
Integrating Smart Greenhouse Cover, Reduced Nitrogen Dose and Biostimulant Application as a Strategy for Sustainable Cultivation of Cherry Tomato
 , , ,
, , ,  , , ,
, , , 
Abstract
1. Introduction
2. Results
2.1. Air Temperature
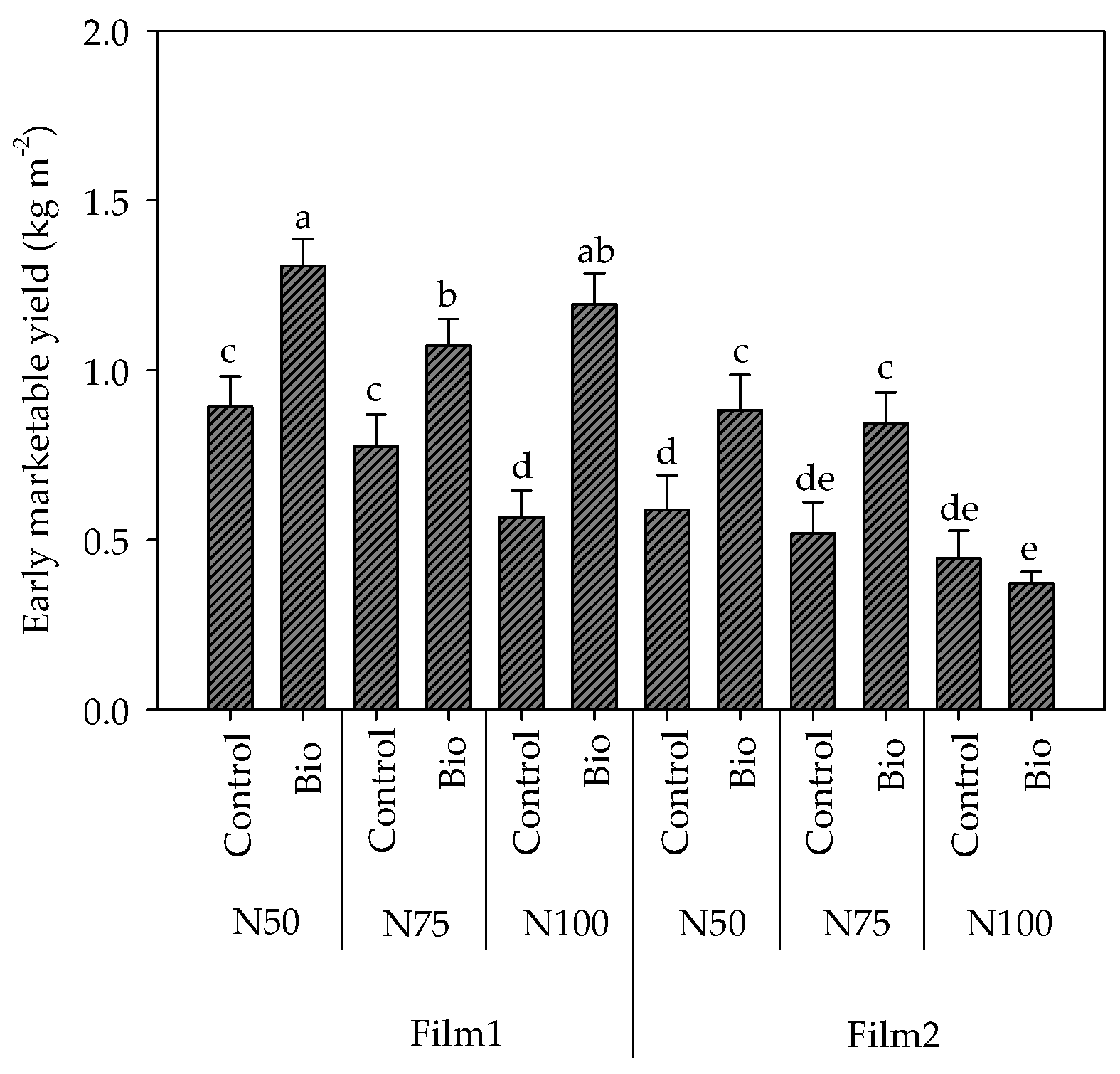
2.2. Early Production and Total Marketable Yield
3. Discussion
4. Materials and Methods
4.1. Plant Material and Crop Management
4.2. Plastic Covers, Nitrogen Rates, and Biostimulant Application
4.3. Time Distribution of Production and Total Yield, and Product Quality
4.3.1. Texture
4.3.2. Total Soluble Solids
4.3.3. Colour Parameters
4.3.4. Lycopene
4.3.5. Carotenoids
4.3.6. Antioxidant Activity
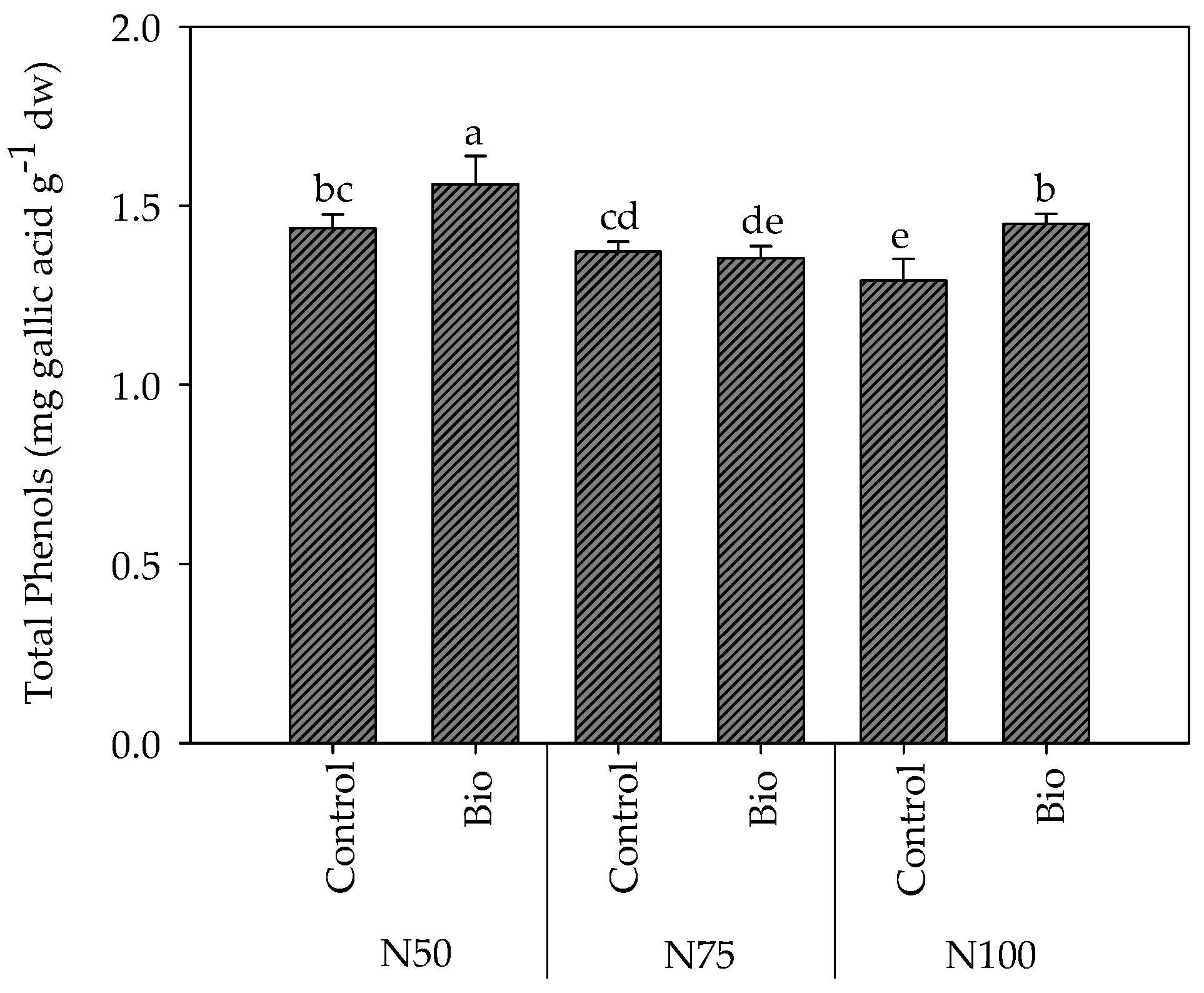
4.3.7. Total Phenols
4.3.8. Total Ascorbic Acid
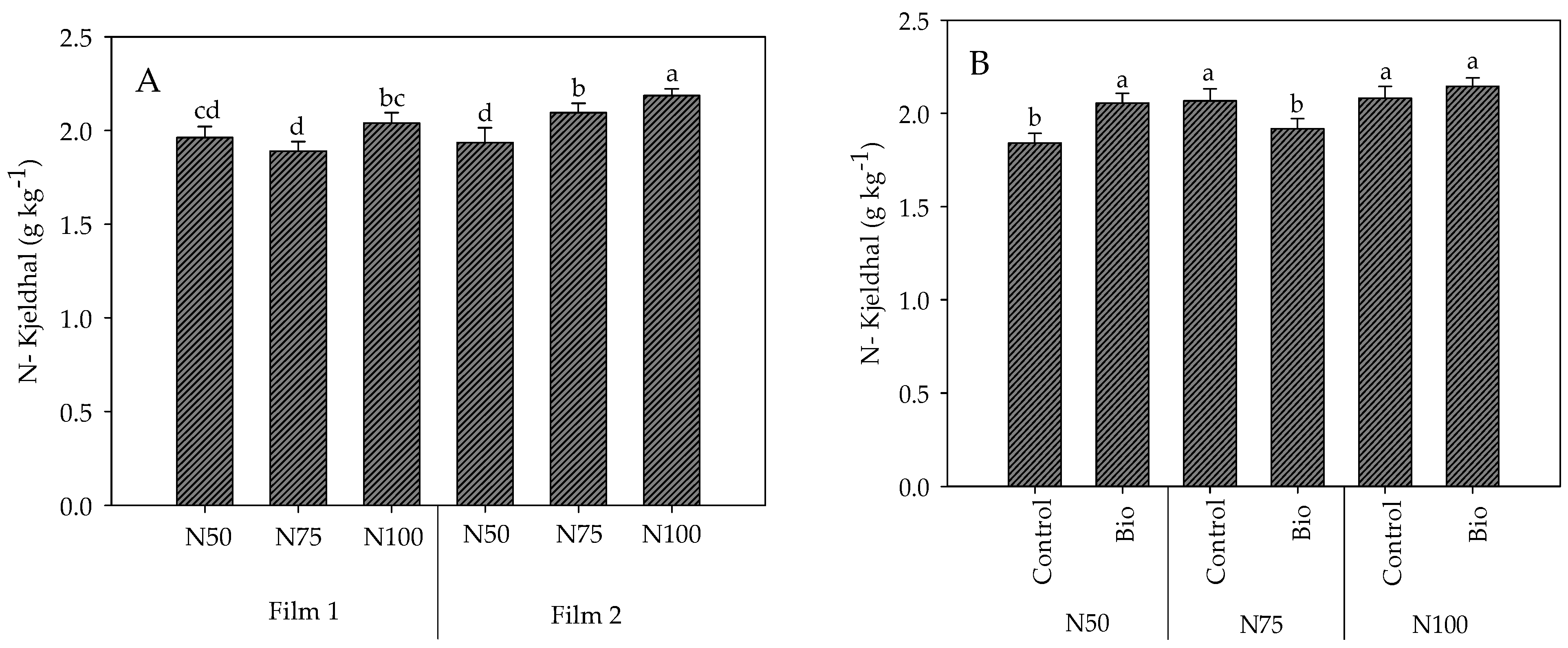
4.3.9. Nitrogen
4.4. Experimental Design and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Gruda, N.; Tanny, J. Protected Crops. Hortic. Plants People Places 2014, 1, 327–405. [Google Scholar] [CrossRef]
- Kumar, N.; Kumar, A.; Marwein, B.M.; Verma, D.K.; Jayabalan, I.; Kumar, A.; Kumar, N. Agricultural activities causing water pollution and its mitigation—A review. Inter. J. Modern Agric. 2021, 10, 590–609. [Google Scholar]
- Wato, T.; Amare, M.; Bonga, E.; Demand, B.B.O.; Coalition, B.B.R. The agricultural water pollution and its minimization strategies—A review. J. Resour. Dev. Manag. 2020, 64, 10–22. [Google Scholar]
