
The Effect of Heat Shock on Seed Dormancy Release and Germination in Two Rare and Endangered Astragalus L. Species (Fabaceae)

 ,
,  ,
,  , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Results
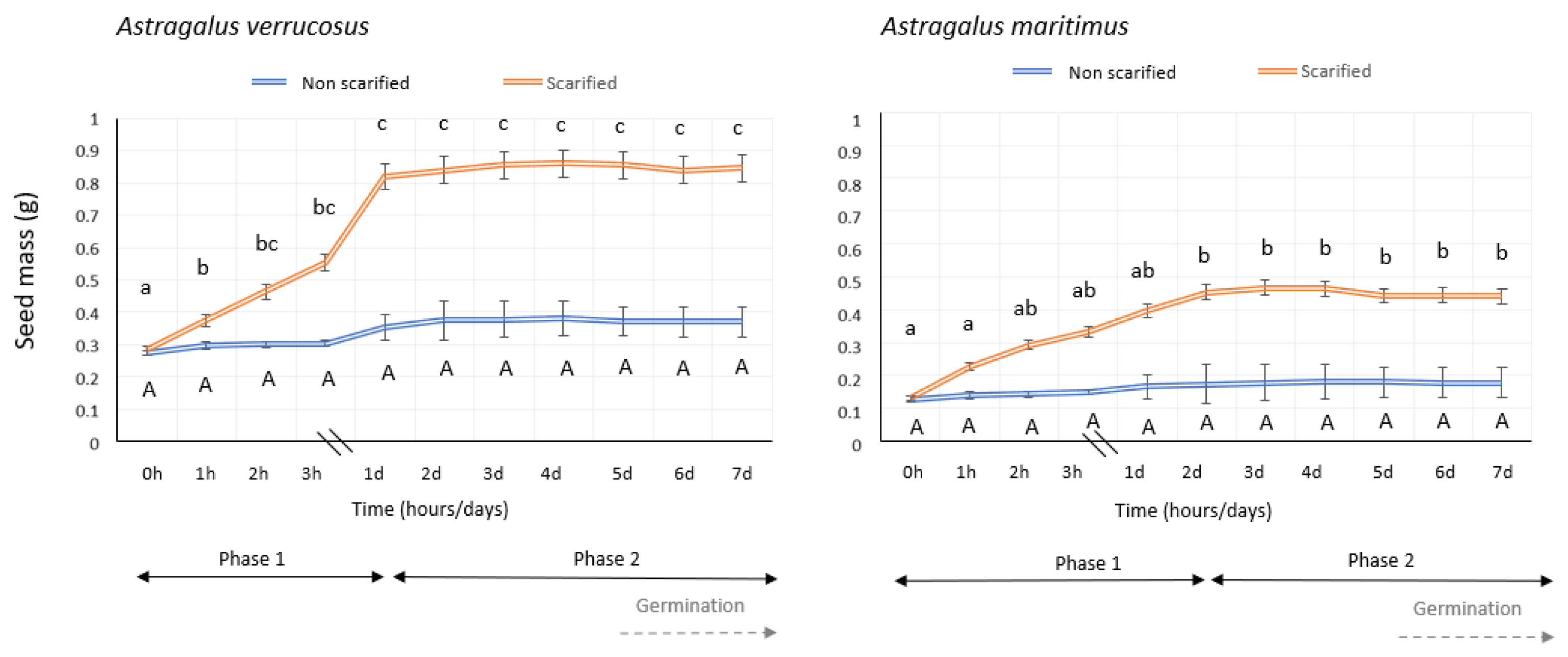
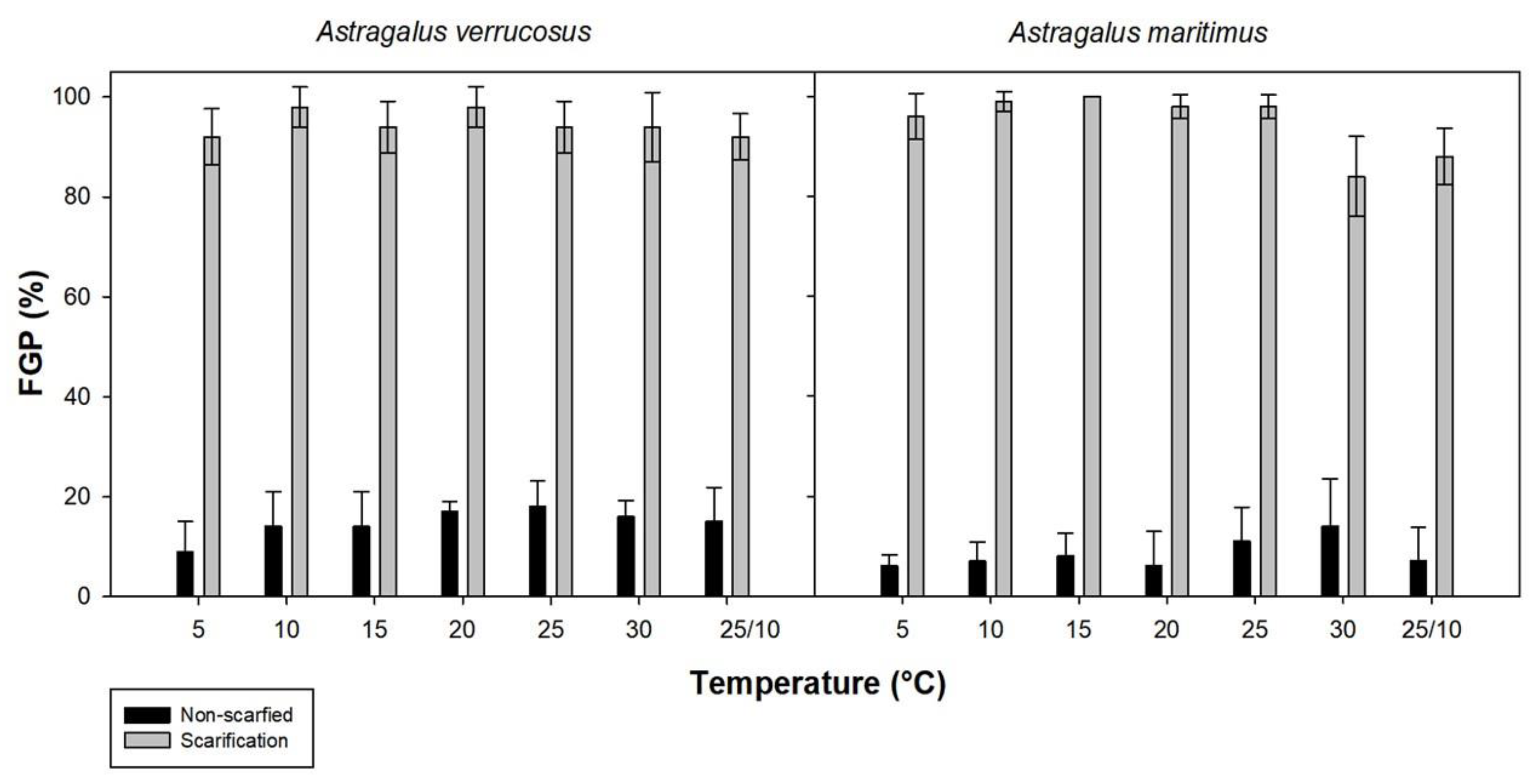
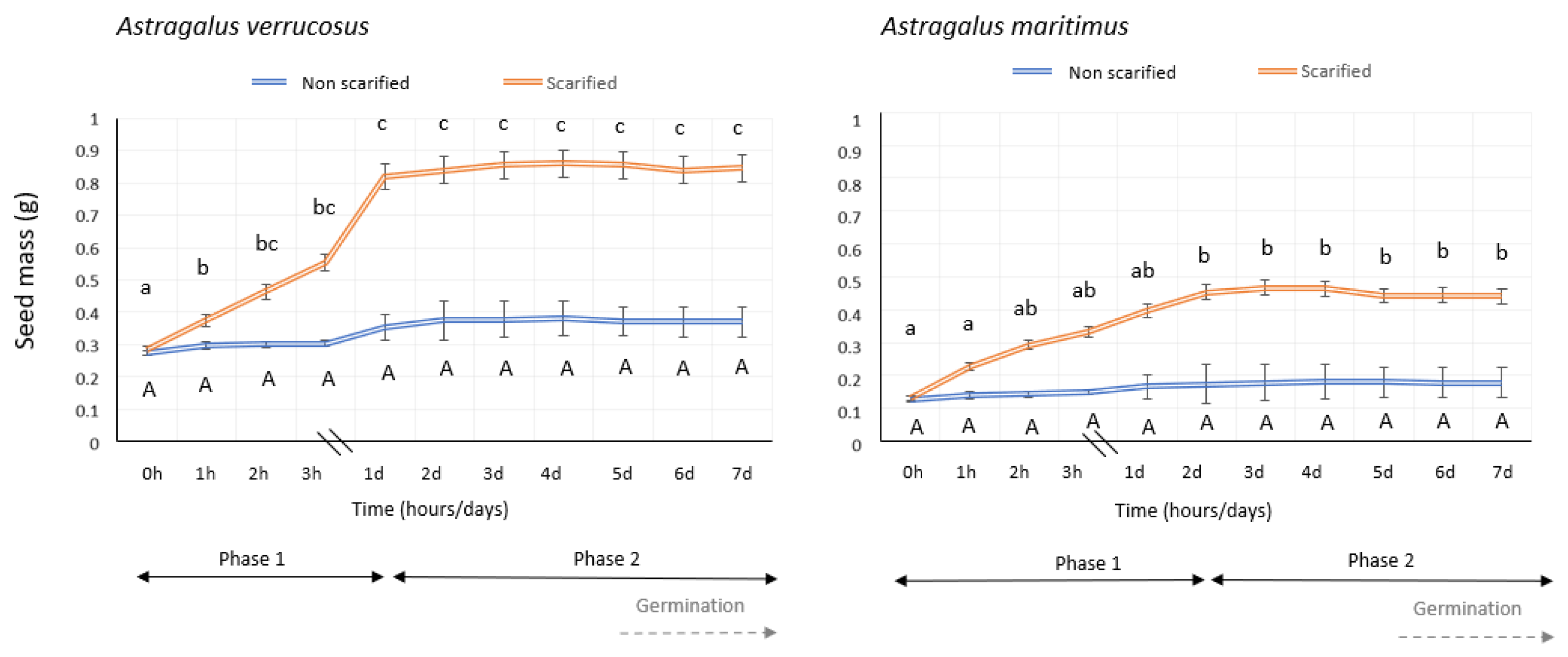
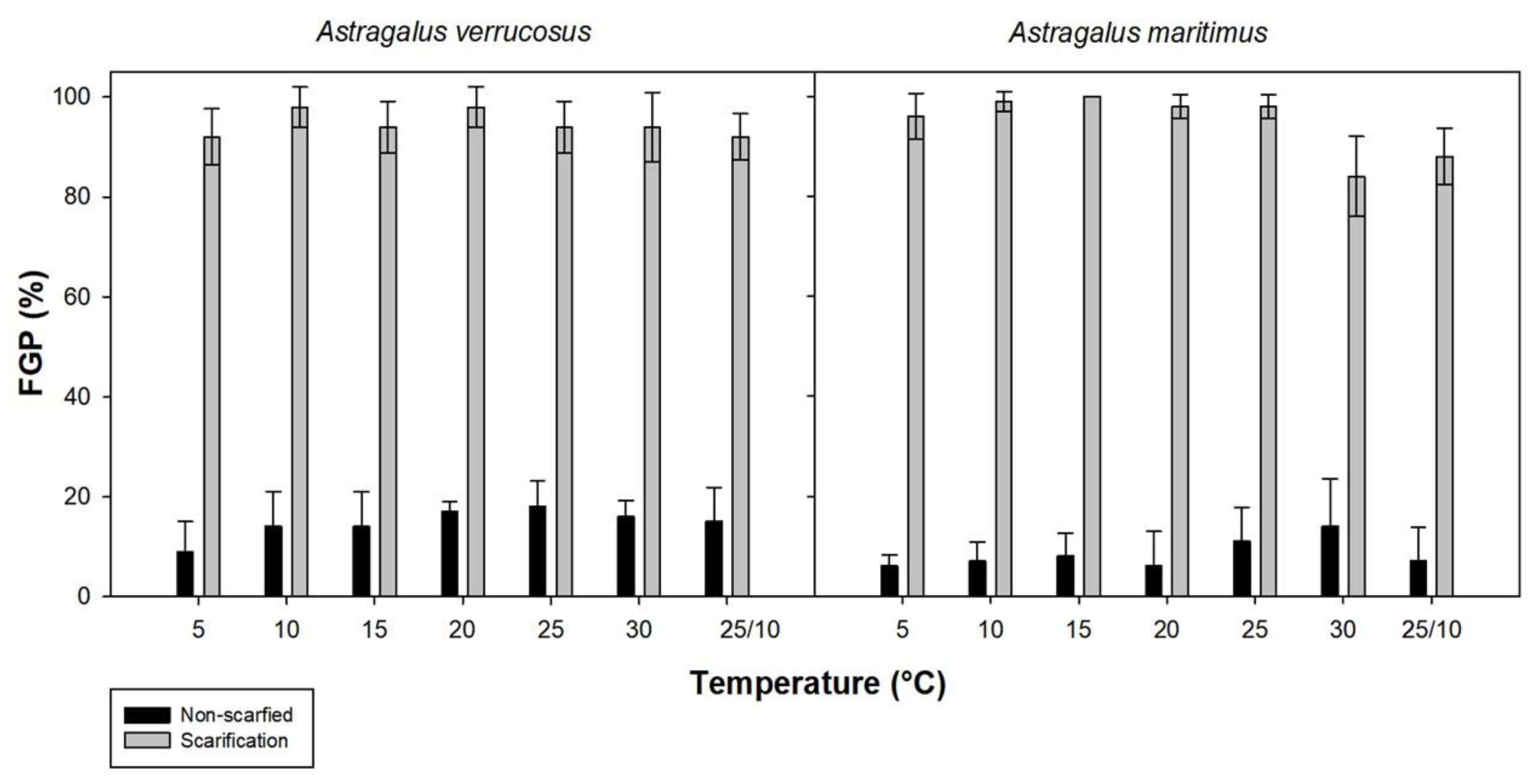
2.1. Imbibition of Water and Germination Trials
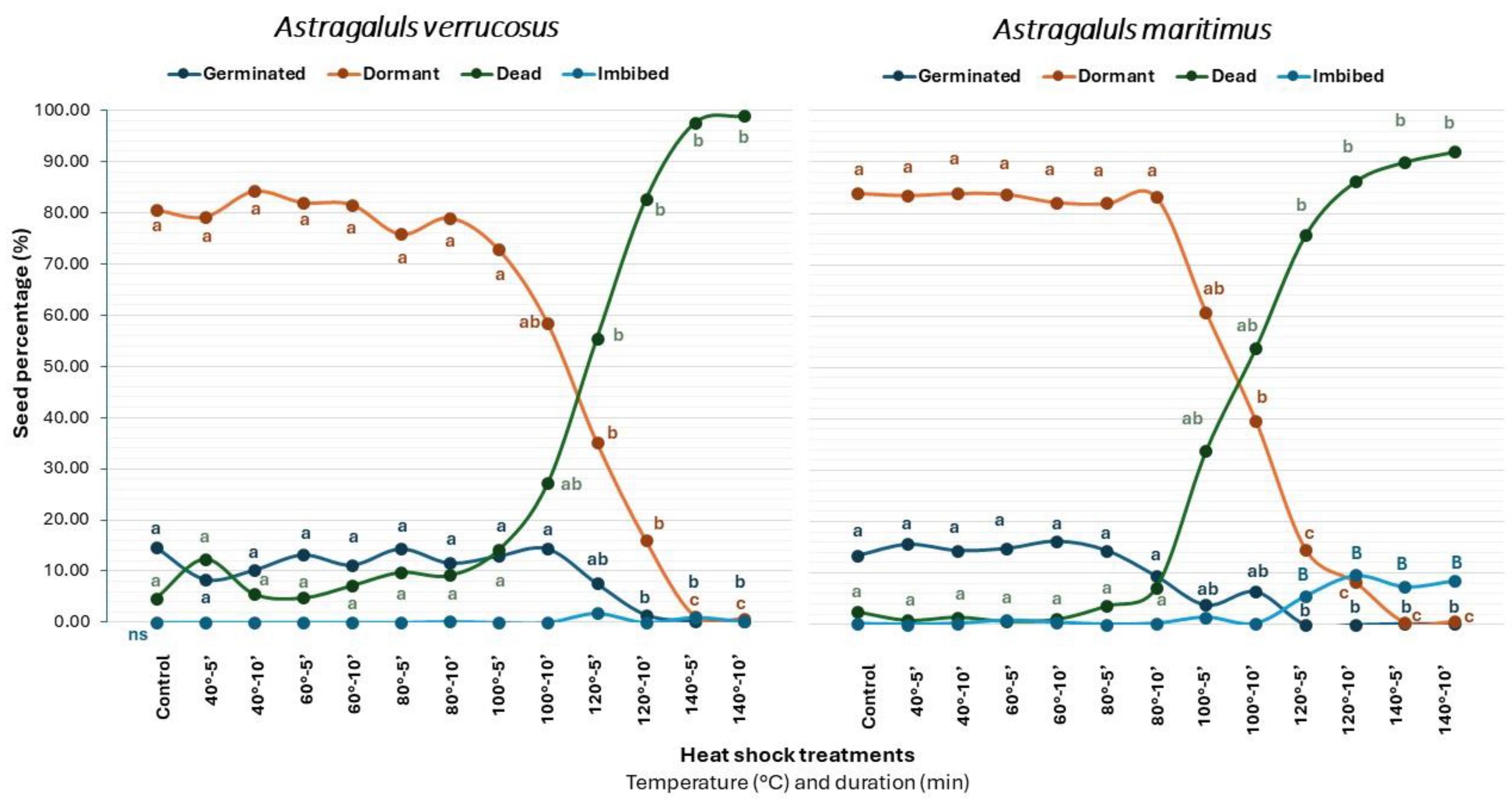
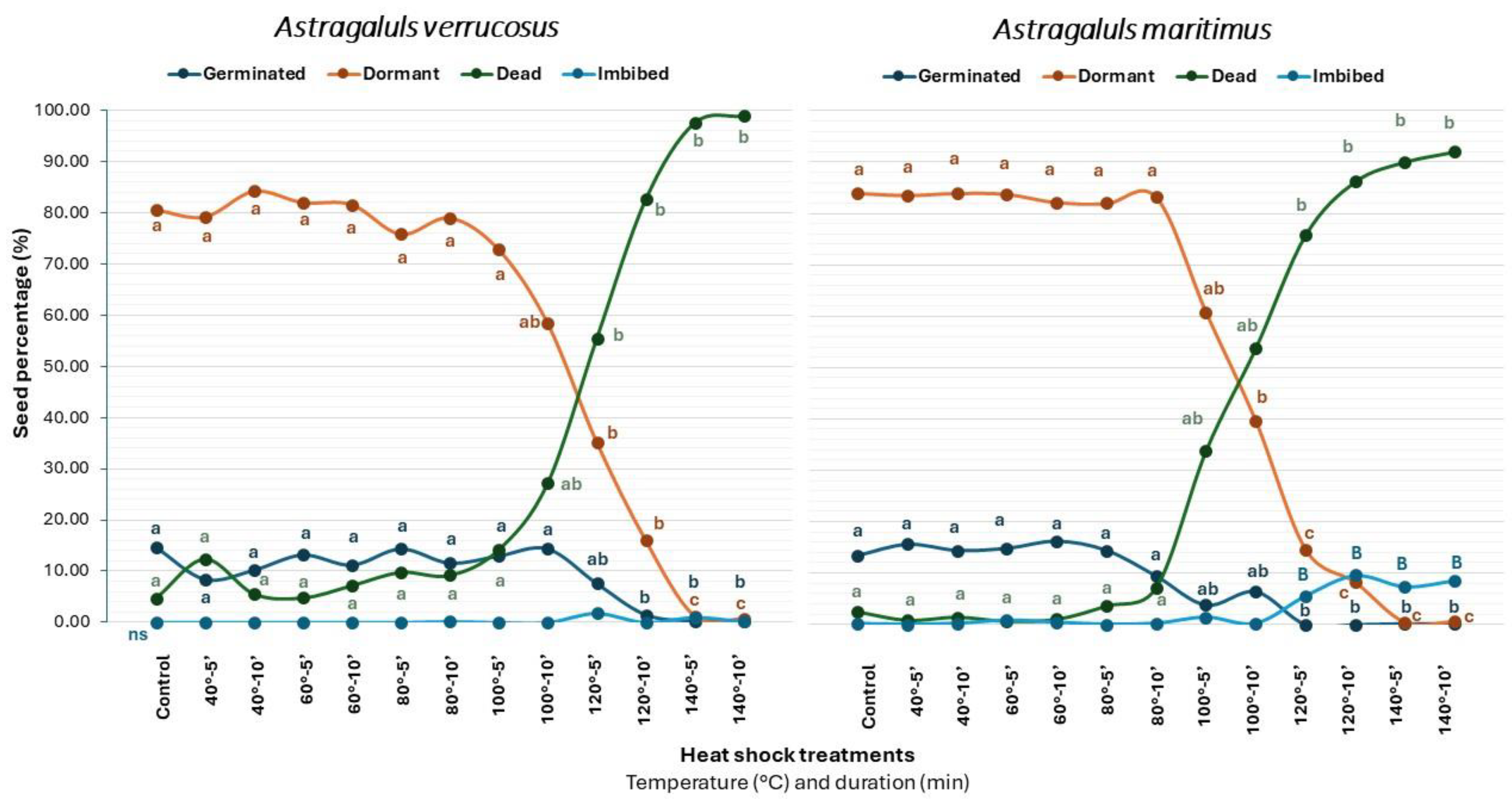
2.2. Heat Shock (HS)
3. Discussion
4. Materials and Methods
4.1. Study Species, Field Site Description, and Seed Collection
4.2. Imbibition of Water and Germination Trials
4.3. Heat Shock (HS)
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lock, J.M.; Simpson, K. Legumes of West Asia: A Checklist; Royal Botanic Gardens: Kew, UK, 1991. [Google Scholar]
- Mattana, E.; Grillo, O.; Venora, G.; Bacchetta, G. Germplasm image analysis of Astragalus maritimus and A. verrucosus of Sardinia (subgen. Trimeniaeus, Fabaceae). Anales Jard. Bot. Madrid 2008, 65, 149–155. [Google Scholar]
- Soltani, E.; Baskin, J.M.; Baskin, C.C.; Benakashani, F. A meta-analysis of the effects of treatments used to break dormancy in seeds of the megagenus Astragalus (Fabaceae). Seed Sci. Res. 2020, 30, 224–233. [Google Scholar] [CrossRef]
- Segura, F.; Vicente, M.J.; Franco, J.A.; Martínez-Sánchez, J.J. Effects of maternal environmental factors on physical dormancy of Astragalus nitidiflorus seeds (Fabaceae), a critically endangered species of SE Spain. Flora 2015, 216, 71–76. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Evolutionary considerations of claims for physical dormancy-break by microbial action and abrasion by soil particles. Seed Sci. Res. 2000, 10, 409–413. [Google Scholar] [CrossRef]
