their contribution to pollen growth in pear (Pyrus bretschneideri). Plant Sci. 2016, 253, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.T.; Wang, S.Y.; Chen, Y.Y.; Fang, Y.X.; Zhang, X.; Tong, T.; Zhang, Z.L.; Zheng, J.J.; Xue, D.W.; Zhang, X.Q. Genome-wide identification of the F-box gene family and expression analysis under drought and salt stress in Barley. Phyton-Int. J. Exp. Bot. 2020, 89, 229–251. [Google Scholar] [CrossRef]
- Li, Y.; Yapa, M.M.; Hua, Z.H. A machine learning aapproach to prioritizing functionally active F-box members in Arabidopsis thaliana. Front. Plant Sci. 2021, 12, 639253. [Google Scholar] [CrossRef] [PubMed]
- Hua, Z.H. Deciphering the protein ubiquitylation system in plants. J. Exp. Bot. 2023, 74, 6487–6504. [Google Scholar] [CrossRef] [PubMed]
- Hua, Z. Diverse Evolution in 111 Plant Genomes Reveals Purifying and Dosage Balancing Selection Models for F-Box Genes. Int. J. Mol. Sci. 2021, 22, 871. [Google Scholar] [CrossRef] [PubMed]
- Baute, J.; Polyn, S.; De Block, J.; Blomme, J.; Van Lijsebettens, M.; Inze, D. F-box protein FBX92 affects leaf size in Arabidopsis thaliana. Plant Cell Physiol. 2017, 58, 962–975. [Google Scholar] [CrossRef]
- Gray, W.M.; Kepinski, S.; Rouse, D.; Leyser, O.; Estelle, M. Auxin regulates SCF(TIR1)-dependent degradation of AUX/IAA proteins. Nature 2001, 414, 271–276. [Google Scholar] [CrossRef]
- Xu, L.H.; Liu, F.Q.; Lechner, E.; Genschik, P.; Crosby, W.L.; Ma, H.; Peng, W.; Huang, D.F.; Xie, D.X. The SCF(COI1) ubiquitin-ligase complexes are required for jasmonate response in Arabidopsis. Plant Cell 2002, 14, 1919–1935. [Google Scholar] [CrossRef]
- Knauer, S.; Holt, A.L.; Rubio-Somoza, I.; Tucker, E.J.; Hinze, A.; Pisch, M.; Javelle, M.; Timmermans, M.C.; Tucker, M.R.; Laux, T. A protodermal miR394 signal defines a region of stem cell competence in the Arabidopsis shoot meristem. Dev. Cell 2013, 24, 125–132. [Google Scholar] [CrossRef]
- Hong, J.; Lee, J.; Jeong, C.W.; Brooks, J.S.; Choi, Y.; Lee, J.S. Identification of elements responsible for maternally-silenced imprinted gene expression of Upward Curly Leaf 1, an F-box protein gene that regulates curly leaf in Arabidopsis. J. Plant Biol. 2020, 63, 337–346. [Google Scholar] [CrossRef]
- Qu, L.; Lin, L.B.; Xue, H.W. Rice miR394 suppresses leaf inclination through targeting an F-box gene, LEAF INCLINATION 4. J. Integr. Plant Biol. 2019, 61, 406–416. [Google Scholar] [CrossRef]
- Pena-Garcia, Y.; Shinde, S.; Natarajan, P.; Lopez-Ortiz, C.; Balagurusamy, N.; Chavez, A.C.D.; Saminathan, T.; Nimmakayala, P.; Reddy, U.K. Arsenic Stress-Related F-Box (ASRF) gene regulates arsenic stress tolerance in Arabidopsis thaliana. J. Hazard. Mater. 2021, 407, 124831. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.X.; Wang, W.Q.; Wang, W.L.; Zhang, G.Q.; Liu, Y.; Wang, Y.; Wang, W. Wheat F-box protein gene TaFBA1 is involved in plant tolerance to heat stress. Front. Plant Sci. 2018, 9, 521. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.M.; Sun, X.D.; Yin, S.H.; Kong, X.Z.; Zhou, S.; Xu, Y.; Luo, Y.; Wang, W. The role of the F-box gene TaFBA1 from wheat (Triticum aestivum L.) in drought tolerance. Plant Physiol. Bioch. 2014, 84, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Paquis, S.; Mazeyrat-Gourbeyre, F.; Fernandez, O.; Crouzet, J.; Clement, C.; Baillieul, F.; Dorey, S. Characterization of a F-box gene up-regulated by phytohormones and upon biotic and abiotic stresses in grapevine. Mol. Biol. Rep. 2011, 38, 3327–3337. [Google Scholar] [CrossRef] [PubMed]