- Paradiso, R.; Proietti, S. Light-quality manipulation to control plant growth and photomorphogenesis in greenhouse horticulture: The state of the art and the opportunities of modern led systems. J. Plant Growth Regul. 2021, 41, 742–780. [Google Scholar] [CrossRef]
- Li, T.; Yang, Q. Advantages of diffuse light for horticultural production and perspectives for further research. Front. Plant Sci. 2015, 6, 704. [Google Scholar] [CrossRef]
- Lamnatou, C.; Chemisana, D. Solar radiation manipulations and their role in greenhouse claddings: Fluorescent solar concentrators, photoselective and other materials. Renew. Sustain. Energy Rev. 2013, 27, 175–190. [Google Scholar] [CrossRef]
- Warren, W.J.; Hand, D.W.; Hannah, M.A. Light interception and photosynthetic efficiency in some glasshouse crops. J. Exp. Bot. 1992, 43, 363–373. [Google Scholar]
- Scarascia-Mugnozza, G.; Sica, C.; Russo, G. Plastic materials in European agriculture: Actual use and perspectives. J. Agric. Eng. 2011, 42, 15–28. [Google Scholar] [CrossRef]
- Hemming, S.; Dueck, T.A.; Janse, J.; Van Noort, F.R. The effect of diffuse light on crops. Acta Hortic. 2008, 801, 1293–1300. [Google Scholar] [CrossRef]
- Timmermans, G.H.; Hemming, S.; Baeza, E.; van Thoor, E.A.J.; Schenning, A.P.H.J.; Debije, M.G. Advanced optical materials for sunlight control in greenhouses. Adv. Opt. Mater. 2020, 8, 2000738. [Google Scholar] [CrossRef]