- Morrison, D.A.; Auld, T.D.; Rish, S.; Porter, C.; Mcclay, K. Patterns of testa-imposed seed dormancy in native Australian legumes. Ann. Bot. 1992, 70, 157–163. [Google Scholar] [CrossRef]
- Morrison, D.A.; Mcclay, K.; Porter, C.; Rish, S. The role of the lens in controlling heat-induced breakdown of testa-imposed dormancy in native Australian legumes. Ann. Bot. 1998, 82, 35–40. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Pausas, J.G.; Lamont, B.B. Fire-released seed dormancy-a global synthesis. Biol. Rev. 2022, 97, 1612–1639. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C.; Li, X. Taxonomy, anatomy and evolution of physical dormancy in seeds. Plant Species Biol. 2000, 15, 139–152. [Google Scholar] [CrossRef]
- Ooi, M.K.J.; Denham, A.J.; Santana, V.M.; Auld, T.D. Temperature thresholds of physically dormant seeds and plant functional response to fire: Variation among species and relative impact of climate change. Ecol. Evol. 2014, 4, 656–671. [Google Scholar] [CrossRef]
- Herranz, J.M.; Ferrandis, P.; Martínez-Sánchez, J.J. Influence of heat on seed germination of seven Mediterranean Leguminosae species. Plant Ecol. 1998, 136, 95–103. [Google Scholar] [CrossRef]
- Daibes, L.F.; Zupo, T.; Silveira, F.A.O.; Fidelis, A.A. field perspective on effects of fire and temperature fluctuation on Cerrado legume seeds. Seed Sci. Res. 2017, 27, 74–83. [Google Scholar] [CrossRef]
