
Meta-Analysis of the Effects of Overexpressed bZIP Transcription Factors in Plants under Drought Stress



Abstract
1. Introduction
2. Results
2.1. Summary of Overall Effects
2.2. Heterogeneity Analysis
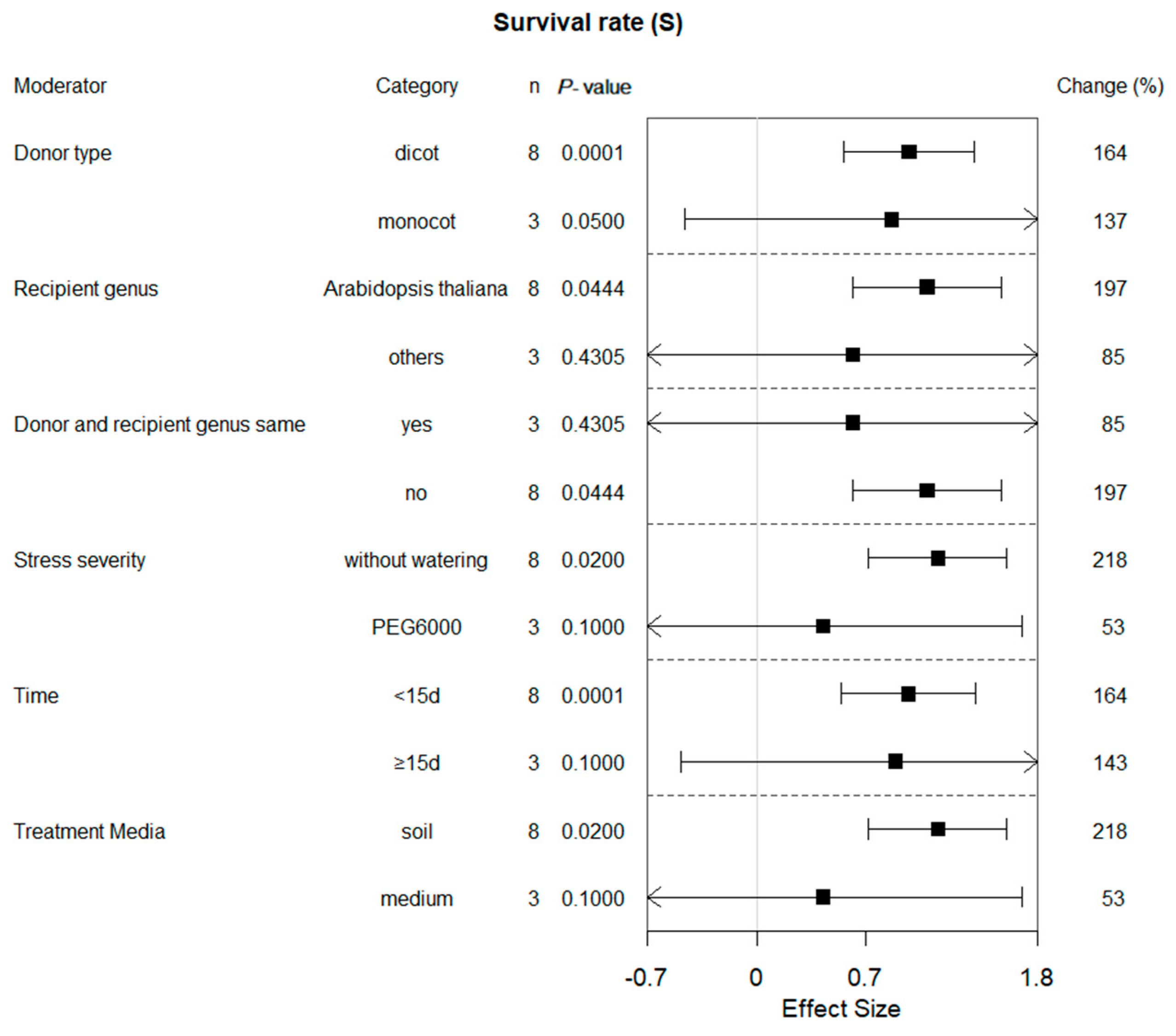
2.3. Subgroup Effect within Each Moderator on Survival Rate in Plants under Drought Conditions
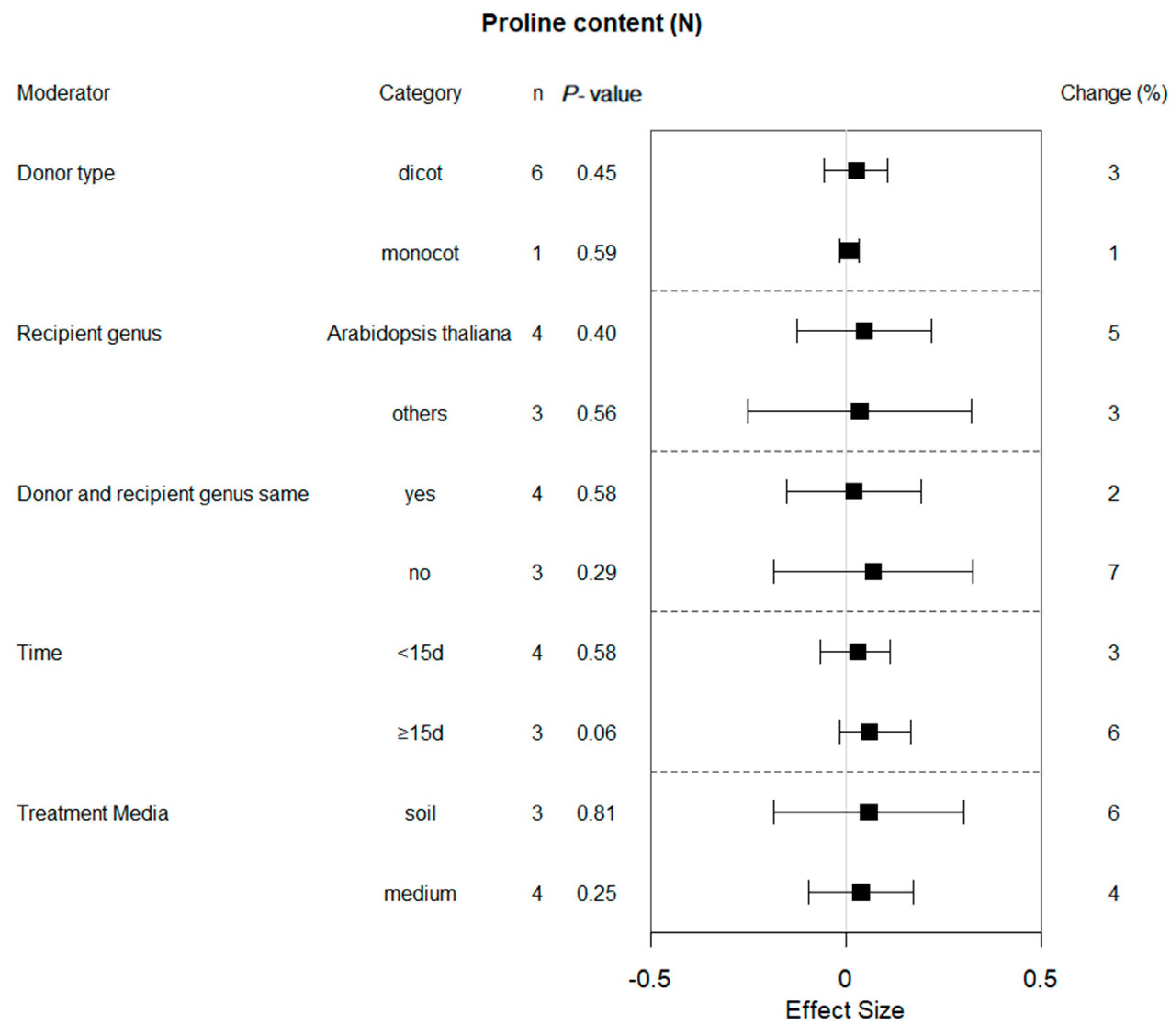
2.4. Subgroup Effect within Each Moderator on Proline Content in Plants That Experienced Drought and Non-Stressed Conditions
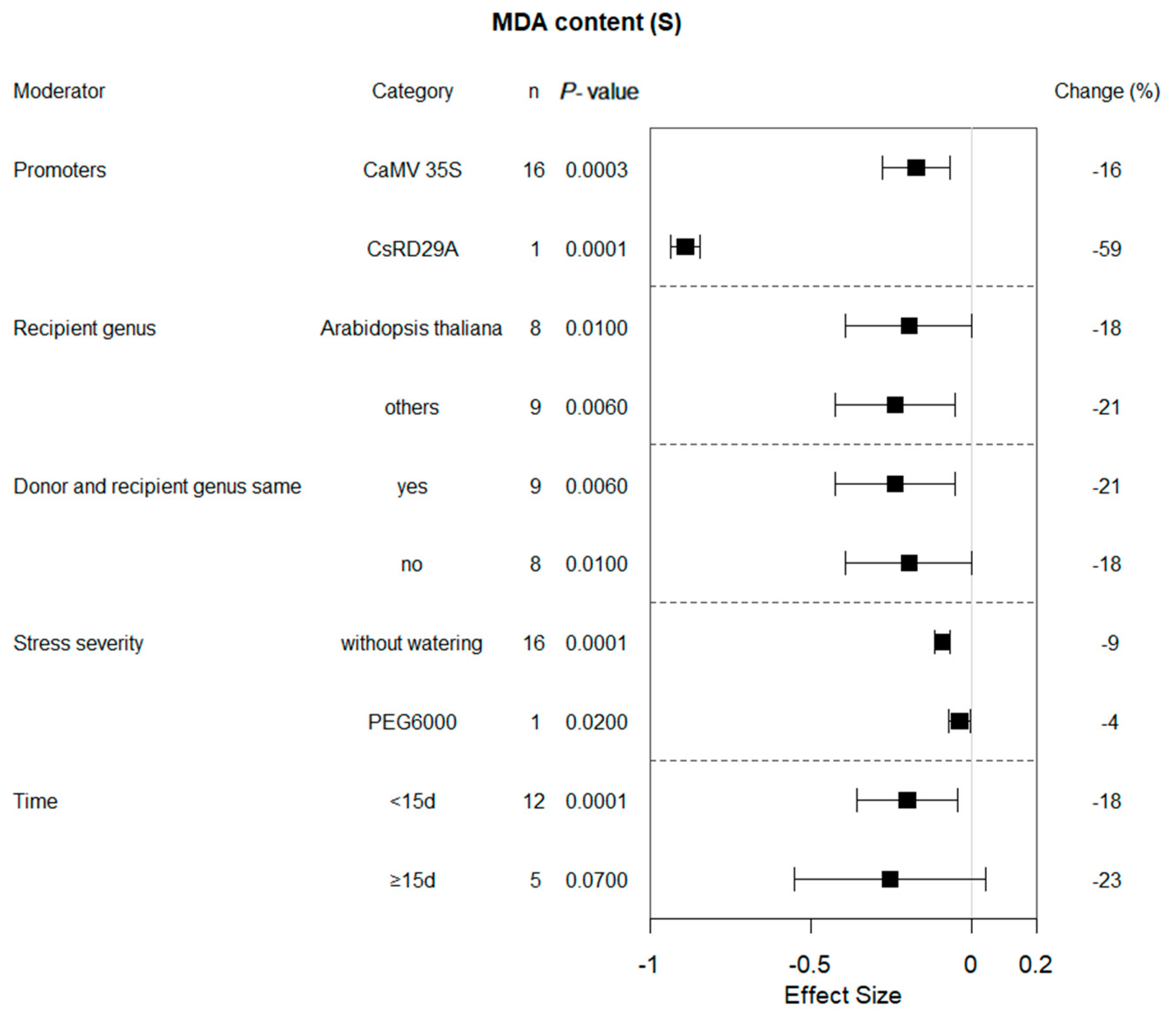
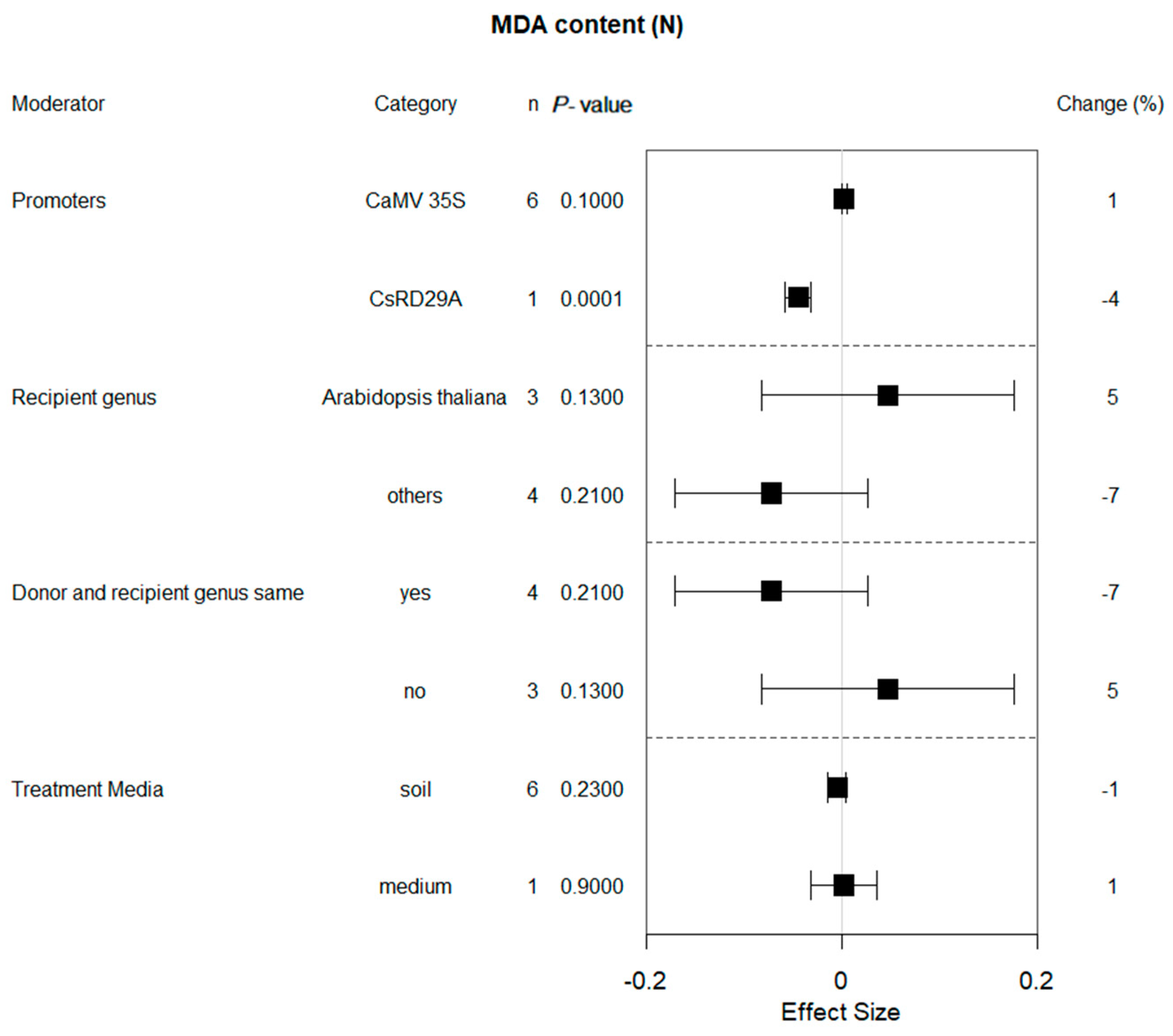
2.5. Moderator Analysis of MDA Content in Plants Subjected to Drought and Non-Stressed Conditions
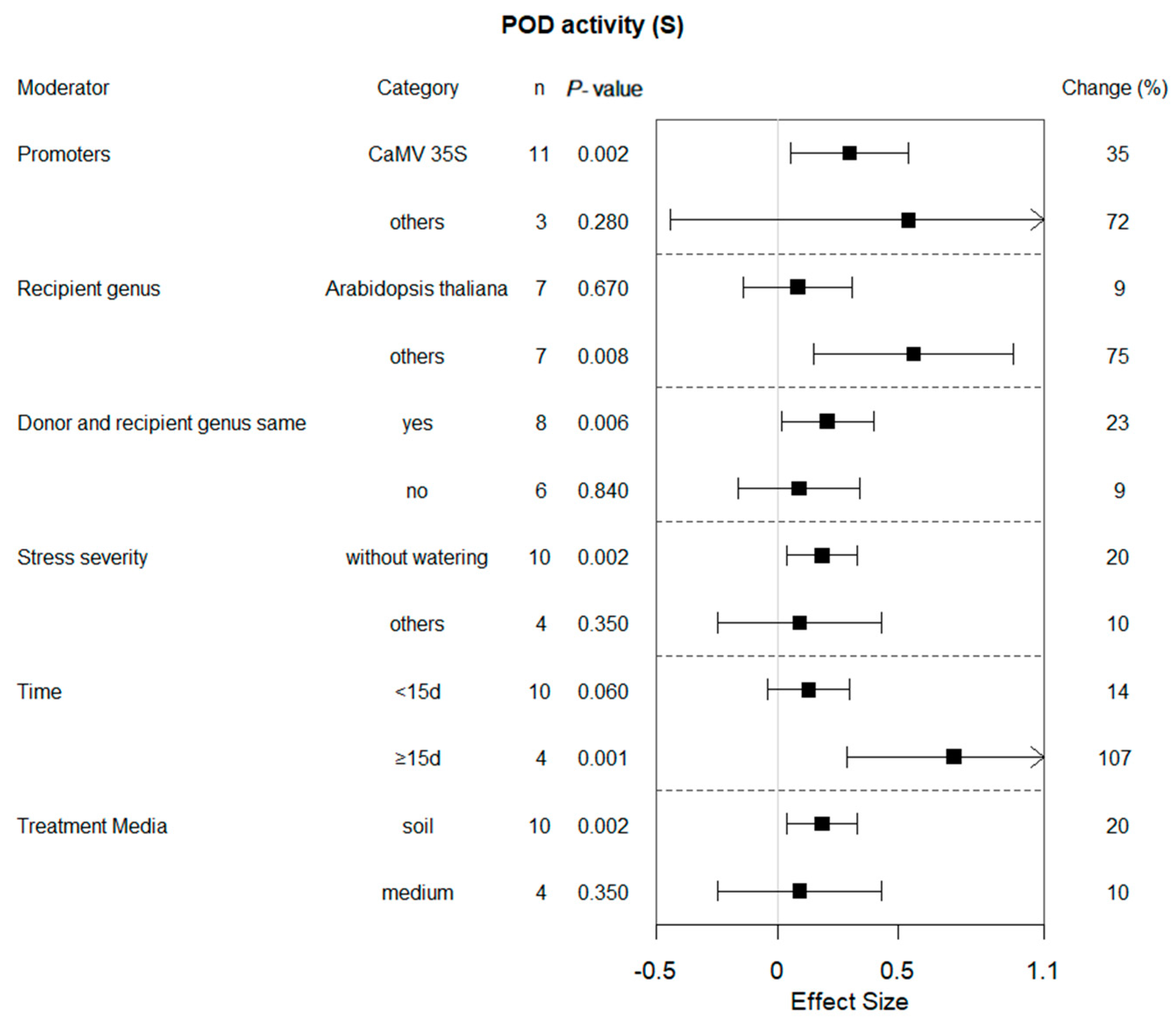
2.6. Subgroup Analysis of POD Activity in Plants under Drought Conditions
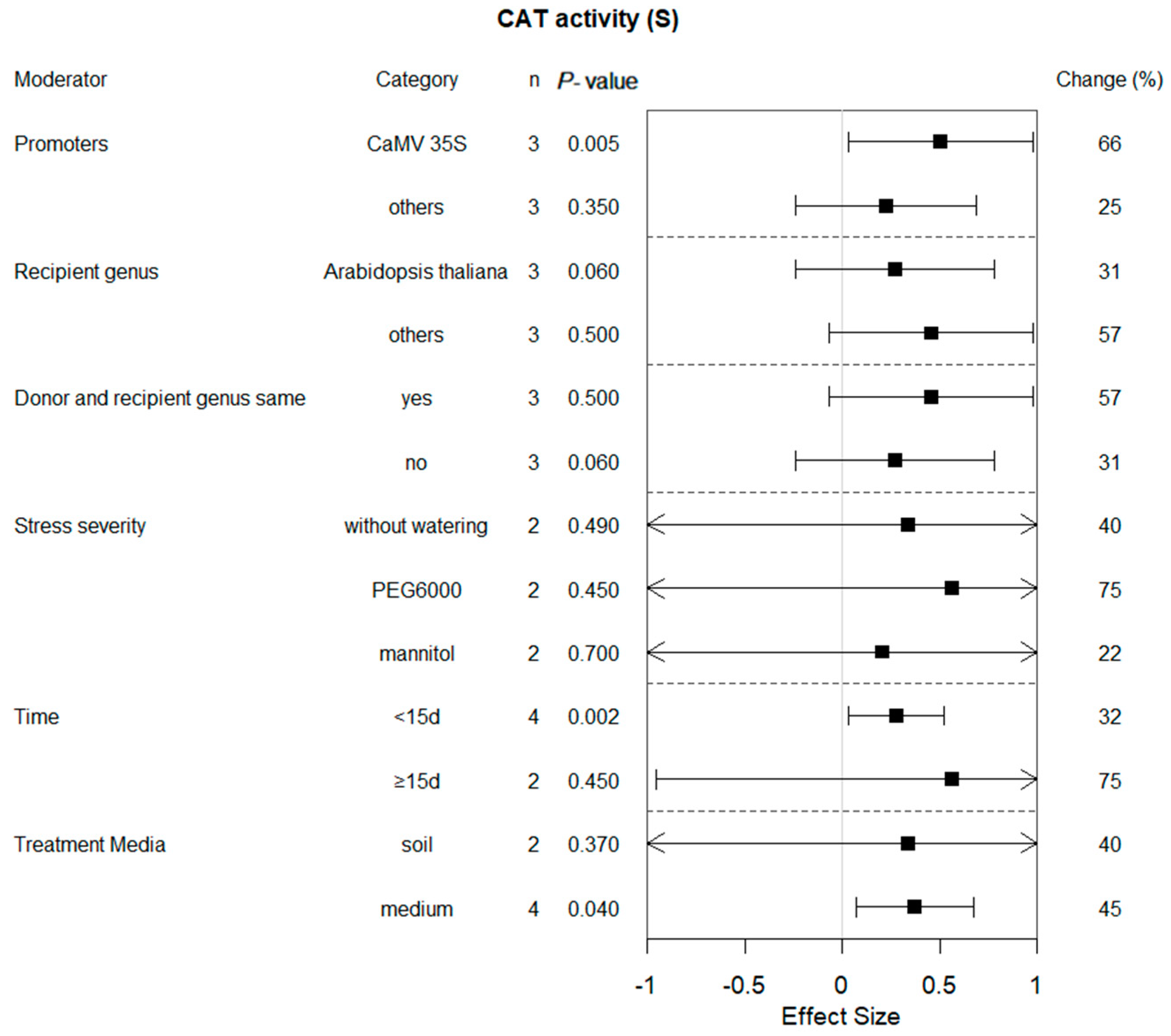
2.7. Subgroup Analysis of CAT Activity in Plants under Drought Conditions
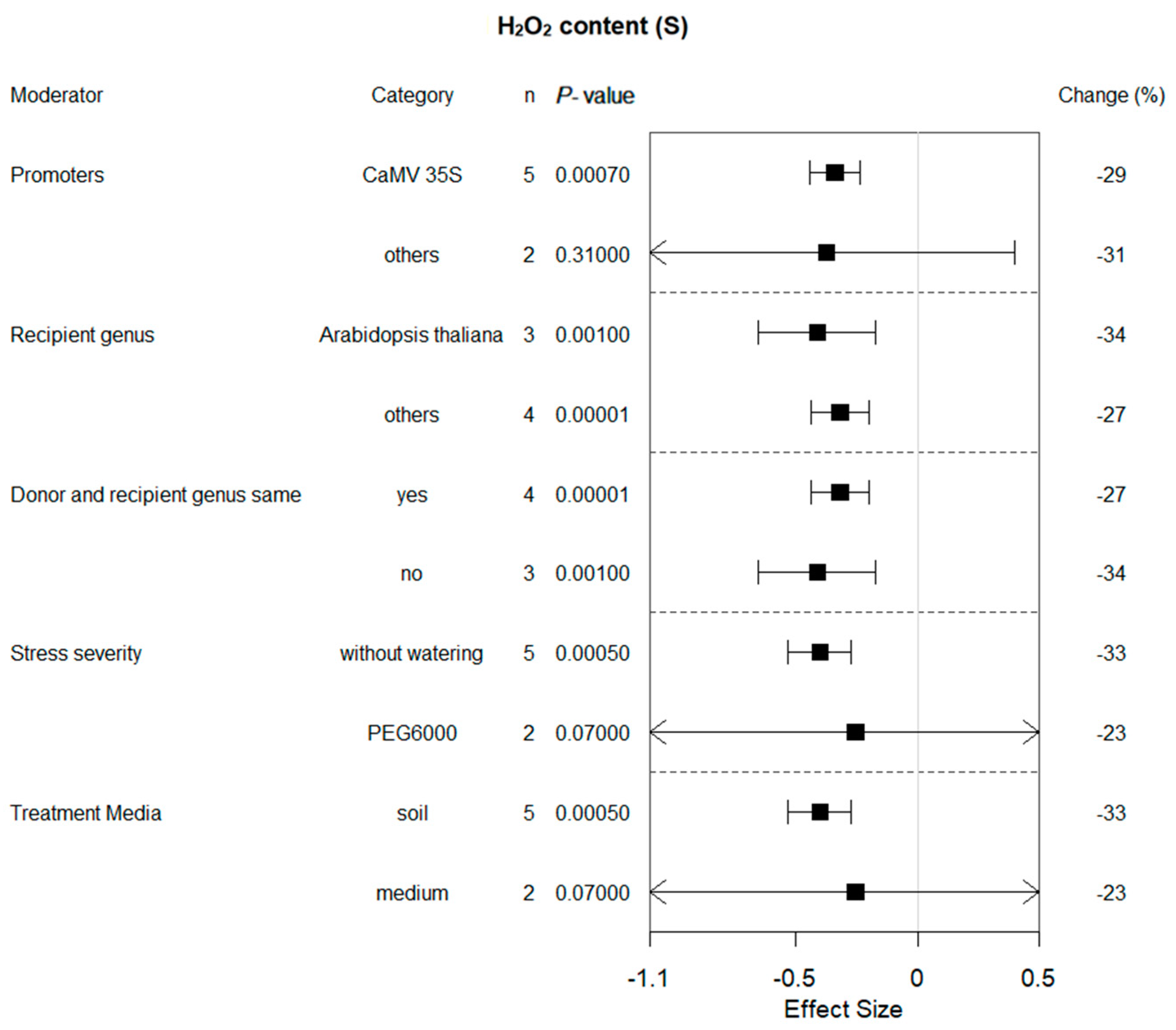
2.8. Subgroup Analysis of H2O2 Content in Plants under Drought Conditions
3. Discussion
4. Materials and Methods
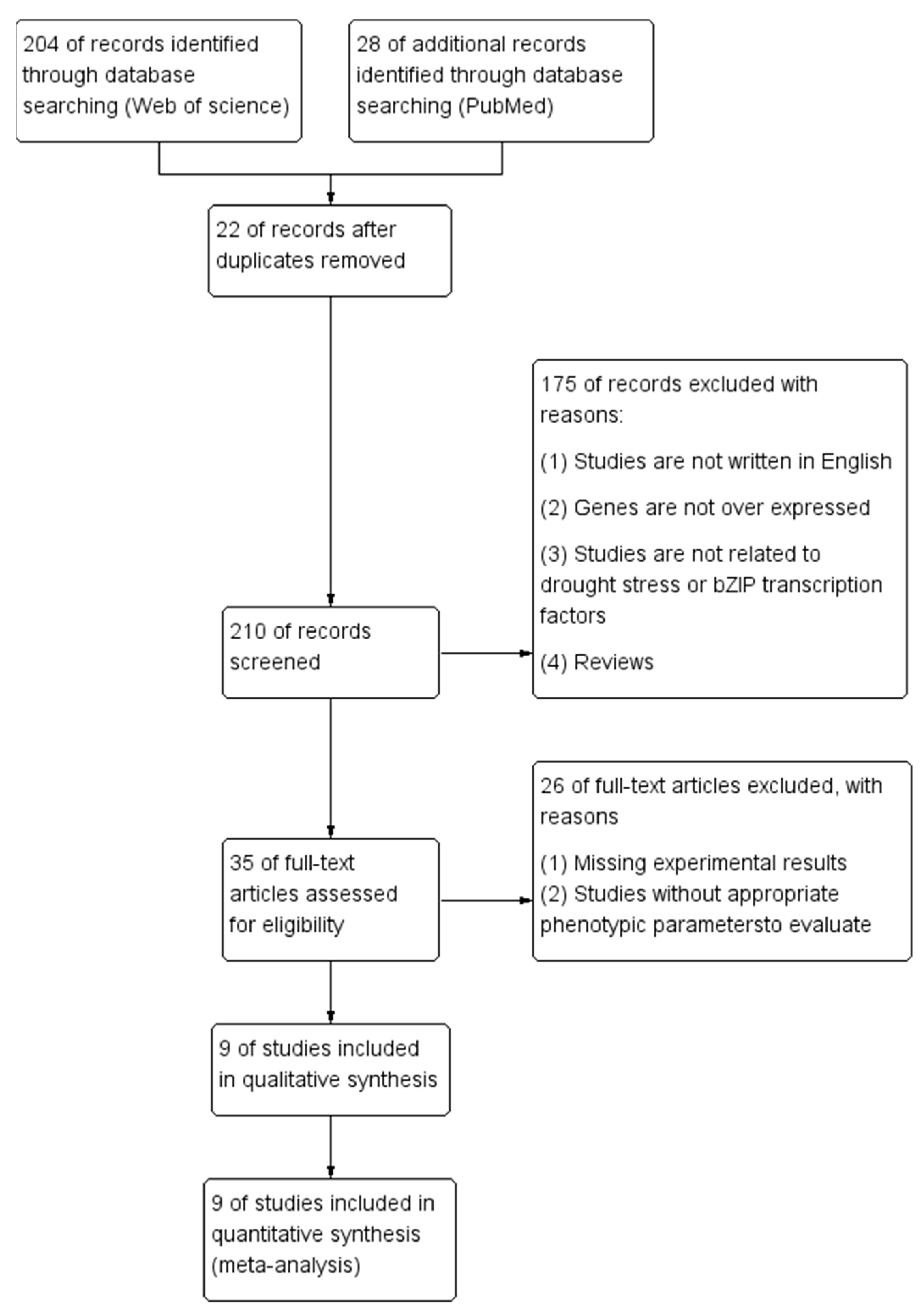
4.1. Data Collection
- It should be written in English;
- It should be published between 2013 and 2023;
- It should quantitatively assess the effect of drought on at least some outcome parameters of interest (such as proline content and survival rate);
- Mutant plants or co-expressed with other genes are not considered;
- It is not a review or meeting report.
4.2. Effect Size and Moderator
4.3. Meta-Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, J.; Wang, J.; Hui, W.; Zhao, F.; Wang, P.; Su, C.; Gong, W. Physiology of Plant Responses to Water Stress and Related Genes: A Review. Forests 2022, 13, 324. [Google Scholar] [CrossRef]