- Li, M.P.; Wang, D.; Long, X.F.; Hao, Z.D.; Lu, Y.; Zhou, Y.W.; Peng, Y.; Cheng, T.L.; Shi, J.S.; Chen, J.H. Agrobacterium-mediated genetic transformation of embryogenic callus in a Liriodendron Hybrid (L. chinense × L. tulipifera). Front. Plant Sci. 2022, 13, 802128. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.D.; Liu, S.Q.; Hu, L.F.; Shi, J.S.; Chen, J.H. Transcriptome analysis and metabolic profiling reveal the key role of carotenoids in the petal coloration of Liriodendron tulipifera. Hort. Res. 2020, 7, 1955–1970. [Google Scholar] [CrossRef]
- Chen, J.H.; Hao, Z.D.; Guang, X.M.; Zhao, C.X.; Wang, P.K.; Xue, L.J.; Zhu, Q.H.; Yang, L.F.; Sheng, Y.; Zhou, Y.W.; et al. Liriodendron genome sheds light on angiosperm phylogeny and species-pair differentiation. Nat. Plants 2019, 5, 18–25. [Google Scholar] [CrossRef]
- Guan, Y.L.; Liu, S.Q.; Wu, W.H.; Hong, K.Y.; Li, R.X.; Zhu, L.M.; Liu, Y.; Lu, Y.; Chen, J.H.; Yang, L.M.; et al. Genome-wide identification and cold stress-induced expression analysis of the CBF gene family in Liriodendron chinense. J. For. Res. 2021, 32, 2531–2543. [Google Scholar] [CrossRef]
- Wu, W.H.; Zhu, S.; Xu, L.; Zhu, L.M.; Wang, D.D.; Liu, Y.; Liu, S.Q.; Hao, Z.D.; Lu, Y.; Yang, L.M.; et al. Genome-wide identification of the Liriodendron chinense WRKY gene family and its diverse roles in response to multiple abiotic stress. BMC Plant Biol. 2022, 22, 25. [Google Scholar] [CrossRef]
- Wu, W.H.; Zhu, S.; Zhu, L.M.; Wang, D.D.; Liu, Y.; Liu, S.Q.; Zhang, J.J.; Hao, Z.D.; Lu, Y.; Cheng, T.L.; et al. Characterization of the Liriodendron chinense MYB gene family and its role in abiotic stress response. Front. Plant Sci. 2021, 12, 641280. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.J.; Ma, Q.X.; Weng, Y.H.; Shi, J.S.; Chen, J.H.; Hao, Z.D. Genome-wide identification and expression analysis of TPS gene family in Liriodendron chinense. Genes 2023, 14, 770. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.F.; Wang, P.K.; Long, X.F.; Wu, W.H.; Zhang, J.J.; Pan, Y.; Cheng, T.L.; Shi, J.S.; Chen, J.H. The PIN gene family in relicplant L. chinense: Genome-wide identification and gene expression profiling in different organizations and abiotic stress responses. Plant Physiol. Bioch. 2021, 162, 634–646. [Google Scholar] [CrossRef]
- Hua, Z.; Zou, C.; Shiu, S.H.; Vierstra, R.D. Phylogenetic comparison of F-Box (FBX) gene superfamily within the plant kingdom reveals divergent evolutionary histories indicative of genomic drift. PLoS ONE 2011, 6, e16219. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Y.; Kalluri, U.C.; Jawdy, S.; Gunter, L.E.; Yin, T.M.; Tschaplinski, T.J.; Weston, D.J.; Ranjan, P.; Tuskan, G.A. The F-box gene family is expanded in herbaceous annual plants relative to woody perennial plants. Plant Physiol. 2008, 148, 1189–1200. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef]
- Schultz, T.F.; Kiyosue, T.; Yanovsky, M.; Wada, M.; Kay, S.A. A role for LKP2 in the circadian clock of Arabidopsis. Plant Cell 2001, 13, 2659–2670. [Google Scholar] [CrossRef]
- Gomez-Gomez, L.; Boller, T. FLS2: An LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Mol. Cell 2000, 5, 1003–1011. [Google Scholar] [CrossRef]
- Buscaill, P.; Chandrasekar, B.; Sanguankiattichai, N.; Kourelis, J.; Kaschani, F.; Thomas, E.L.; Morimoto, K.; Kaiser, M.; Preston, G.M.; Ichinose, Y.; et al. Glycosidase and glycan polymorphism control hydrolytic release of immunogenic flagellin peptides. Science 2019, 364, 145. [Google Scholar] [CrossRef]