- Cochrane, M.A. Fire science for rainforests. Nature 2003, 42, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Stark, K.E.; Arsenault, A.; Bradfield, G.E. Soil seed banks and plant community assembly following disturbance by fire and logging in Interior Douglas-fir forests of south-central British Columbia. Canad. J. Bot. 2006, 84, 1548–1560. [Google Scholar] [CrossRef]
- Keeley, J.E.; Pausas, J.G.; Rundel, P.W.; Bond, W.J.; Bradstock, R.A. Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci. 2011, 8, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, L.C.; Pedrosa, M.; Borghetti, F. Heat shock effects on seed germination of five Brazilian savanna species. Plant Biol. 2013, 15, 152–157. [Google Scholar] [CrossRef]
- Stark, K.E.; Arsenault, A.; Bradfield, G.E. Variation in soil seed bank species composition of a dry coniferous forest: Spatial scale and sampling considerations. Plant Ecol. 2008, 197, 173–181. [Google Scholar] [CrossRef]
- Keeley, J.E. Seed germination and life history syndromes in the California chaparral. Bot. Rev. 1991, 57, 81–116. [Google Scholar] [CrossRef]
- Lamont, B.B.; Pausas, J.G.; He, T.; Witkowski, E.T.; Hanley, M.E. Fire as a selective agent for both serotiny and nonserotiny over space and time. Crit. Rev. Plant Sci. 2020, 39, 140–172. [Google Scholar] [CrossRef]
- Pausas, J.G.; Lamont, B.B. Ecology and biogeography in 3D: The case of the Australian Proteaceae. J. Biogeogr. 2018, 45, 1469–1477. [Google Scholar] [CrossRef]