- Erpen, L.; Devi, H.; Grosser, J.W.; Dutt, M. Potential use of the DREB/ERF, MYB, NAC and WRKY transcription factors to improve abiotic and biotic stress in transgenic plants. Plant Cell Tissue Organ Cult. 2018, 132, 1–25. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, Z.-C.; Ji, A.-J.; Song, J.-Y. Regulation of secondary metabolite biosynthesis by bZIP transcription factors in plants. Plant Sci. J. 2017, 35, 128–137. [Google Scholar]
- Huang, X.-S.; Liu, J.-H.; Chen, X.-J. Overexpression of PtrABF gene, a bZIP transcription factor isolated from Poncirus trifoliata, enhances dehydration and drought tolerance in tobacco via scavenging ROS and modulating expression of stress-responsive genes. BMC Plant Biol. 2010, 10, 230. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Tang, X.; Zhang, N.; Li, S.; Si, H. Role of bZIP Transcription Factors in Plant Salt Stress. Int. J. Mol. Sci. 2023, 24, 7893. [Google Scholar] [CrossRef]
- Borenstein, M.; Hedges, L.V.; Higgins, J.P.T.; Rothstein, H.R. Cumulative Meta-Analysis. Introduction to Meta Analysis. Int. Coach. Psychol. Rev. 2009, 15, 121–122. [Google Scholar] [CrossRef]
- Kong, W.; Zhong, H.; Gong, Z.; Fang, X.; Sun, T.; Deng, X.; Li, Y. Meta-Analysis of Salt Stress Transcriptome Responses in Different Rice Genotypes at the Seedling Stage. Plants 2019, 8, 64. [Google Scholar] [CrossRef]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front. Plant Sci. 2014, 5, 151. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, T.F.; Ma, J.; Chen, J.; Zhou, Y.-B.; Chen, M.; Ma, Y.-Z.; Wei, W.-L.; Xu, Z.-S. The Soybean bZIP Transcription Factor Gene GmbZIP2 Confers Drought and Salt Resistances in Transgenic Plants. Int. J. Mol. Sci. 2020, 21, 670. [Google Scholar] [CrossRef]