- Teitel, M.; Vitoshkin, H.; Geoola, F.; Karlsson, S.; Stahl, N. Greenhouse and screenhouse cover materials: Literature review and industry perspective. Acta Hortic. 2017, 1227, 31–44. [Google Scholar] [CrossRef]
- Rietra, R.P.J.J.; Heinen, M.; Dimkpa, C.O.; Bindraban, P.S. Effects of nutrient antagonism and synergism on yield and fertilizer use efficiency. Commun. Soil. Sci. Plant Anal. 2017, 48, 1895–1920. [Google Scholar] [CrossRef]
- He, X.; Maier, C.; Chavan, S.G.; Zhao, C.-C.; Alagoz, Y.; Cazzonelli, C.; Ghannoum, O.; Tissue, D.T.; Chen, Z.-H. Light-altering cover materials and sustainable greenhouse production of vegetables: A review. Plant Growth Regul. 2021, 95, 1–17. [Google Scholar] [CrossRef]
- Amirahmadi, E.; Ghorbani, M.; Moudrý, J.; Konvalina, P.; Kopecký, M. Impacts of Environmental Factors and Nutrients Management on Tomato Grown under Controlled and Open Field Conditions. Agronomy 2023, 13, 916. [Google Scholar] [CrossRef]
- Chaudhary, P.; Sharma, A.; Singh, B.; Nagpal, A.K. Bioactivities of phytochemicals present in tomato. J. Food Sci. Technol. 2018, 55, 2833–2849. [Google Scholar] [CrossRef] [PubMed]
- Dorais, M.; Ehret, D.L.; Papadopoulos, A.P. Tomato (Solanum lycopersicum) health components: From the seed to the consumer. Phytochem. Rev. 2008, 7, 231–250. [Google Scholar] [CrossRef]