- Moreira, B.; Pausas, J.G. Tanned or burned: The role of fire in shaping physical seed dormancy. PLoS ONE 2012, 7, e51523. [Google Scholar] [CrossRef]
- Lloret, F.; Peñuelas, J.; Estiarte, M. Experimental evidence of reduced diversity of seedlings due to climate modification in a Mediterranean-type community. Glob. Chang Biol. 2004, 10, 248–258. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Zema, D.A.; Carrà, B.G.; Cerdà, A.; Plaza-Alvarez, P.A.; Cózar, J.S.; Gonzalez-Romero, J.; Moya, D.; de las Heras, J. Short-term changes in infiltration between straw mulched and non-mulched soils after wildfire in Mediterranean forest ecosystems. Ecol. Eng. 2018, 122, 27–31. [Google Scholar] [CrossRef]
- Long, R.L.; Gorecki, M.J.; Renton, M.; Scott, J.K.; Colville, L.; Goggin, D.E.; Commander, L.E.; Westcott, D.A.; Cherry, H.; Finch-Savage, W.E. The ecophysiology of seed persistence: A mechanistic view of the journey to germination or demise. Biol. Rev. 2015, 90, 31–59. [Google Scholar] [CrossRef] [PubMed]
- Paula, S.; Pausas, J.G. Burning seeds: Germinative response to heat treatments in relation to resprouting ability. J. Ecol. 2008, 96, 543–552. [Google Scholar] [CrossRef]
- Luna, B.; Chamorro, D.; Pérez, B. Effect of heat on seed germination and viability in species of Cistaceae. Plant Ecol. Divers. 2019, 12, 151–158. [Google Scholar] [CrossRef]
- Luna, B. Fire and summer temperatures work together breaking physical seed dormancy. Sci. Rep. 2020, 10, 6031. [Google Scholar] [CrossRef]