- Yang, S.; Xu, K.; Chen, S.; Li, T.; Xia, H.; Chen, L.; Liu, H.; Luo, L. A stress-responsive bZIP transcription factor OsbZIP62 improves drought and oxidative tolerance in rice. BMC Plant Biol. 2019, 19, 260. [Google Scholar] [CrossRef]
- Li, Q.; Wu, Q.; Wang, A.; Lv, B.; Dong, Q.; Yao, Y.; Wu, Q.; Zhao, H.; Li, C.; Chen, H. Tartary buckwheat transcription factor FtbZIP83 improves the drought/salt tolerance of Arabidopsis via an ABA-mediated pathway. Plant Physiol. Biochem. 2019, 144, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Feng, B.E.; Zhang, F.J.; Tang, Y.M.; Zhang, L.; Ma, L.; Zhao, C.; Gao, S. Bioinformatic Analyses of Subgroup-A Members of the Wheat bZIP Transcription Factor Family and Functional Identification of TabZIP174 Involved in Drought Stress Response. Front. Plant Sci. 2016, 7, 1643. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, L.; Xia, C.; Zhao, G.; Liu, J.; Jia, J.; Kong, X. A novel wheat bZIP transcription factor, TabZIP60, confers multiple abiotic stress tolerances in transgenic Arabidopsis. Physiol. Plant. 2015, 153, 538–554. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Liu, X.; Yao, Y.; Li, Y.H.; Liu, S.; He, C.Y.; Li, J.M.; Lin, Y.Y.; Li, L. Overexpression of Arachis hypogaea AREB1 Gene Enhances Drought Tolerance by Modulating ROS Scavenging and Maintaining Endogenous ABA Content. Int. J. Mol. Sci. 2013, 14, 12827–12842. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.Y.; Hu, Y.F.; Li, J.J.; Yao, X.; Liu, K.D. BnaABF2, a bZIP transcription factor from rapeseed (Brassica napus L.), enhances drought and salt tolerance in transgenic Arabidopsis. Bot. Stud. 2016, 57, 12. [Google Scholar] [CrossRef]