- Di Mola, I.; Ottaiano, L.; Cozzolino, E.; El-Nakhel, C.; Rippa, M.; Mormile, P.; Corrado, G.; Rouphael, Y.; Mori, M. Assessment of yield and nitrate content of wall rocket grown under diffuse-light- or clear-plastic films and subjected to different nitrogen fertilization levels and biostimulant application. Horticulturae 2022, 8, 138. [Google Scholar] [CrossRef]
- Paradiso, R.; Di Mola, I.; Cozzolino, E.; Ottaiano, L.; El-Nakhel, C.; Rouphael, Y.; Mori, M. Nutrient and Nutraceutical Quality of Rocket as a Function of Greenhouse Cover film, Nitrogen Dose and Biostimulant Application. Agronomy 2023, 13, 638. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef]
- Takeno, K. Stress-induced flowering: The third category of flowering response. J. Exp. Botany 2016, 67, 4925–4934. [Google Scholar] [CrossRef]
- Moreno-Teruel, M.D.L.Á.; Molina-Aiz, F.D.; Peña-Fernández, A.; López-Martínez, A.; Valera-Martínez, D.L. The effect of diffuse film covers on microclimate and growth and production of tomato (Solanum lycopersicum L.) in a mediterranean greenhouse. Agronomy 2021, 11, 860. [Google Scholar] [CrossRef]
- Dueck, T.; Janse, J.; Li, T.; Kempkes, F.; Eveleens, B. Influence of diffuse glass on the growth and production of tomato. Acta Hortic. 2012, 956, 75–82. [Google Scholar] [CrossRef]
- Campania Region Fertilization Plan. Available online: http://www.agricoltura.regione.campania.it/concimazione/PRCFA_intro.html (accessed on 1 March 2023).