- Pedrotta, T.; Gobet, E.; Schwörer, C.; Beffa, G.; Butz, C.; Henne, P.D.; Morales-Molino, C.; Pasta, S.; van Leeuwen, J.F.N.; Vogel, H.; et al. 8000 years of climate, vegetation, fire and land-use dynamics in the thermo-mediterranean vegetation belt of northern Sardinia (Italy). Veg. Hist. Archaeobot. 2021, 30, 789–813. [Google Scholar] [CrossRef]
- Kazancı, D.D.; Tavşanoğlu, Ç. Heat shock-stimulated germination in Mediterranean Basin plants in relation to growth form, dormancy type and distributional range. Folia Geobot. 2019, 54, 85–98. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: Burlington, MA, USA, 2014. [Google Scholar]
- Verdú, M.; Pausas, J.G. Fire drives phylogenetic clustering in Mediterranean Basin woody plant communities. J. Ecol. 2007, 95, 1316–1323. [Google Scholar] [CrossRef]
- Moreira, B.; Tormo, J.; Estrelles, E.; Pausas, J. Disentangling the role of heat and smoke as germination cues in Mediterranean Basin flora. Ann. Bot. 2010, 105, 627–635. [Google Scholar] [CrossRef]
- Luna, B.; Moreno, J.M.; Cruz, A.; Fernández-González, F. Heat-shock and seed germination of a group of Mediterranean plant species growing in a burned area: An approach based on plant functional types. Environ. Exp. Bot. 2007, 60, 324–333. [Google Scholar] [CrossRef]
- Bacchetta, G.; Fenu, G.; Mattana, E.; Pontecorvo, C. Ecological remarks on Astragalus maritimus and A. verrucosus, two threatened exclusive endemic species of Sardinia. Acta Bot. Gall. 2011, 158, 79–91. [Google Scholar] [CrossRef]
- Rodrigues-Junior, A.G.; Faria, J.M.; Vaz, T.A.; Nakamura, A.T.; José, A.C. Physical dormancy in Senna multijuga (Fabaceae: Caesalpinioideae) seeds: The role of seed structures in water uptake. Seed Sci. Res. 2014, 24, 147–157. [Google Scholar] [CrossRef]
