

Allelopathy and Identification of Five Allelochemicals in the Leaves of the Aromatic Medicinal Tree Aegle marmelos (L.) Correa


Abstract

1. Introduction
2. Results
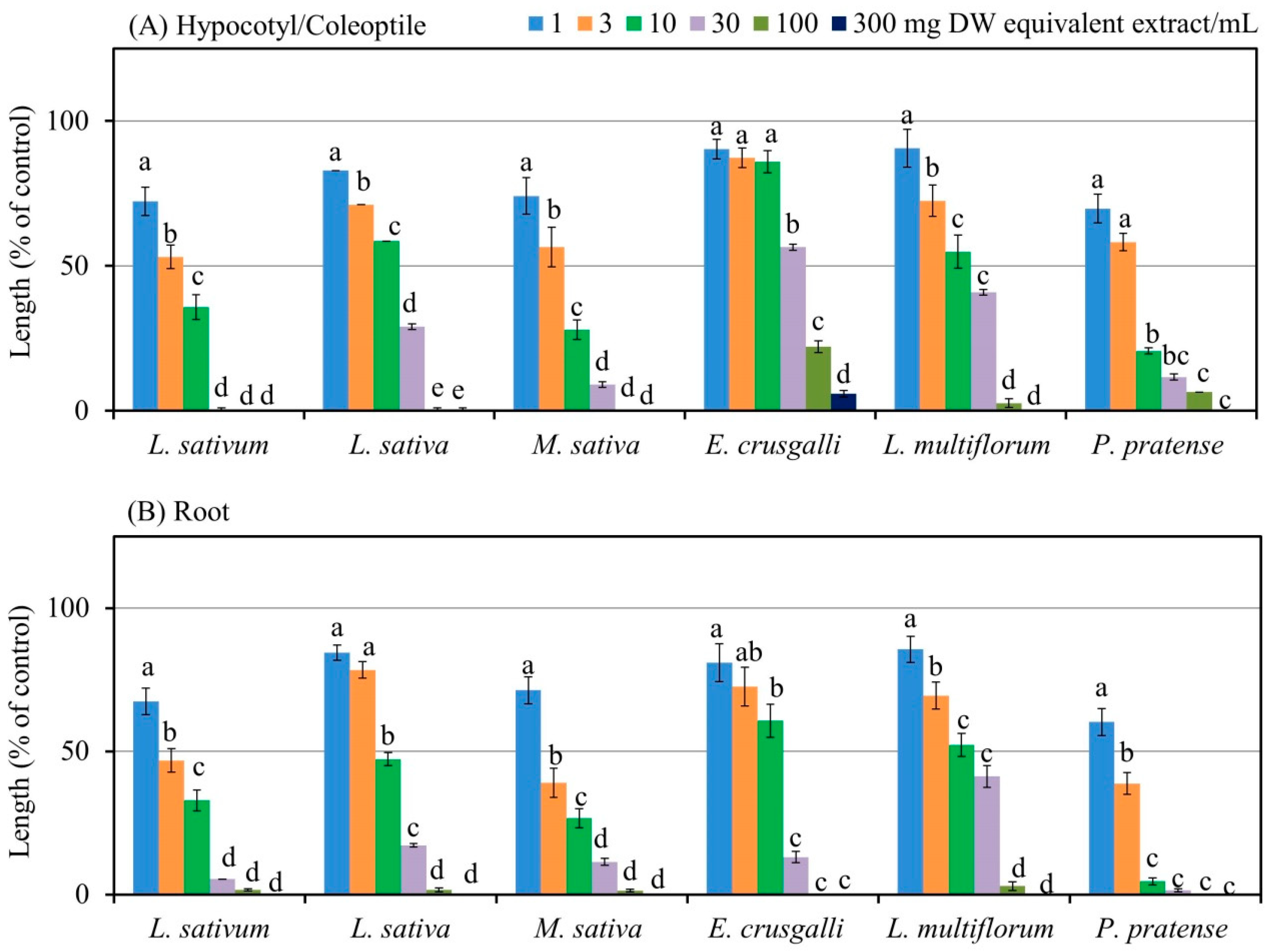
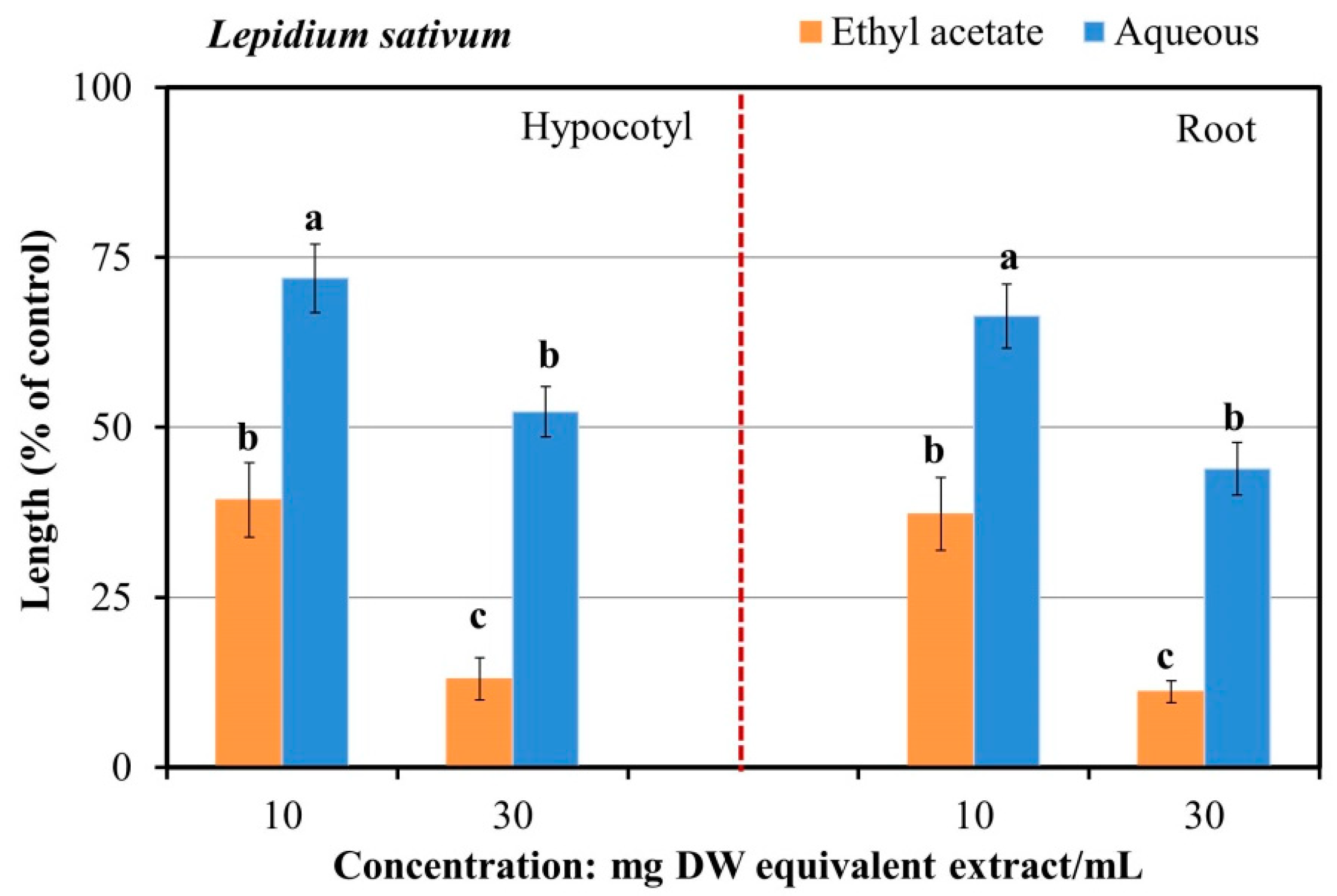
2.1. Growth-Suppressive Activity of A. marmelos
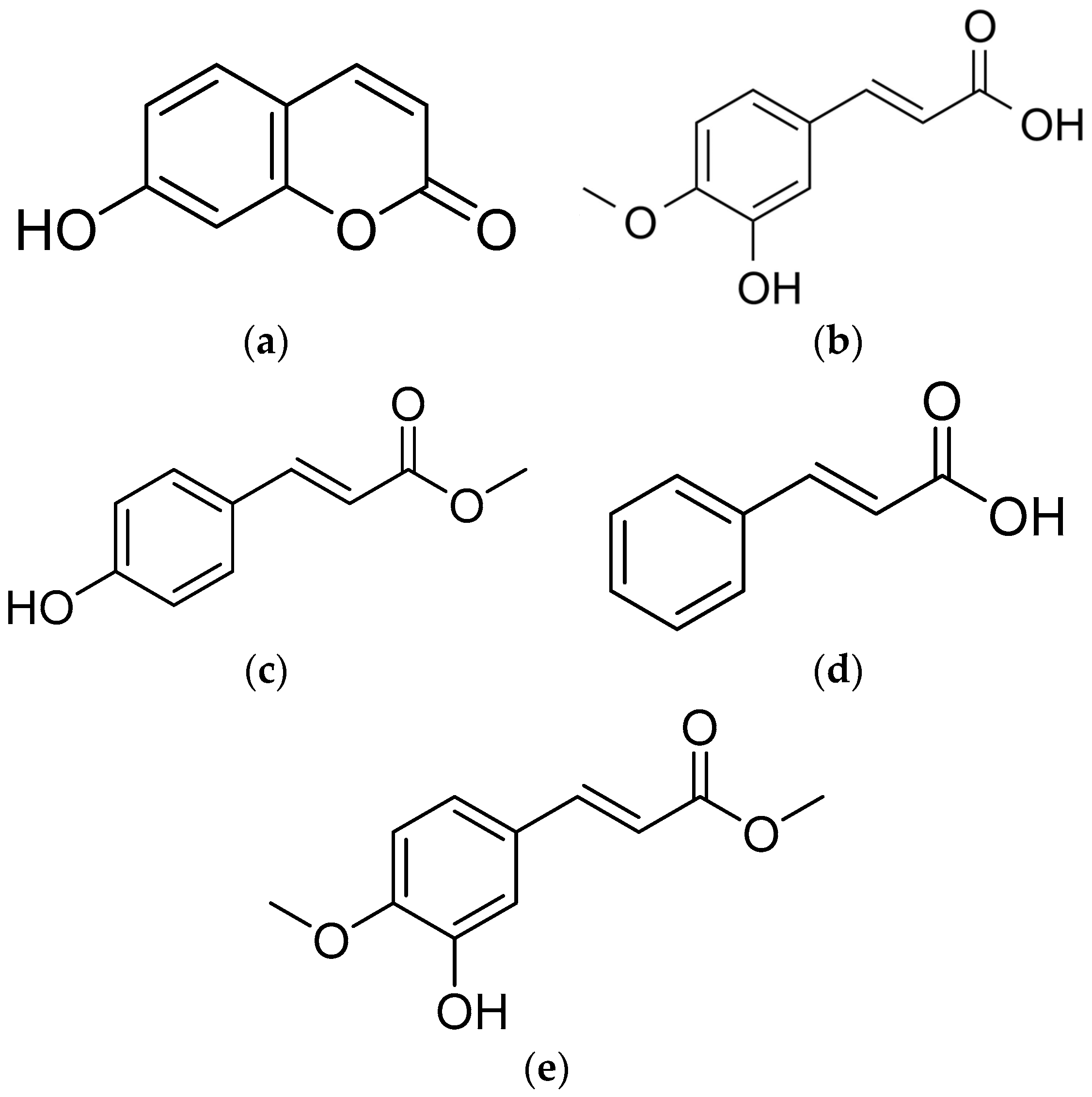
2.2. Identification of the Five Active Compounds
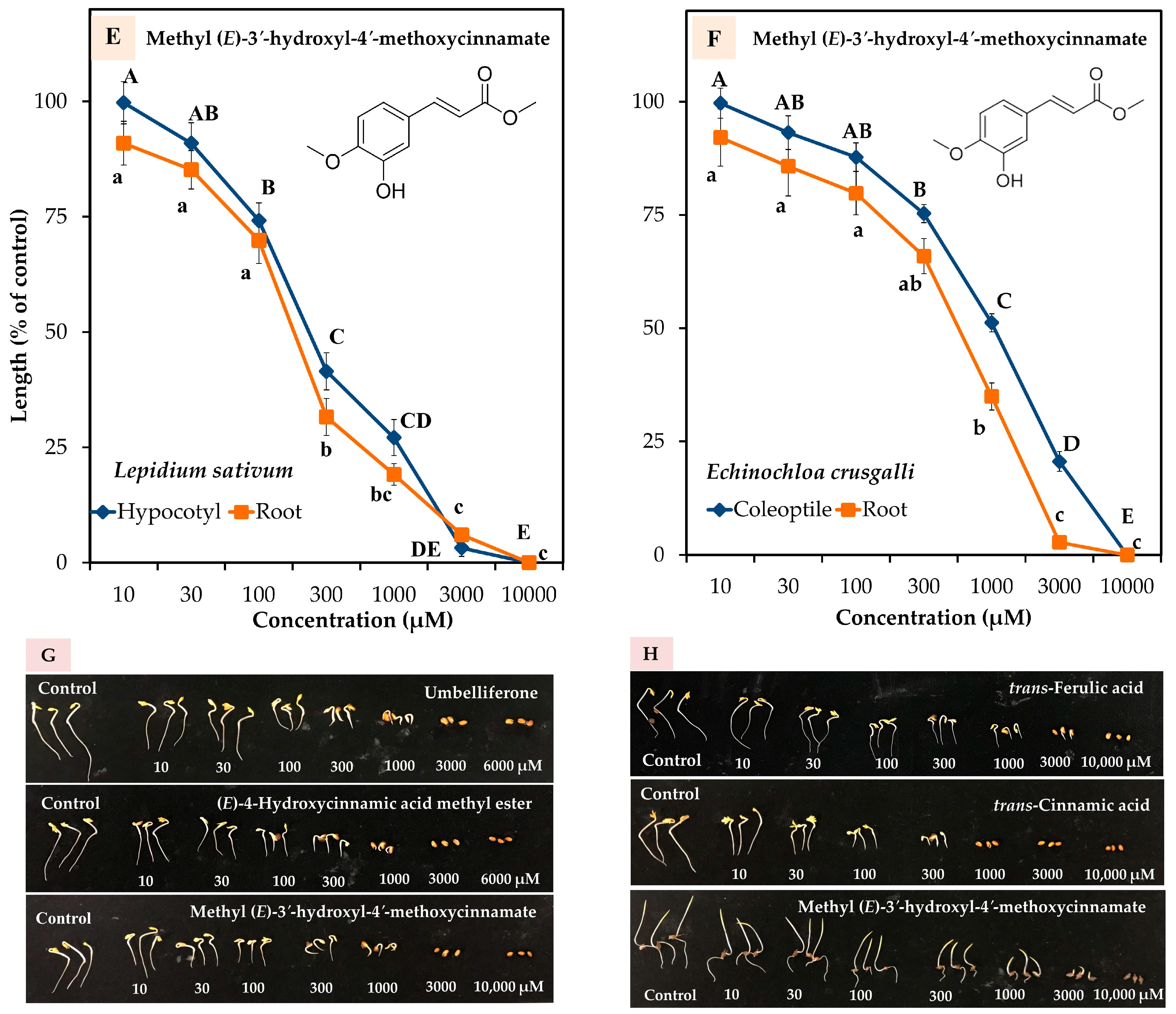
2.3. Allelopathic Effect of the Five Compounds
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Test Plant Species
4.2. Extraction and Growth Bioassay
4.3. Isolation and Purification of the Active Compounds
4.4. Bioassays of the Identified Compounds
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stokstad, E.; Grullon, G. Infographic: Pesticide planet. Science 2013, 341, 730–731. [Google Scholar]
- Xuan, T.D.; Tsuzuki, E.; Tawata, S.; Khanh, T.D. Methods to determine allelopathic potential of crop plants for weed control. Allelopath. J. 2004, 13, 149–164. [Google Scholar]
- Friesen, L.F.; Jones, T.L.; Acker, R.C.V.; Morrison, I.N. Identification of Avena fatua populations resistant to imazamethabenz, flamprop and fenoxaprop-P. Weed Sci. 2000, 48, 532–540. [Google Scholar] [CrossRef]
- Scarabel, L.; Varotto, S.; Sattin, M.A. European biotype of Amaranthus retroflexus cross-resistant to ALS inhibitors and response to alternative herbicides. Weed Res. 2007, 47, 527–533. [Google Scholar] [CrossRef]
- Kumar, J.I.N.; Amb, M.K.; Bora, A. Chronic response of Anabaena fertilissima on growth, metabolites and enzymatic activities by chlorophenoxy herbicide. Pestic. Biochem. Physiol. 2010, 98, 168–174. [Google Scholar] [CrossRef]
- Rawat, L.S.; Maikhuri, R.K.; Bahuguna, Y.M.; Jha, N.K.; Phondani, P.C. Sunflower allelopathy for weed control in agriculture systems. J. Crop Sci. Biotechnol. 2017, 20, 45–46. [Google Scholar] [CrossRef]
- Scavo, A.; Abbate, C.; Mauromicale, G. Plant allelochemicals: Agronomic, nutritional and ecological relevance in the soil system. Plant Soil 2019, 442, 23–48. [Google Scholar] [CrossRef]
- Fujii, Y.; Hiradate, S. Allelopathy: New concepts and methodology. Sci. Publ. 2007, 36, 173–183. [Google Scholar]