- Dueck, T.; Janse, J.; Kempkes, F.; Li, T.; Elings, A.; Hemming, S. Diffuse Light in Tomato; Wageningen University and Research, Stichting Dienst Landbouwkundig Onderzoek (DLO): Wageningen, Germany, 2012. [Google Scholar]
- Nawkar, G.; Maibam, P.; Park, J.; Sahi, V.; Lee, S.; Kang, C. UV-induced cell death in plants. Int. J. Mol. Sci. 2013, 14, 1608–1628. [Google Scholar] [CrossRef]
- Jenkins, G.I. Signal Transduction in responses to UV-B radiation. Annu. Rev. Plant Biol. 2009, 60, 407–431. [Google Scholar] [CrossRef] [PubMed]
- Loconsole, D.; Santamaria, P. UV Lighting in Horticulture: A Sustainable Tool for Improving Production Quality and Food Safety. Horticulturae 2021, 7, 9. [Google Scholar] [CrossRef]
- Ilić, Z.S.; Fallik, E. Light quality manipulation improves vegetable quality at harvest and postharvest: A review. Environ. Exp. Bot. 2017, 139, 79–90. [Google Scholar] [CrossRef]
- Ahmadi, L.; Hao, X.; Tsao, R. The effect of greenhouse covering materials on phytochemical composition and antioxidant capacity of tomato cultivars. J. Sci. Food Agric. 2018, 98, 4427–4435. [Google Scholar] [CrossRef] [PubMed]
- Szymańska, R.; Ślesak, I.; Orzechowska, A.; Kruk, J. Physiological and biochemical responses to high light and temperature stress in plants. Environ. Exp. Bot. 2017, 139, 165–177. [Google Scholar] [CrossRef]
- Bénard, C.; Gautier, H.; Bourgaud, F.; Grasselly, D.; Navez, B.; Caris-Veyrat, C.; Génard, M. Effects of low nitrogen supply on tomato (Solanum lycopersicum) fruit yield and quality with special emphasis on sugars, acids, ascorbate, carotenoids, and phenolic compounds. J. Agric. Food Chem. 2009, 57, 4112–4123. [Google Scholar] [CrossRef] [PubMed]
- Brandt, S.; Pék, Z.; Barna, É.; Lugasi, A.; Helyes, L. Lycopene content and colour of ripening tomatoes as affected by environmental conditions. J. Sci. Food Agric. 2006, 86, 568–572. [Google Scholar] [CrossRef]
- Helyes, L.; Lugasi, A.; Pék, Z. Effect of natural light on surface temperature and lycopene content of vine ripened tomato fruit. Can. J. Plant Sci. 2007, 87, 927–929. [Google Scholar] [CrossRef]
- Ahmed, B.; Zaidi, A.; Khan, M.S.; Rizvi, A.; Saif, S.; Shahid, M. Perspectives of plant growth promoting rhizobacteria in growth enhancement and sustainable production of tomato. In Microbial Strategies for Vegetable Production; Zaidi, A., Khan, M.S., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 125–149. [Google Scholar]
- He, Y.; Pantigoso, H.A.; Wu, Z.; Vivanco, J.M. Co-inoculation of Bacillus sp. and Pseudomonas putida at different development stages acts as a biostimulant to promote growth, yield and nutrient uptake of tomato. J. Appl. Microbiol. 2019, 127, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, V.; Romano, I.; Woo, S.L.; Stasio, E.D.; Lombardi, N.; Comite, E.; Maggio, A. Inoculation with a Microbial Consortium Increases Soil Microbial Diversity and Improves Agronomic Traits of Tomato Under Water and Nitrogen Deficiency. Front. Plant Sci. 2023, 14, 1304627. [Google Scholar] [CrossRef] [PubMed]
- Francesca, S.; Arena, C.; Hay Mele, B.; Schettini, C.; Ambrosino, P.; Barone, A.; Rigano, M.M. The use of a plant-based biostimulant improves plant performances and fruit quality in tomato plants grown at elevated temperatures. Agronomy 2020, 10, 363. [Google Scholar] [CrossRef]
- López-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrella, A. Trichoderma as biostimulant: Exploiting the multilevel properties of a plant beneficial fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Plantgest. Available online: https://plantgest.imagelinenetwork.com/it/varieta/orticole/pomodoro-da-mensa/sakura-f1/8267 (accessed on 2 October 2023).