- Nonogaki, H.; Bassel, G.W.; Bewley, J.D. Germination—Still a mystery. Plant Sci. 2010, 179, 574–581. [Google Scholar] [CrossRef]
- Bewley, J.D.; Black, M. Seeds: Physiology of Development and Germination; Springer Science & Business Media: Berlin, Germany, 1994. [Google Scholar]
- Pereira, W.V.S.; José, A.C.; Tonetti, O.A.O.; de Melo, L.A.; Faria, J.M.R. Imbibition curve in forest tree seeds and the triphasic pattern: Theory versus practice. S. Afr. J. Bot. 2022, 144, 105–114. [Google Scholar] [CrossRef]
- de Almeida, L.G.; Brandão, A.S.; Rossetto, C.A.V. Imbibition and white lupine seeds physiological quality affected by micronutrients supply. Ciência Rural 2015, 45, 612–619. [Google Scholar]
- Nunes, U.R.; Reis, M.S.; Giúdice, M.P.D.; Sediyama, T.; Sediyama, C.S. Embebição e qualidade fisiológica de sementes de soja submetidas ao condicionamento osmótico e condicionamento seguido de secagem. Ceres 2004, 51, 1–18. [Google Scholar]
- Araújo, P.C.; Torres, S.B.; Benedito, C.P.; Paiva, E.P.D. Priming and seed vigor in gherkins (Cucumis anguria L.). Rev. Bras. Sementes 2011, 33, 482–489. [Google Scholar] [CrossRef]
- Custódio, C.C.; Ambiel, A.C.; Rodrigues, D.Z.; de Agostini, E.A.T.; Factur, V.D.; Pavanelli, L.E. Film coating of intact and scarified Brachiaria brizantha (Hochst. ex A. Rich.) Stapf seeds. Pesqui. Agropecu. Trop. 2011, 41. [Google Scholar] [CrossRef]