- Singh, H.P.; Kohli, R.K.; Batish, D.R. Allelopathy in agroecosystems: An overview. J. Crop Prod. 2001, 4, 1–41. [Google Scholar] [CrossRef]
- Seigler, D.S. Chemistry and mechanisms of allelopathic interactions. Agron. J. 1996, 88, 876–885. [Google Scholar] [CrossRef]
- Siqueira, J.O.; Nair, M.G.; Hammerschmidt, R.; Safir, G.R.; Putnam, A.R. Significance of phenolic compounds in plant–soil–microbial systems. Crit. Rev. Plant Sci. 1991, 10, 63–121. [Google Scholar] [CrossRef]
- Francisco, A.M.; Molinillo, J.M.G.; Rosa, M.V.; Galindo, J.C.G. Allelopathy—A natural alternative for weed control. Pest Manag. Sci. 2007, 63, 327–348. [Google Scholar]
- John, J.; Sarada, S. Role of phenolics in allelopathic interactions. Allelopath. J. 2012, 29, 215–230. [Google Scholar]
- Islam, A.K.M.M.; Kato-Noguchi, H. Phytotoxic activity of Ocimum tenuiflorum extracts on germination and seedling growth of different plant species. Sci. World J. 2014, 2014, 676242. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Furukawa, M.; Hayakawa, Y.; Sugawara, K.; Shibuya, T. Survey of Japanese medicinal plants for the detection of allelopathic properties. Weed Res. Jpn. 1991, 36, 36–42. [Google Scholar]
- Sun, W.; Shahrajabian, M.H. Therapeutic potential of phenolic compounds in medicinal plants—Natural health products for human health. Molecules 2023, 28, 1845. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Salam, M.A.; Ohno, O.; Suenaga, K. Nimbolide B and nimbic acid B, phytotoxic substances in neem leaves with allelopathic activity. Molecules 2014, 19, 6929–6940. [Google Scholar] [CrossRef]
- Krumsri, R.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Phytotoxic effects of Senna garrettiana and identification of phytotoxic substances for the development of bioherbicides. Agriculture 2022, 12, 1338. [Google Scholar] [CrossRef]
- Sharma, P.C.; Bhatia, V.; Bansal, N.; Sharma, A. A review on bael tree. Indian J. Nat. Prod. Resour. 2007, 6, 171–178. [Google Scholar]
- Akbar, S. Aegle marmelos (L.) Correa (Rutaceae). In Handbook of 200 Medicinal Plants; Springer: Cham, Switzerland, 2020; pp. 109–122. [Google Scholar]