- Hargreaves, G.H.; Samani, Z.A. Reference crop evapotranspiration from temperature. Appl. Eng. Agric. 1985, 1, 96–99. [Google Scholar] [CrossRef]
- Fertilidea Srl. Pompei, Naples, Italy. Available online: https://fertilidea.it/prodotto/515-lifestrong-vam-l.html (accessed on 2 October 2023).
- Sadler, G.; Davis, J.; Dezman, D. Rapid extraction of lycopene and-carotene from reconstituted tomato paste and pink grapefruit homogenates. J. Food Sci. 1990, 55, 1460–1461. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Fogliano, V.; Verde, V.; Randazzo, G.; Ritieni, A. Method for measuring antioxidant activity and its application to monitoring the antioxidant capacity of wines. J. Agric. Food Chem. 1999, 47, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Kampfenkel, K.; Vanmontagu, M.; Inze, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Bremner, J.M. Total nitrogen. In Methods of Soil Analysis, Part 2: Chemical and Microbiological Properties; Black, C.A., Evans, D.D., White, I.L., Ensminger, L.E., Clark, F.E., Eds.; American Society of Agronomy: Madison, WI, USA, 1965; Volume 2, pp. 1149–1178. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Early Marketable Yield | Total Marketable Yield | Texture | TSS | Dry Matter % | Dry Matter % | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| kg m−2 | n. Fruits | g Fruit−1 | kg m−2 | n. Fruits | g Fruit−1 | 3rd Harvest | 7th Harvest | |||
| Film (F) | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.05 | ns | ns | 0.001 | 0.05 |
| N Fertilization (N) | 0.001 | 0.001 | ns | 0.001 | 0.001 | 0.001 | ns | ns | ns | ns |
| Biostimulant (B) | 0.001 | 0.01 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | ns | 0.01 |
| F × N | ns | 0.05 | ns | ns | ns | ns | ns | ns | ns | ns |
| F × B | 0.05 | ns | ns | 0.001 | 0.01 | ns | ns | ns | ns | ns |
| N × B | ns | ns | 0.01 | 0.001 | 0.05 | ns | ns | ns | ns | ns |
| F × N × B | 0.05 | 0.01 | ns | 0.001 | 0.01 | ns | ns | ns | ns | ns |
| L* | a* | b* | Lycopene | Carotenoids | HAA | ABTS AA | Total Phenols | TAA | N-Kjeldhal | |
|---|---|---|---|---|---|---|---|---|---|---|
| Film (F) | 0.01 | 0.001 | ns | 0.001 | 0.001 | ns | 0.001 | 0.01 | 0.001 | 0.001 |
| N fertilization (N) | ns | 0.05 | ns | ns | ns | ns | ns | 0.001 | ns | 0.001 |
| Biostimulant (B) | ns | ns | ns | ns | ns | ns | 0.05 | 0.001 | 0.001 | ns |
| F × N | ns | ns | ns | 0.001 | ns | ns | ns | ns | ns | 0.05 |