- Xu, D.B.; Gao, S.Q.; Ma, Y.Z.; Xu, Z.S.; Zhao, C.P.; Tang, Y.M.; Li, X.Y.; Li, L.C.; Chen, Y.F.; Chen, M.J.F.; et al. ABI-like transcription factor gene TaABL1 from wheat improves multiple abiotic stress tolerances in transgenic plants. Funct. Integr. Genom. 2014, 14, 717–730. [Google Scholar] [CrossRef]
- Amack, S.C.; Antunes, M.S. CaMV35S promoter—A plant biology and biotechnology workhorse in the era of synthetic biology. Curr. Plant Biol. 2020, 24, 100179. [Google Scholar] [CrossRef]
- de Dios Alché, J. A concise appraisal of lipid oxidation and lipoxidation in higher plants. Redox Biol. 2019, 23, 101136. [Google Scholar]
- Li, Y.; Xian, X.; Guo, L.; Zhang, J.; Gan, C.; Wang, Z.; Li, H.; Li, X.; Yuan, X.; Zhang, N.; et al. CsbZIP50 binds to the G-box/ABRE motif in CsRD29A promoter to enhance drought tolerance in cucumber. Environ. Exp. Bot. 2022, 199, 104884. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Liu, C.; Yin, H.; Liu, H.; Luo, H.; He, M.; Zhou, Y. CgbZIP1: A bZIP Transcription Factor from Chrysanthemum Grandiflora Confers Plant Tolerance to Salinity and Drought Stress. Agronomy 2022, 12, 556. [Google Scholar] [CrossRef]
- Zhou, M.; Cheng, H.; Chiang, V.L.; Li, W.; Yang, C.; Wang, C. PtrbZIP3 transcription factor regulates drought tolerance of Populus trichocarpa. Environ. Exp. Bot. 2023, 208, 105231. [Google Scholar] [CrossRef]