- Bhar, K.; Mondal, S.; Suresh, P. An eye-catching review of Aegle marmelos L. (golden apple). Pharmacogn. J. 2019, 11, 207–224. [Google Scholar] [CrossRef]
- Kintzios, S.E. Terrestrial plant-derived anticancer agents and plant species used in anticancer research. Crit. Rev. Plant Sci. 2006, 25, 79–113. [Google Scholar] [CrossRef]
- Lambole, V.B.; Murti, K.; Kumar, U.; Sandipkumar, B.P.; Gajera, V. Phytopharmacological properties of Aegle marmelos as a potential medicinal tree: An overview. Int. J. Pharm. Sci. Rev. Res. 2010, 5, 67–72. [Google Scholar]
- Baliga, M.S.; Bhat, H.P.; Joseph, N.; Fazal, F. Phytochemistry and medicinal uses of the bael fruit (Aegle marmelos Correa): A concise review. Food Res. Int. 2011, 44, 1768–1775. [Google Scholar] [CrossRef]
- Mujeeb, F.; Bajpai, P.; Pathak, N. Phytochemical evaluation, antimicrobial activity, and determination of bioactive components from leaves of Aegle marmelos. Biomed. Res. Int. 2014, 11, 497606. [Google Scholar]
- Sarkar, T.; Salauddin, M.; Chakraborty, R. In-depth pharmacological and nutritional properties of bael (Aegle marmelos): A critical review. J. Agric. Food Res. 2020, 2, 100081. [Google Scholar] [CrossRef]
- Mahomoodally, M.F.; Mollica, A.; Stefanucci, A.; Aumeeruddy, M.Z.; Poorneeka, R.; Zengin, G. Volatile components, pharmacological profile, and computational studies of essential oil from Aegle marmelos (Bael) leaves: A functional approach. Ind. Crops Prod. 2018, 126, 13–21. [Google Scholar] [CrossRef]
- Chhabra, R.; Sharma, R.; Kaur, T. Phyto-allelopathic effect of different trees leaves’ aqueous extracts on seed germination and seedling growth of Echinochloa crus-galli (L.) Beauv. Indian J. Weed Sci. 2021, 53, 318–323. [Google Scholar] [CrossRef]
- Islam, A.K.M.M.; Hasan, M.; Musha, M.M.H.; Uddin, M.K.; Juraimi, A.S.; Anwar, M.P. Exploring 55 tropical medicinal plant species available in Bangladesh for their possible allelopathic potentiality. Ann. Agric. Sci. 2018, 63, 99–107. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Matsumoto, K.; Sakamoto, C.; Tojo, S.; Teruya, T. Allelopathy and allelopathic substances in the leaves of Metasequoia glyptostroboides from pruned branches for weed management. Agronomy 2023, 13, 1017. [Google Scholar] [CrossRef]