| F × B | ns | ns | ns | 0.05 | ns | ns | ns | ns | ns | ns |
| N × B | ns | ns | ns | ns | ns | ns | ns | 0.01 | ns | 0.001 |
| F × N × B | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Early Marketable Yield | Total Marketable Yield | |||||
|---|---|---|---|---|---|---|
| n. Fruits m−2 | g Fruit−1 | n. Fruits m−2 | g Fruit−1 | |||
| Film1 | N50 | Control | 52.6 ± 4.6 a | 18.9 ± 1.6 | 115.3 ± 6.1 e | 22.3 ± 0.9 |
| N50 | Bio | 53.6 ± 4.6 a | 25.6 ± 1.1 | 144.2 ± 4.3 d | 25.7 ± 0.4 | |
| N75 | Control | 37.6 ± 3.0 cd | 21.6 ± 1.6 | 138.7 ± 4.2 d | 24.7 ± 0.8 | |
| N75 | Bio | 40.3 ± 4.1 cd | 27.1 ± 1.6 | 197.9 ± 7.4 b | 28.8 ± 0.7 | |
| N100 | Control | 22.9 ± 5.5 e | 26.2 ± 2.2 | 143.4 ± 5.2 d | 26.7 ± 0.8 | |
| N100 | Bio | 50.8 ± 3.2 ab | 24.7 ± 1.2 | 220.3 ± 8.6 a | 28.5 ± 0.7 | |
| Film2 | N50 | Control | 36.3 ± 4.7 cd | 16.8 ± 2.0 | 85.5 ± 5.5 f | 24.4 ± 0.7 |
| N50 | Bio | 43.4 ± 4.7 bc | 21.6 ± 1.3 | 118.9 ± 4.5 e | 24.7 ± 0.6 | |
| N75 | Control | 34.7 ± 7.9 d | 18.2 ± 1.4 | 115.0 ± 7.9 e | 23.9 ± 1.0 | |
| N75 | Bio | 36.3 ± 6.0 cd | 24.3 ± 0.8 | 157.9 ± 8.4 c | 26.7 ± 0.5 | |
| N100 | Control | 20.3 ± 3.9 e | 21.0 ± 1.2 | 133.7 ± 2.9 d | 26.1 ± 0.2 | |
| N100 | Bio | 18.9 ± 2.0 e | 20.4 ± 2.2 | 163.7 ± 4.3 c | 26.9 ± 0.4 | |
| Texture kg cm−2 | Total Soluble Solids (TSS, °Brix) | Dry Matter % 3rd Harvest | Dry Matter % 7th Harvest | |
|---|---|---|---|---|
| Film1 | 0.74 ± 0.02 | 9.66 ± 0.13 | 9.4 ± 0.1 b | 9.9 ± 0.2 b |
| Film2 | 0.74 ± 0.02 | 9.84 ± 0.13 | 10.6 ± 0.2 a | 10.5 ± 0.3 a |
| N50 | 0.75 ± 0.02 | 9.57 ± 0.15 | 10.1 ± 0.2 | 10.1 ± 0.2 |
| N75 | 0.73 ± 0.02 | 9.87 ± 0.16 | 9.9 ± 0.2 | 10.0 ± 0.3 |
| N100 | 0.74 ± 0.02 | 9.79 ± 0.16 | 9.9 ± 0.1 | 10.4 ± 0.3 |
| Control | 0.68 ± 0.01 b | 9.42 ± 0.13 b | 10.2 ± 0.2 | 10.6 ± 0.3 a |
| Bio | 0.80 ± 0.02 a | 10.07 ± 0.10 a | 9.8 ± 0.2 | 9.8 ± 0.2 b |
| L* | a* | b* | Carotenoids mg g−1 fw | |
|---|---|---|---|---|
| Film1 | 30.8 ± 0.3 b | 20.0 ± 0.4 b | 13.7 ± 0.3 | 0.136 ± 0.006 a |
| Film2 | 32.1 ± 0.3 a | 22.7 ± 0.5 a | 14.1 ± 0.2 | 0.110 ± 0.005 b |
| N50 | 31.5 ± 0.4 | 20.2 ± 0.7 b | 13.7 ± 03 | 0.123 ± 0.007 |
| N75 | 31.2 ± 0.4 | 21.9 ± 0.6 ab | 13.7 ± 0.2 | 0.124 ± 0.007 |
| N100 | 31.7 ± 0.4 | 22.0 ± 0.6 a | 14.2 ± 0.3 | 0.122 ± 0.009 |
| Control | 31.8 ± 0.3 | 21.2 ± 0.5 | 14.0 ± 0.2 | 0.120 ± 0.005 |
| Bio | 31.1 ± 0.3 | 21.5 ± 0.5 | 13.7 ± 0.2 | 0.126 ± 0.007 |
| HAA mmol ascorbic acid equ. 100 g−1 dw | ABTS AA mmol Trolox equ. 100 g−1 dw | TAA mg 100 g−1 fw | Total Phenols mg gallic acid g−1 dw | |