- Wang, B.; Li, L.; Liu, M.; Peng, D.; Wei, A.; Hou, B.; Lei, Y.; Li, X. TaFDL2-1A confers drought stress tolerance by promoting ABA biosynthesis, ABA responses, and ROS scavenging in transgenic wheat. Plant J. 2022, 112, 722–737. [Google Scholar] [CrossRef]
- Joo, J.; Lee, Y.H.; Song, S.I. Overexpression of the rice basic leucine zipper transcription factor OsbZIP12 confers drought tolerance to rice and makes seedlings hypersensitive to ABA. Plant Biotechnol. Rep. 2014, 8, 431–441. [Google Scholar] [CrossRef]
- Li, Z.; Li, S.; Wu, W.; Shao, M.; Zhang, X. Grain-filling characters of different genotype winter wheat under nitrogen fertilization in semi-humid area of south Loess Plateau. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2006, 17, 75–79. [Google Scholar]
- Serraj, R.; Sinclair, T.R. Osmolyte accumulation: Can it really help increase crop yield under drought conditions? Plant Cell Environ. 2002, 25, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Slama, I.; Messedi, D.; Ghnaya, T.; Savoure, A.; Abdelly, C. Effects of water deficit on growth and proline metabolism in Sesuvium portulacastrum. Environ. Exp. Bot. 2006, 56, 231–238. [Google Scholar] [CrossRef]
- Kang, C.; Zhai, H.; He, S.; Zhao, N.; Liu, Q. A novel sweetpotato bZIP transcription factor gene, IbbZIP1, is involved in salt and drought tolerance in transgenic Arabidopsis. Plant Cell Rep. 2019, 38, 1373–1382. [Google Scholar] [CrossRef]
- Zhu, M.; Meng, X.; Cai, J.; Li, G.; Dong, T.; Li, Z. Basic leucine zipper transcription factor SlbZIP1 mediates salt and drought stress tolerance in tomato. BMC Plant Biol. 2018, 18, 83. [Google Scholar] [CrossRef]