- Ito, C.; Furukawa, H. Constituents of Murraya exotica L. Structure elucidation of new coumarins. Chem. Pharm. Bull. 1987, 35, 4277–4285. [Google Scholar] [CrossRef]
- Prachayasittikul, S.; Suphapong, S.; Worachartcheewan, A.; Lawung, R.; Ruchirawat, S.; Prachayasittikul, V. Bioactive metabolites from Spilanthes acmella Murr. Molecules 2009, 14, 850–867. [Google Scholar] [CrossRef]
- Lee, Y.G.; Cho, J.Y.; Kim, C.M.; Lee, S.H.; Kim, W.S.; Jeon, T.I.; Park, K.H.; Moon, J.H. Coumaroyl quinic acid derivatives and flavonoids from immature pear (Pyrus pyrifolia nakai) fruit. Food Sci. Biotechnol. 2013, 22, 803–810. [Google Scholar] [CrossRef]
- Tang, Y.Q.; Chu, C.Y.; Zhu, L.; Qian, B.; Shao, L.X. N-Heterocyclic carbene-Pd (II) complex derived from proline for the mizoroki-heck reaction in water. Tetrahedron 2011, 67, 9479–9483. [Google Scholar] [CrossRef]
- Mamidi, N.; Manna, D. Zn (OTf) 2-promoted chemoselective esterification of hydroxyl group bearing carboxylic acids. J. Org. Chem. 2013, 78, 2386–2396. [Google Scholar] [CrossRef]
- Bari, I.N.; Kato-Noguchi, H. Phytotoxic effects of Cerbera manghas L. leaf extracts on seedling elongation of four monocot and four dicot test species. Acta Agrobot. 2017, 70, 1–7. [Google Scholar] [CrossRef]
- Moh, S.M.; Tojo, S.; Teruya, T.; Kato-Noguchi, H. Allelopathic activity of a novel compound, two known sesquiterpenes, and a C13 nor-isopenoid from the leave of Croton oblongifolius Roxb. for weed control. Plants 2023, 12, 3384. [Google Scholar] [CrossRef]
- Rob, M.M.; Hossen, K.; Ozaki, K.; Teruya, T.; Kato-Noguchi, H. Phytotoxicity and phytotoxic substances in Calamus tenuis Roxb. Toxins 2023, 15, 595. [Google Scholar] [CrossRef]
- Sim, M.O.; Ham, J.R.; Lee, H.I.; Seo, K.I.; Lee, M.K. Long-term supplementation of umbelliferone and 4-methylumbelliferone alleviates high-fat diet induced hypertriglyceridemia and hyperglycemia in mice. Chem. Biol. Interact. 2014, 216, 9–16. [Google Scholar] [CrossRef]
- Radha, G.V.; Sadhana, B.; Sastri, K.T.; Ganapaty, S. Bioactive umbelliferone and its derivatives: An update. J. Pharmacogn. Phytochem. 2019, 8, 59–66. [Google Scholar]