- Jaganathan, G.K.; Li, J.; Biddick, M.; Han, K.; Song, D.; Yang, Y.; Liu, B. Mechanisms underpinning the onset of seed coat impermeability and dormancy-break in Astragalus adsurgens. Sci. Rep. 2019, 9, 9695. [Google Scholar] [CrossRef]
- Jaganathan, G.K.; Yule, K.; Liu, B. On the evolutionary and ecological value of breaking physical dormancy by endozoochory. Perspect. Plant Ecol. Evol. Syst. 2016, 22, 11–22. [Google Scholar] [CrossRef]
- Bacchetta, G.; Guarino, R.; Brullo, S.; Giusso Del Galdo, G. Indagine fitosociologica sulle praterie a Brachypodium retusum (Pers.) Beauv. della Sardegna. Parlatorea 2005, 7, 27–38. [Google Scholar]
- EU. Council Directive 92/43/EEC on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Commun. L 1992, 206, 7–50. [Google Scholar]
- Bacchetta, G.; Fenu, G.; Mattana, E.; Piotto, B.; Virevaire, M. Manuale per la Raccolta, Studio, Conservazione E Gestione Ex Situ del Germoplasma; Manuali e Linee guida APAT: Roma, Italy, 2006; p. 244. [Google Scholar]
- Bacchetta, G.; Bueno Sánchez, A.; Fenu, G.; Jiménez-Alfaro, B.; Mattana, E.; Piotto, B.; Virevaire, M. Conservación Ex Situ de Plantas Silvestres; La Caixa: Principado de Asturias, Spain, 2008; p. 375. [Google Scholar]
- Porceddu, M.; Santo, A.; Orru, M.; Meloni, F.; Ucchesu, M.; Picciau, R.; Sarigu, M.; Cuena-Lombraña, A.; Podda, L.; Sau, S.; et al. Seed conservation actions for the preservation of plant diversity: The case of the Sardinian Germplasm Bank (BG-SAR). Plant Sociol. 2017, 54, 111–117. [Google Scholar] [CrossRef]
- ISTA—International Rules for Seed Testing, 2006th ed; The International Seed Testing Association (ISTA): Bassersdorf, Zwitserland, 2006.