- Bekka, S.; Abrous-Belbachir, O.; Slovenica, R. Effects of exogenous proline on the physiological characteristics of Triticum aestivum L. and Lens culinaris Medik. under drought stress. Acta Agric. Slov. 2018, 111, 477–491. [Google Scholar] [CrossRef]
- Schaeffer, R.N.; Manson, J.S.; Irwin, R.E. Effects of abiotic factors and species interactions on estimates of male plant function: A meta-analysis. Ecol. Lett. 2013, 16, 399–408. [Google Scholar] [CrossRef]
- Pan, Y.; Hu, X.; Li, C.; Xu, X.; Su, C.; Li, J.; Song, H.; Zhang, X.; Pan, Y. SlbZIP38, a Tomato bZIP Family Gene Downregulated by Abscisic Acid, Is a Negative Regulator of Drought and Salt Stress Tolerance. Genes 2017, 8, 402. [Google Scholar] [CrossRef]
- Zhong, L.; Chen, D.; Min, D.; Li, W.; Xu, Z.; Zhou, Y.; Li, L.; Chen, M.; Ma, Y. AtTGA4, a bZIP transcription factor, confers drought resistance by enhancing nitrate transport and assimilation in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2015, 457, 433–439. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, Y.; Cai, H.; Guo, M.; Qin, Y. The bZIP Transcription Factor GmbZIP15 Negatively Regulates Salt- and Drought-Stress Responses in Soybean. Int. J. Mol. Sci. 2020, 21, 7778. [Google Scholar] [CrossRef]
- Kurepa, J.; Smalle, J.; Va, M.; Montagu, N.; Inzé, D. Oxidative stress tolerance and longevity in Arabidopsis: The late-flowering mutant gigantea is tolerant to paraquat. Plant J. 2010, 14, 759–764. [Google Scholar] [CrossRef]
- Liang, C.; Meng, Z.; Meng, Z.; Malik, W.; Yan, R.; Lwin, K.M.; Lin, F.; Wang, Y.; Sun, G.; Zhou, T.; et al. GhABF2, a bZIP transcription factor, confers drought and salinity tolerance in cotton (Gossypium hirsutum L.). Sci. Rep. 2016, 6, 35040. [Google Scholar] [CrossRef]