- Hoult, J.R.S.; Payá, M. Pharmacological and biochemical actions of simple coumarins: Natural products with therapeutic potential. Gen. Pharmacol. Vasc. Syst. 1996, 27, 713–722. [Google Scholar] [CrossRef]
- Navarro-García, V.M.; Rojas, G.; Avilés, M.; Fuentes, M.; Zepeda, G. In vitro antifungal activity of coumarin extracted from Loeselia mexicana Brand. Mycoses 2011, 54, 569–571. [Google Scholar] [CrossRef]
- Facenda, G.; Real, M.; Galán-Pérez, J.A.; Gámiz, B.; Celis, R. Soil effects on the bioactivity of hydroxycoumarins as plant allelochemicals. Plants 2023, 12, 1278. [Google Scholar] [CrossRef]
- Rezaeiroshan, A.; Saeedi, M.; Morteza-Semnani, K.; Akbari, J.; Hedayatizadeh-Omran, A.; Goli, H.; Nokhodchi, A. Vesicular formation of trans-ferulic acid: An efficient approach to improve the radical scavenging and antimicrobial properties. J. Pharm. Investig. 2022, 17, 652–661. [Google Scholar] [CrossRef]
- Trombino, S.; Cassano, R.; Ferrarelli, T.; Barone, E.; Picci, N.; Mancuso, C. Trans-ferulic acid-based solid lipid nanoparticles and their antioxidant effect in rat brain microsomes. Colloids Surf. B 2013, 109, 273–279. [Google Scholar] [CrossRef]
- Singh, H.P.; Kaur, S.; Batish, D.R.; Kohli, R.K. Ferulic acid impairs rhizogenesis and root growth, and alters associated biochemical changes in mung bean (Vigna radiata) hypocotyls. J. Plant Interact. 2014, 9, 267–274. [Google Scholar] [CrossRef]
- Fu, Y.H.; Quan, W.X.; Li, C.C.; Qian, C.Y.; Tang, F.H.; Chen, X.J. Allelopathic effects of phenolic acids on seedling growth and photosynthesis in Rhododendron delavayi Franch. Photosynthetica 2019, 57, 377–387. [Google Scholar] [CrossRef]
- Chandra, S.; Roy, A.; Jana, M.; Pahan, K. Cinnamic acid activates PPARα to stimulate lysosomal biogenesis and lower amyloid plaque pathology in an Alzheimer’s disease mouse model. Neurobiol. Dis. 2019, 124, 379–395. [Google Scholar] [CrossRef]
- Chafer, A.; Tiziana, F.; Stateva, R.P.; Angel, B. Trans-cinnamic acid solubility enhancement in the presence of ethanol as a supercritical CO2 co-solvent. J. Chem. Eng. 2009, 54, 2263–2268. [Google Scholar]