- Yamazaki, M.; Saito, K. Molecular genetic study on the anthocyanin chemotypes of Perilla frutescens var. crispa. Nat. Prod. Commun. 2011, 6, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Carey, C.C.; Strahle, J.T.; Selinger, D.A.; Chandler, V.L. Mutations in the pale aleurone color1 regulatory gene of the Zea mays anthocyanin pathway have distinct phenotypes relative to the functionally similar TRANSPARENT TESTA GLABRA1 gene in Arabidopsis thaliana. Plant Cell 2004, 16, 450–464. [Google Scholar] [CrossRef]
- Matus, J.T.; Poupin, M.J.; Canon, P.; Bordeu, E.; Alcalde, J.A.; Arce-Johnson, P. Isolation of WDR and bHLH genes related to flavonoid synthesis in grapevine (Vitis vinifera L.). Plant Mol. Biol. 2010, 72, 607–620. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.R.; Davison, P.A.; Bolognesi-Winfield, A.C.; James, C.M.; Srinivasan, N.; Blundell, T.L.; Esch, J.J.; Marks, M.D.; Gray, J.C. The TRANSPARENT TESTA GLABRA1 locus, which regulates trichome differentiation and anthocyanin biosynthesis in Arabidopsis, encodes a WD40 repeat protein. Plant Cell 1999, 11, 1337–1350. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.E.; Advani, V.R. Chloroplast and cytoplasmic enzymes: Three distinct isoenzymes associated with the reductive pentose phosphate cycle. Plant Physiol. 1970, 45, 583–585. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.H.; Yoo, S.D. Signaling role of fructose mediated by FINS1/FBP in Arabidopsis thaliana. PLoS Genet. 2011, 7, e1001263. [Google Scholar] [CrossRef]
- Guo, X.Y.; Zhang, L.; Dong, G.Q.; Xu, Z.H.; Li, G.M.; Liu, N.; Wang, A.Y.; Zhu, J.B. A novel cold-regulated protein isolated from Saussurea involucrata confers cold and drought tolerance in transgenic tobacco (Nicotiana tabacum). Plant Sci. 2019, 289, 110246. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef]
- Lu, S.N.; Wang, J.Y.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene_Id | Go Number | Arabidopsis Ortholog Genes | Putative Function of Arabidopsis Orthologs |
|---|---|---|---|
| Lchi01743 | 3 | AT5G39450 | biological process |
| Lchi01886 | 4 | AT1G21890 | enables transmembrane transporter activity |
| Lchi03271 | 2 | AT5G57360 | flower development and protein ubiquitination |
| Lchi14963 | 6 | AT5G43190 | ubiquitin-dependent protein catabolic process |
| Lchi15076 | 6 | AT5G43190 | ubiquitin-dependent protein catabolic process |
| Lchi17591 | 3 | AT3G54460 | enables ATP-dependent chromatin remodeling activity |
| Lchi23560 | 2 | AT2G17020 | ubiquitin-dependent protein catabolic process |
| Lchi26033 | 2 | AT1G12870 | regulation of transcription |
| Lchi31059 | 2 | AT1G68050 | positive regulation of flower development |
| Lchi31193 | 4 | AT4G08850 | hormone-mediated signaling pathway |
| Lchi33849 | 2 | AT1G13200 | regulation of transcription |
| Lchi34026 | 2 | AT1G23770 | biological process |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, S.; Xu, L.; Wu, W.; Zhang, J.; Hao, Z.; Lu, L.; Shi, J.; Chen, J. The Identification and Expression Analysis of the Liriodendron chinense F-Box Gene Family. Plants 2024, 13, 171. https://doi.org/10.3390/plants13020171
He S, Xu L, Wu W, Zhang J, Hao Z, Lu L, Shi J, Chen J. The Identification and Expression Analysis of the Liriodendron chinense F-Box Gene Family. Plants. 2024; 13(2):171. https://doi.org/10.3390/plants13020171
Chicago/Turabian StyleHe, Shichan, Lin Xu, Weihuang Wu, Jiaji Zhang, Zhaodong Hao, Lu Lu, Jisen Shi, and Jinhui Chen. 2024. "The Identification and Expression Analysis of the Liriodendron chinense F-Box Gene Family" Plants 13, no. 2: 171. https://doi.org/10.3390/plants13020171