|---|---|---|---|---|
| Film1 | 8.83 ± 0.13 | 8.49 ± 0.26 b | 17.6 ± 1.2 b | 1.44 ± 0.04 a |
| Film2 | 8.71 ± 0.16 | 11.83 ± 0.22 a | 45.3 ± 1.2 a | 1.38 ± 0.04 b |
| N50 | 9.03 ± 0.13 | 10.19 ± 0.46 | 32.9 ± 4.9 | 1.50 ± 0.05 a |
| N75 | 8.71 ± 0.20 | 10.03 ± 0.74 | 29.6 ± 4.3 | 1.36 ± 0.05 b |
| N100 | 8.59 ± 0.19 | 10.26 ± 0.52 | 31.8 ± 4.0 | 1.37 ± 0.05 b |
| Control | 8.85 ± 0.14 | 9.81 ± 0.45 | 29.0 ± 3.6 b | 1.37 ± 0.04 b |
| Bio | 8.69 ± 0.15 | 10.51 ± 0.48 | 33.9 ± 3.4 a | 1.45 ± 0.04 a |
| Soil Properties | Measure Units | Mean Values |
|---|---|---|
| Sand | % | 91.0 |
| Silt | % | 4.5 |
| Clay | % | 4.5 |
| N—total (Kjeldahl method) | % | 0.101 |
| P2O5 (Olsen method) | ppm | 253.0 |
| K2O (Tetraphenylborate method) | ppm | 490.0 |
| Organic matter (Bichromate method) | % | 2.5 |
| pH | 7.4 |
| Agronomic Practices | DAT | ||||||
|---|---|---|---|---|---|---|---|
| Nitrogen fertilization | 30 | 60 | 90 | ||||
| Biostimulant application | 1 | 26 | 51 | 76 | 101 | ||
| Harvest time | 85 | 93 | 103 | 110 | 115 | 124 | 129 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paradiso, R.; Di Mola, I.; Ottaiano, L.; Cozzolino, E.; Pelosi, M.E.; Rippa, M.; Mormile, P.; Mori, M. Integrating Smart Greenhouse Cover, Reduced Nitrogen Dose and Biostimulant Application as a Strategy for Sustainable Cultivation of Cherry Tomato. Plants 2024, 13, 440. https://doi.org/10.3390/plants13030440
Paradiso R, Di Mola I, Ottaiano L, Cozzolino E, Pelosi ME, Rippa M, Mormile P, Mori M. Integrating Smart Greenhouse Cover, Reduced Nitrogen Dose and Biostimulant Application as a Strategy for Sustainable Cultivation of Cherry Tomato. Plants. 2024; 13(3):440. https://doi.org/10.3390/plants13030440
Chicago/Turabian StyleParadiso, Roberta, Ida Di Mola, Lucia Ottaiano, Eugenio Cozzolino, Maria Eleonora Pelosi, Massimo Rippa, Pasquale Mormile, and Mauro Mori. 2024. "Integrating Smart Greenhouse Cover, Reduced Nitrogen Dose and Biostimulant Application as a Strategy for Sustainable Cultivation of Cherry Tomato" Plants 13, no. 3: 440. https://doi.org/10.3390/plants13030440
APA StyleParadiso, R., Di Mola, I., Ottaiano, L., Cozzolino, E., Pelosi, M. E., Rippa, M., Mormile, P., & Mori, M. (2024). Integrating Smart Greenhouse Cover, Reduced Nitrogen Dose and Biostimulant Application as a Strategy for Sustainable Cultivation of Cherry Tomato. Plants, 13(3), 440. https://doi.org/10.3390/plants13030440