- El-Raouf, O.M.A.; El-Sayed, M.; Manie, M.F. Cinnamic acid and cinnamaldehyde ameliorate cisplatin-induced splenotoxicity in rats. J. Biochem. Mol. Toxicol. 2015, 29, 426–431. [Google Scholar] [CrossRef]
- Baziramakenga, R.; Simard, R.R.; Leroux, G.D. Effects of benzoic and cinnamic acids on growth, mineral composition, and chlorophyll content of soybean. J. Chem. Ecol. 1994, 20, 2821–2833. [Google Scholar] [CrossRef]
- Yu, J.Q.; Matsui, Y. Phytotoxic substances in the root exudates of cucumber (Cucumis sativus L.). J. Chem. Ecol. 1994, 20, 21–31. [Google Scholar] [CrossRef]
- Chon, S.U.; Choi, S.K.; Jung, S.; Jang, H.G.; Pyo, B.S.; Kim, S.M. Effects of alfalfa leaf extracts and phenolic allelochemicals on early seedling growth and root morphology of alfalfa and barnyard grass. Crop. Prot. 2002, 21, 1077–1082. [Google Scholar] [CrossRef]
- Anh, L.H.; Quan, N.V.; Nghia, L.T.; Xuan, T.D. Phenolic allelochemicals: Achievements, limitations, and prospective approaches in weed management. Weed Biol. Manag. 2021, 21, 37–67. [Google Scholar]
- Sova, M. Antioxidant and antimicrobial activities of cinnamic acid derivatives. Mini Rev. Med. Chem. 2012, 12, 749–767. [Google Scholar] [CrossRef] [PubMed]
- Medvedeva, M.; Barinova, K.; Melnikova, A.; Semenyuk, P.; Kolmogorov, V.; Gorelkin, P.; Erofeev, A.; Muronetz, V. Naturally occurring cinnamic acid derivatives prevent amyloid transformation of alpha-synuclein. Biochimie 2020, 170, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Da Cunha, F.M.; Duma, D.; Assreuy, J.; Buzzi, F.C.; Niero, R.; Campos, M.M.; Calixto, J.B. Caffeic acid derivatives: In vitro and in vivo anti-inflammatory properties. Free Radic. Res. 2004, 38, 1241–1253. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N. Chlorogenic acids and other cinnamates—Nature, occurrence and dietary burden. J. Sci. Food Agric. 1999, 79, 362–372. [Google Scholar] [CrossRef]
- Jakobek, L.; Seruga, M.; Novak, I.; Medvidovic-Kosanovic, M. Flavonols, phenolic acids and antioxidant activity of some red fruits. Deut. Lebensm-Rundsch. 2007, 103, 369–378. [Google Scholar]