- Zhang, H.; Mao, L.; Xin, M.; Xing, H.; Zhang, Y.; Wu, J.; Xu, D.; Wang, Y.; Shang, Y.; Wei, L.; et al. Overexpression of GhABF3 increases cotton(Gossypium hirsutum L.) tolerance to salt and drought. BMC Plant Biol. 2022, 22, 313. [Google Scholar] [CrossRef]
- Wang, W.; Qiu, X.; Yang, Y.; Kim, H.S.; Kwak, S.S. Sweetpotato bZIP Transcription Factor IbABF4 Confers Tolerance to Multiple Abiotic Stresses. Front. Plant Sci. 2019, 10, 630. [Google Scholar] [CrossRef]
- Klümper, W.; Qaim, M. A meta-analysis of the impacts of genetically modified crops. PLoS ONE 2014, 9, e111629. [Google Scholar] [CrossRef]
- Ma, Y.; Robert, A.; Dong, C.; Cheng, M. Increased salt tolerance with overexpression of cation/proton antiporter 1 genes: A meta-analysis. Plant Biotechnol. J. 2016, 15, 162–173. [Google Scholar] [CrossRef]
- Kivlin, S.N.; Emery, S.M.; Rudgers, J.A. Fungal symbionts alter plant responses to global change. Am. J. Bot. 2013, 100, 1445–1457. [Google Scholar] [CrossRef]
- Lehmann, A.; Barto, E.K.; Powell, J.R.; Rillig, M.C. Mycorrhizal responsiveness trends in annual crop plants and their wild relatives—A meta-analysis on studies from 1981 to 2010. Plant Soil 2012, 355, 231–250. [Google Scholar] [CrossRef]
- Higgins, J.P.T.; Thompson, S.G. Quantifying heterogeneity in a meta-analysis. Stat. Med. 2002, 21, 1539–1558. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Qt | p | I2 (%) |
|---|---|---|---|
| Survival | 18.714 | 0.044 | 46.56 |
| Root length | 4.3180 | 0.5046 | 0 |
| Electrolyte leakage | 1 | 0.31731 | 0.00 |
| Proline content | 19.7831 | 0.04841 | 44.40 |