- Crawley, M.J. The R Book; John Wiley & Sons Ltd.: Chichester, UK, 2007. [Google Scholar]
- R Development Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2023. Available online: https://www.rproject.org/ (accessed on 4 September 2023).

{kind=link}
{kind=link}
{kind=link}
| Coefficients: | Estimate | Std. Error | t Value | p Value |
|---|---|---|---|---|
| (Intercept) | −1.743 | 0.103 | −16.813 | <0.001 *** |
| Treatment | 0.934 | 0.131 | 7.148 | <0.001 *** |
| Time | 0.028 | 0.017 | 1.601 | 0.11 |
| Species | 0.973 | 0.132 | 7.429 | <0.001 *** |
| Treatment x Time | 0.051 | 0.022 | 2.344 | 0.019 * |
| Treatment x Species | 0.009 | 0.173 | 0.053 | 0.957 |
| Time x Species | 0.005 | 0.022 | 0.231 | 0.817 |
| Treatment x Time x Species | 0.134 | 0.031 | 4.283 | <0.001 *** |
| Estimate | Std. Error | t Value | p Value | |
|---|---|---|---|---|
| (Intercept) | −280.185 | 0.596 | −4.696 | <0.001 *** |
| Treatment | 1048.816 | 197.371 | 5.314 | <0.001 *** |
| Temperature | 0.039 | 0.024 | 1.582 | 0.119 |
| Species | 100.929 | 0.793 | 1.272 | 0.208 |
| Treatment x Temperature | −0.197 | 0.074 | −2.671 | 0.009 ** |
| Treatment x Species | −495.658 | 222.635 | −2.226 | 0.030 * |
| Temperature x Species | −0.033 | 0.033 | −0.995 | 0.324 |
| Treatment x Temperature x Species | 0.152 | 0.085 | 1.787 | 0.079 |
| F-Tests | Df | Sum Sq | Mean Sq | F | p Value | |
|---|---|---|---|---|---|---|
| Germinated Percentage | Treatment | 12 | 1.585 | 0.132 | 36.137 | 0.001 *** |
| Species | 1 | 0.009 | 0.009 | 2.546 | 0.111 | |
| Species x Treatment | 12 | 0.301 | 0.251 | 6.867 | <0.05 * | |
| Dead Percentage | Treatment | 12 | 67.730 | 5.644 | 678.256 | <0.001*** |
| Species | 1 | 0.020 | 0.024 | 2.929 | 0.680 | |
| Treatment x Species | 12 | 2.450 | 0.204 | 24.503 | <0.05 * | |
| Dormant Percentage | Treatment | 12 | 55.050 | 4.587 | 613.39 | <0.001*** |
| Species | 1 | 0.130 | 0.128 | 17.12 | <0.05 * | |
| Treatment x Species | 12 | 1.220 | 0.101 | 13.55 | <0.05 * | |
| Imbibed Percentage | Treatment | 12 | 0.189 | 0.015 | 6.34 | <0.05 * |
| Species | 1 | 0.074 | 0.074 | 30.115 | <0.05 * | |
| Treatment x Species | 12 | 0.146 | 0.012 | 4.869 | <0.05 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuena Lombraña, A.; Dessì, L.; Podda, L.; Fois, M.; Luna, B.; Porceddu, M.; Bacchetta, G. The Effect of Heat Shock on Seed Dormancy Release and Germination in Two Rare and Endangered Astragalus L. Species (Fabaceae). Plants 2024, 13, 484. https://doi.org/10.3390/plants13040484
Cuena Lombraña A, Dessì L, Podda L, Fois M, Luna B, Porceddu M, Bacchetta G. The Effect of Heat Shock on Seed Dormancy Release and Germination in Two Rare and Endangered Astragalus L. Species (Fabaceae). Plants. 2024; 13(4):484. https://doi.org/10.3390/plants13040484
Chicago/Turabian StyleCuena Lombraña, Alba, Ludovica Dessì, Lina Podda, Mauro Fois, Belén Luna, Marco Porceddu, and Gianluigi Bacchetta. 2024. "The Effect of Heat Shock on Seed Dormancy Release and Germination in Two Rare and Endangered Astragalus L. Species (Fabaceae)" Plants 13, no. 4: 484. https://doi.org/10.3390/plants13040484
APA StyleCuena Lombraña, A., Dessì, L., Podda, L., Fois, M., Luna, B., Porceddu, M., & Bacchetta, G. (2024). The Effect of Heat Shock on Seed Dormancy Release and Germination in Two Rare and Endangered Astragalus L. Species (Fabaceae). Plants, 13(4), 484. https://doi.org/10.3390/plants13040484