- Papetti, A.; Daglia, M.; Aceti, C.; Sordelli, B.; Spini, V.; Carazzone, C.; Gazzani, G. Hydroxycinnamic acid derivatives occurring in Cichorium endivia vegetables. J. Pharmaceut. Biomed. 2008, 48, 472–476. [Google Scholar] [CrossRef] [PubMed]
- DellaGreca, M.; Previtera, L.; Purcaro, R.; Zarrelli, A. Cinnamic ester derivatives from Oxalis pes-caprae (Bermuda buttercup). J. Nat. Prod. 2007, 70, 1664–1667. [Google Scholar] [CrossRef]
- Glass, A.D.M.; Dunlop, J. Influence of phenolic acids on ion uptake. Plant Physiol. 1974, 54, 855–858. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Q.; Ruan, X.; Pan, C.; Jiang, D. Phenolics and plant allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef]
- Patterson, D.T. Effects of allelopathic chemicals on growth and physiological response of soybean (Glycine max). Weed Sci. 1981, 29, 53–58. [Google Scholar] [CrossRef]
- Możdżeń, K.; Barabasz-Krasny, B.; Stachurska-Swakoń, A.; Turisová, I.; Zandi, P. Germination and growth of radish under influence of nipplewort aqueous extracts. Not. Bot. Horti Agrobot. Cluj.-Napoca. 2021, 49, 12195. [Google Scholar] [CrossRef]
- Doblinski, P.M.F.; Ferrarese, M.L.L.; Huber, D.A.; Scapim, C.A.; Braccini, A.L.; Ferrarese-Filho, O. Peroxidase and lipid peroxidation of soybean roots in response to p-coumaric and p-hydroxybenzoic acids. Braz. Arch. Biol. Technol. 2003, 46, 193–198. [Google Scholar] [CrossRef]
- Ju, D.; Sun, Y.; Xiao, C.L.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Physiological basis of different allelopathic reactions of cucumber and figleaf gourd plants to cinnamic acid. J. Exp. Bot. 2007, 58, 3765–3773. [Google Scholar]
- Pinho, I.A.; Lopes, D.V.; Martins, R.C.; Quina, M.J. Phytotoxicity assessment of olive mill solid wastes and the influence of phenolic compounds. Chemosphere 2017, 185, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Jose, C.M.; Torres, L.M.B.; Torres, M.A.M.G.; Shirasuna, R.T.; Farias, D.A.; dos Santos, N.A., Jr.; Grombone-Guaratini, M.T. Phytotoxic effects of phenolic acids from Merostachys riedeliana, a native and over abundant Brazilian bamboo. Chemoecology 2016, 26, 235–246. [Google Scholar] [CrossRef]