| Chlorophyll content | 9.8793 | 0.2736 | 19.02 |
| H2O2 content | 12.6098 | 0.04967 | 52.42 |
| CAT activity | 11.0961 | 0.04951 | 54.94 |
| POD activity | 13.6856 | 0.39635 | 5.01 |
| SOD activity | 7.9343 | 0.89272 | 0 |
| MDA content | 26.4189 | 0.04841 | 39.44 |
| Trait | Qt | p | I2 (%) |
|---|---|---|---|
| Survival | |||
| Root length | 3.244 | 0.3555 | 7.52 |
| Electrolyte leakage | 0.1628 | 0.68656 | 0 |
| Proline content | 4.6492 | 0.58952 | 0 |
| Chlorophyll content | 4.9435 | 0.42282 | 0 |
| H2O2 content | 3.7056 | 0.59253 | 0 |
| CAT activity | 1.3531 | 0.50838 | 0 |
| POD activity | 1.5722 | 0.66571 | 0 |
| SOD activity | 7.3340 | 0.19696 | 31.82 |
| MDA content | 3.6885 | 0.71874 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, R.; Liu, Y.; Chen, S.; Shityakov, S. Meta-Analysis of the Effects of Overexpressed bZIP Transcription Factors in Plants under Drought Stress. Plants 2024, 13, 337. https://doi.org/10.3390/plants13030337
Tao R, Liu Y, Chen S, Shityakov S. Meta-Analysis of the Effects of Overexpressed bZIP Transcription Factors in Plants under Drought Stress. Plants. 2024; 13(3):337. https://doi.org/10.3390/plants13030337
Chicago/Turabian StyleTao, Ran, Yaqiu Liu, Su Chen, and Sergey Shityakov. 2024. "Meta-Analysis of the Effects of Overexpressed bZIP Transcription Factors in Plants under Drought Stress" Plants 13, no. 3: 337. https://doi.org/10.3390/plants13030337
APA StyleTao, R., Liu, Y., Chen, S., & Shityakov, S. (2024). Meta-Analysis of the Effects of Overexpressed bZIP Transcription Factors in Plants under Drought Stress. Plants, 13(3), 337. https://doi.org/10.3390/plants13030337