- Ladhari, A.; Romanucci, V.; De Marco, A.; Di Fabio, G.; Zarrelli, A. Phytotoxic effects of Mediterranean plants extract on lettuce, tomato and onion as possible additive in irrigation drips. Allelopath. J. 2018, 44, 233–244. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Plant Species | I50 (mg DW Equivalent A. marmelos Extract/mL) | Correlation Coefficient ® | |||
|---|---|---|---|---|---|
| Hypocotyl/Coleoptile | Root | Hypocotyl/Coleoptile | Root | ||
| Dicot | L. sativum | 3.3 | 2.6 | −0.79 ** | −0.79 ** |
| L. sativa | 10.0 | 8.3 | −0.83 ** | −0.91 ** | |
| M. sativa | 3.5 | 2.3 | −0.70 ** | −0.75 ** | |
| Monocot | E. crusgalli | 36.1 | 8.9 | −0.78 ** | −0.71 ** |
| L. multiflorum | 12.0 | 10.6 | −0.70 ** | −0.77 ** | |
| P. pratense | 3.1 | 1.61 | −0.76 ** | −0.73 ** | |
| Identified Compound | I50 (µM) | |||
|---|---|---|---|---|
| L. sativum | E. crusgalli | |||
| Hypocotyl | Root | Coleoptile | Root | |
| Compound-1 | 378.2 | 153.1 | ||
| Compound-2 | 785.4 | 552.6 | ||
| Compound-3 | 179.3 | 83.8 | ||
| Compound-4 | 119.6 | 74.1 | ||
| Compound-5 | 260.5 | 181.8 | 885.1 | 453.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moh, S.M.; Tojo, S.; Teruya, T.; Kato-Noguchi, H. Allelopathy and Identification of Five Allelochemicals in the Leaves of the Aromatic Medicinal Tree Aegle marmelos (L.) Correa. Plants 2024, 13, 559. https://doi.org/10.3390/plants13040559
Moh SM, Tojo S, Teruya T, Kato-Noguchi H. Allelopathy and Identification of Five Allelochemicals in the Leaves of the Aromatic Medicinal Tree Aegle marmelos (L.) Correa. Plants. 2024; 13(4):559. https://doi.org/10.3390/plants13040559
Chicago/Turabian StyleMoh, Seinn Moh, Shunya Tojo, Toshiaki Teruya, and Hisashi Kato-Noguchi. 2024. "Allelopathy and Identification of Five Allelochemicals in the Leaves of the Aromatic Medicinal Tree Aegle marmelos (L.) Correa" Plants 13, no. 4: 559. https://doi.org/10.3390/plants13040559
APA StyleMoh, S. M., Tojo, S., Teruya, T., & Kato-Noguchi, H. (2024). Allelopathy and Identification of Five Allelochemicals in the Leaves of the Aromatic Medicinal Tree Aegle marmelos (L.) Correa. Plants, 13(4), 559. https://doi.org/10.3390/plants13040559